Г. Мендель установил важнейшие закономерности наследственности организмов и вскрыл дискретную (прерывную) природу ее. Доказав возможность наследования одного признака неза-висимо от других, он тем самым показал, что наследственность дискретна, делима, и генотип со-стоит из отдельных единиц, определяющих отдельные признаки и относительно независимых друг от друга. Принцип дискретности наследственности лежит в основе всех современных методов селекции: сложной ступенчатой гибридизации, индивидуального отбора, насыщающих скрещиваний, получения гетерозисных гибридов, методов создания стерильных аналогов и восстановителей фертильности и т. д.
Тема. Наследование признаков при взаимодействии генов.
Лекция 4
План.
1. Плейотропное действие генов.
2. Комплементарное взаимодействие генов.
3. Эпистаз.
4. Криптомерия.
5. Полимерия.
6. Модифицирующее действие генов.
Список литературных источников.
1. Гуляев Г.В. Генетика. – М.: Колос, 1984. – С 7487.
2. Дубинин Н.П. Горизонты генетики. Пособие для учителей. – М.: Просвещение, 1970. – С.
108116.
3. Жученко А.А., Гужов Юрий Леонидович. Генетика. – М.: КолосС, 2003. – С. 5365.
4. ИнгеВечтомов С.Г. Генетика с основами селекции. – М.: Выс. шк., 1989. – С. 4254.
1. Плейотропное действие генов.
Г. Мендель установил важнейшие закономерности наследственности организмов и вскрыл
дискретную (прерывную) природу ее. Доказав возможность наследования одного признака
независимо от других, он тем самым показал, что наследственность дискретна, делима, и генотип
состоит из отдельных единиц, определяющих отдельные признаки и относительно независимых друг
от друга. Принцип дискретности наследственности лежит в основе всех современных методов
селекции: сложной ступенчатой гибридизации,
индивидуального отбора, насыщающих
скрещиваний, получения гетерозисных гибридов, методов создания стерильных аналогов и
восстановителей фертильности и т. д.
Наследование при взаимодействии генов. Правильность установленных Г. Менделем
закономерностей наследственности была подтверждена после 1900 г. в многочисленных опытах по
изучению наследования различных признаков, как у растений, так и у животных. В то же время
выяснилось, что полученные Г. Менделем определенные числовые отношения при расщеплении в
потомстве гибридов были верными во всех тех случаях, когда каждый ген определял развитие
одного наследственного признака.
Но было накоплено много фактов, указывающих на то, что взаимоотношения между генами
и признаками, которые они определяют, носят более сложный и многообразный характер.
Выяснилось, что, вопервых, один и тот же ген может оказывать влияние на несколько различных
признаков и, вовторых, происходит взаимодействие генов, когда один и тот же наследственный
признак развивается под влиянием многих из них. Таким образом, фенотипическое выражение
большинства признаков и свойств организма определяется в онтогенезе взаимодействием многих
генов. Это отражается и на характере расщепления гибридов различных скрещиваний, особенно
если родительские формы различаются по нескольким признакам.
Открытие явления взаимодействия генов имело важнейшее значение для всего
последующего развития генетики. На основе этих фактов было отброшено представление об
организме как мозаике наследственных факторов, высказанное в конце XIX в. немецким биологом
А. Вейсманом. Оказалось, что наследственный фактор нельзя рассматривать как зачаток будущего
признака, и в организме абсолютной независимости генов друг от друга, как она представлялась Г.
Менделю, не существует. На смену этим взглядам было выдвинуто положение о сложной связи и
взаимодействии генов в системе генотипа при развитии любого признака организма.
Плейотропия. Влияние одного гена на развитие двух и большего числа признаков
называется множественным, или плейотропным, действием, а само это явление получило
название плейотропии (от греч. pleistos — множественный, наибольший). Биохимическая природа
плейотропного действия генов выяснена довольно хорошо. Один белокфермент, образующийся
1под контролем одного гена, определяет развитие не только данного признака, но воздействует и на
вторичные реакции биосинтеза различных других признаков и свойств, вызывая их изменение.
Плейотропия широко распространена: большинство генов у всех организмов действует
плейотропно. Это явление впервые было обнаружено Г. Менделем. Он установил, что у растений с
пурпурными цветками одновременно всегда имелись красные пятна в пазухах листьев, а семенная
кожура была серого или бурого цвета. Эти три признака определялись действием одного
наследственного фактора. Недавно было установлено, что многим индуцированным мутациям
гороха свойственна высокая степень плейотропии, проявляющаяся в изменении до десяти и более
признаков. Н. И. Вавилов и О. В. Якушкина, изучая наследование некоторых признаков у
персидской пшеницы (Tritikum persikum), выяснили, что доминантный ген черной окраски колоса
одновременно вызывает опушение колосковых чешуй.
Известны два вида взаимодействия генов: аллельное и неаллельное. Простейший пример
аллельного взаимодействия генов неполное доминирование при скрещивании красноцветковых и
белоцветковых растений львиного зева или ночной красавицы. Розовая окраска цветка у гибридов
F1 в этом скрещивании – результат взаимодействия двух аллельных генов А и а. Полное
доминирование также является результатом взаимодействия двух генов одной аллельной пары. При
этом доминантный ген подавляет проявление рецессивного гена.
Взаимодействие генов имеет биохимическую природу. Оно основано на взаимодействии
синтезируемых под контролем генов белковферментов.
Взаимодействие неаллельных генов
проявляется в четырех основных формах:
комплементарное, эпистаза, полимерии и модифицирующего действия генов. Каждая из этих форм
приводит к характерным изменениям известных числовых отношений при расщеплении в
дигибридных скрещиваниях.
Неаллельное взаимодействие генов – это развитие признака при совместном действии двух
или нескольких неаллельных генов. Это явление было открыто В. Бэтсоном в начале ХХ века и
затем получило широкое развитие. Различают несколько типов неаллельного взаимодействия генов:
комплементарное, эпистатическое, полимерное и модифицирующее действие.
Схемы записи результатов гибридологического анализа при всех этих типах взаимодействия
неаллельных генов ничем не отличаются от записи их при дигибридном или полигибридном
скрещивании. Сходны и результаты скрещивания по типам гамет. Единственное различие – в типах
и количественном соотношении фенотипов. Для каждого из этих скрещиваний они специфичны, и
именно по этому показателю определяется сам тип взаимодействия.
2. Комплементарное взаимодействие генов
К комплементарным, или дополнительным генам относят такие гены, которые при
совместном действии в генотипе в гомо или гетерозиготном состояниях (АB) обуславливают
развитие нового фенотипа признака, не свойственного ни одному из родителей.
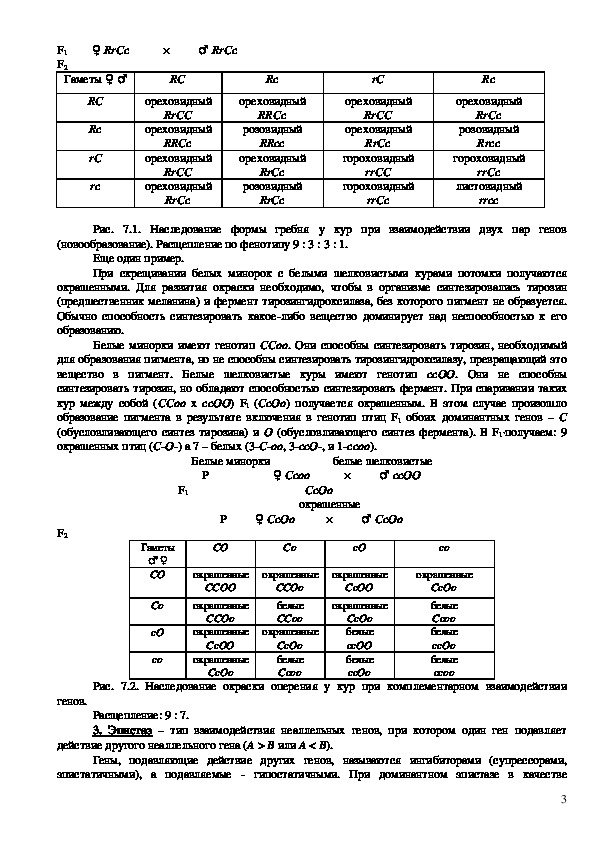
Например, у кур гены розовидного и гороховидного гребня не являются аллельными.
Гороховидный и розовидный гребни доминируют над листовидными. При скрещивании кур породы
виандот, имеющих розовидный гребень (RRcc) с петухами породы брама с гороховидным гребнем
(rrCC) у потомков F1 (RrCc) в результате взаимодействия двух доминантных генов R и C
развивается новая форма гребня – ореховидная. Скрещивание потомков F1 между собой ведет к
получению в F2 четырех разных фенотипов в соотношении: 9 с гребнем ореховидной формы (R–
C–), 3 с гребнем розовидной формы (R–сс), 3 со гороховидным гребнем (rrC) и 1 – с
листовидным (rrcc).
Взаимодействие неаллельных генов R и C обусловливает образование новой формы гребня, в
то время как каждый из этих генов в отдельности проявляет свой собственный эффект. Особь с
листовидным гребнем является двойным рецессивом (rrcc).
розовидный
♀ RRcc
×
гороховидный
♂ rrCC
Р
ореховидный
RrCc
2♀ RrCc
F1
F2
Гаметы ♀ ♂
RC
Rc
rC
rc
×
♂ RrCc
RC
Rc
rC
ореховидный
RrCC
ореховидный
RRCc
ореховидный
RrCC
ореховидный
RrCc
ореховидный
RRCc
розовидный
RRcc
ореховидный
RrCc
розовидный
RrCc
ореховидный
RrCC
ореховидный
RrCc
гороховидный
rrCC
гороховидный
rrCc
Rc
ореховидный
RrCc
розовидный
Rrcc
гороховидный
rrCc
листовидный
rrcc
Рис. 7.1. Наследование формы гребня у кур при взаимодействии двух пар генов
(новообразование). Расщепление по фенотипу 9 : 3 : 3 : 1.
Еще один пример.
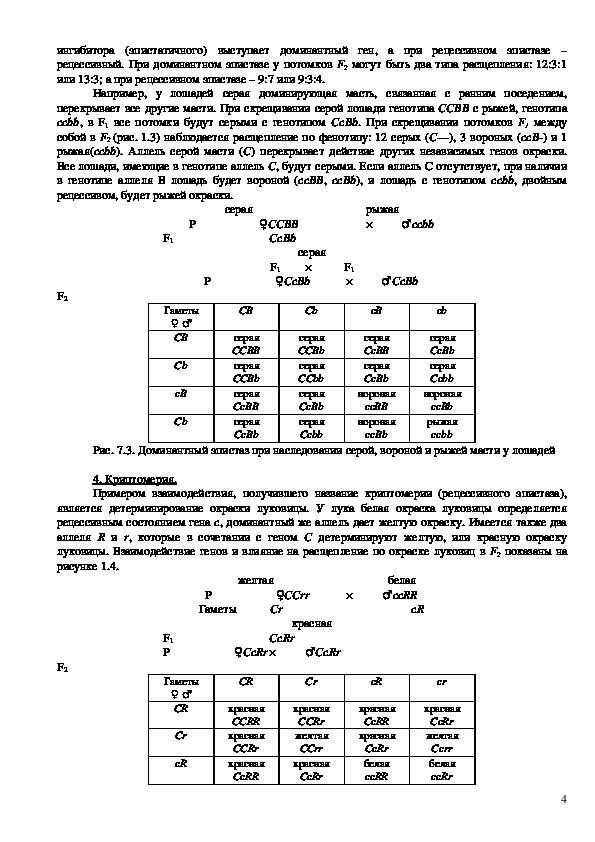
При скрещивании белых минорок с белыми шелковистыми курами потомки получаются
окрашенными. Для развития окраски необходимо, чтобы в организме синтезировались тирозин
(предшественник меланина) и фермент тирозингидроксилаза, без которого пигмент не образуется.
Обычно способность синтезировать какоелибо вещество доминирует над неспособностью к его
образованию.
Белые минорки имеют генотип ССоо. Они способны синтезировать тирозин, необходимый
для образования пигмента, но не способны синтезировать тирозингидроксилазу, превращающий это
вещество в пигмент. Белые шелковистые куры имеют генотип ссОО. Они не способны
синтезировать тирозин, но обладают способностью синтезировать фермент. При спаривании таких
кур между собой (ССоо х ссОО) F1 (СсОо) получается окрашенным. В этом случае произошло
образование пигмента в результате включения в генотип птиц F1 обоих доминантных генов – С
(обусловливающего синтез тирозина) и О (обусловливающего синтез фермента). В F1`получаем: 9
окрашенных птиц (СО) а 7 – белых (3Соо, 3ссО, и 1ссоо).
Белые минорки
♀ Ссоо
Р
F1
белые шелковистые
×
♂ ссОО
СсОо
окрашенные
×
♂ СсОо
♀ СсОо
Со
сО
со
Р
СО
Рис. 7.2. Наследование окраски оперения у кур при комплементарном взаимодействии
Расщепление: 9 : 7.
3. Эпистаз – тип взаимодействия неаллельных генов, при котором один ген подавляет
действие другого неаллельного гена (А > В или А < В).
Гены, подавляющие действие других генов, называются ингибиторами (супрессорами,
эпистатичными), а подавляемые гипостатичными. При доминантном эпистазе в качестве
3
F2
генов.
Гаметы
♂ ♀
СО
Со
сО
со
окрашенные
окрашенные
окрашенные
окрашенные
ССОО
окрашенные
ССОо
ССОо
белые
ССоо
окрашенные
окрашенные
СсОО
окрашенные
СсОо
СсОо
белые
Ссоо
СсОО
окрашенные
СсОо
белые
ссОО
белые
ссОо
СсОо
белые
Ссоо
белые
ссОо
белые
ссооингибитора (эпистатичного) выступает доминантный ген, а при рецессивном эпистазе –
рецессивный. При доминантном эпистазе у потомков F2 могут быть два типа расщепления: 12:3:1
или 13:3; а при рецессивном эпистазе – 9:7 или 9:3:4.
Например, у лошадей серая доминирующая масть, связанная с ранним поседением,
перекрывает все другие масти. При скрещивании серой лошади генотипа ССВВ с рыжей, генотипа
ссbb, в F1 все потомки будут серыми с генотипом СсВb. При скрещивании потомков F1 между
собой в F2 (рис. 1.3) наблюдается расщепление по фенотипу: 12 серых (С), 3 вороных (ссВ) и 1
рыжая(ссbb). Аллель серой масти (С) перекрывает действие других независимых генов окраски.
Все лошади, имеющие в генотипе аллель С, будут серыми. Если аллель С отсутствует, при наличии
в генотипе аллеля В лошадь будет вороной (ссВВ, ссВb), и лошадь с генотипом ссbb, двойным
рецессивом, будет рыжей окраски.
F2
Р
F1
P
Гаметы
♀ ♂
CB
Сb
cB
Cb
серая
♀ССВВ
СсВb
рыжая
×
♂ссbb
серая
F1
×
♀CcBb
F1
×
СВ
серая
ССВВ
серая
ССВb
серая
CcBB
серая
CcBb
Сb
серая
ССВb
серая
ССbb
серая
CcBb
серая
Ccbb
♂CcBb
cB
cb
серая
СсВВ
серая
СсВb
вороная
ccBB
вороная
ccBb
серая
СсВb
серая
Ссbb
вороная
ccBb
рыжая
ccbb
Рис. 7.3. Доминантный эпистаз при наследовании серой, вороной и рыжей масти у лошадей
4. Криптомерия.
Примером взаимодействия, получившего название криптомерии (рецессивного эпистаза),
является детерминирование окраски луковицы. У лука белая окраска луковицы определяется
рецессивным состоянием гена c, доминантный же аллель дает желтую окраску. Имеется также два
аллеля R и r, которые в сочетании с геном С детерминируют желтую, или красную окраску
луковицы. Взаимодействие генов и влияние на расщепление по окраске луковиц в F2 показаны на
рисунке 1.4.
желтая белая
♂ссRR
Р
Гаметы
cR
♀ССrr
Cr
×
красная
F2
F1
P
Гаметы
♀ ♂
CR
Сr
cR
СсRr
♀CcRr×
♂CcRr
СR
Сr
cR
cr
красная
СCRR
красная
ССRr
красная
CcRR
красная
ССRr
желтая
ССrr
красная
CcRr
красная
СсRR
красная
СсRr
белая
ccRR
красная
СсRr
желтая
Ссrr
белая
ccRr
4cr
красная
CcRr
желтая
Ccrr
белая
ccRr
белая
ccrr
Рис. 7.4. Наследование окраски луковицы при криптомерии.
В итоге:
CR красная (9); Crr – желтая (3); ccR белая (3); ccrr – белая (1).
Расщепление в F2 составило 9:3:4.
5. Полимерией называется такое явление, когда развитие того или иного признака
организма обусловлено взаимодействием двух или более пар генов, оказывающих сходное
воздействие на развитие этого признака.
Полимерия может проходить по аддитивному (когда действие полимерных генов
суммируется) или по мультативному типу (действие полимерных генов как бы перемножается).
По типу полимерии наследуются все количественные признаки (живая масса, прирост, удой,
жирномолочность, настриг шерсти, яйценоскость и др.). При полимерном наследовании развитие
признака обусловливается двумя или несколькими парами однозначно действующих генов. Чем
больше доминантных полимерных генов содержит организм, тем сильнее выражен признак.
Полимерные гены обозначаются одинаковыми буквами с соответствующим индексом.
Например, три пары полимерных генов можно обозначить А1 А1 А2 А2 А3 А3 или а1 а1 а2 а2 а3 а3.
Рассмотрим пример наследования окраски зерен пшеницы при взаимодействии 2х пар
полимерных генов. Различают две основные окраски зерен: красную и белую. Исходя из этого,
генотип пшеницы с темнокрасным зерном, будет А1 А1 А2 А2, а с белым – а1 а1 а 2а2. Темнокрасная
окраска – 4А, красная – 3А, светло красная – 2А, бледнокрасная – 1А, белая – 4а.
При скрещивании пшеницы с темнокрасной окраской зерна с белозерной пшеницей
получим:
темнокрасное зерно
♀А1 А1 А2 А2 ×
белое зерно
♂а1 а1 а2 а2
светлокрасное зерно
А1 а1 А2 а2
Продолжая скрещивание, получим F2
x
F1
♀A1 a1 A2 a2
F1
×
♂A1 a1 A2 a2
Р
F1
P
В F2 получили расщепление по фенотипу (рис. 1.5): 1 часть растений с темнокрасным
зерном (А1А1А2А2); 4 – с красным (2 а1А1А2 А2; 2 А1А1а2А2); 6 со светлокрасным (4 А1а1А2а2;
А1А1а2а2; а1а1А2А2); 4 – с бледнокрасным (2 А1а1а2а2; 2а1а1А2а2); 1 – с белым зерном (а1а1а2а2) –
1:4:6:4:1.
F2
Гаметы
♂♀
А1А2
А1а2
а1А2
а1а2
А1А2
темнокрасное
А1А1А2А2
красное
А1А1А2а2
красное
А1а1А2А2
А1а2
красное
А1А1А2а2
а1А2
красное
А1а1А2А2
а1а2
светлокрасное
А1а1А2а2
светлокрасное
светлокрасное
бледнокрасное
А1А1а2а2
А1а1А2а2
А1а1а2а2
светлокрасное
светлокрасное
бледнокрасное
А1а1А2а2
а1а1А2А2
а1а1А2а2
белое зерно
светлокрасное
бледнокрасное
бледнокрасное
А1а1А2а2
а1а1А2а2
Рис. 7.5. Наследование окраски зерен пшеницы при полимерии.
6.Модифицирующее действие генов
Гены, не проявляющие собственного действия, но усиливающие или ослабляющие эффект
действия других генов, называются генамимодификаторами.
А1а1а2а2
а1а1а2а2
5Взаимодействие
неаллельных генов
комплементарное
эпистаз доминантный
эпистаз рецессивный
9 : 3 : 3 : 1
9 : 7
9 : 3 : 4
9 : 6 : 1
13 : 3
12 : 3 : 1
9 : 7
9 : 3 : 4
аллели независимы
аллели суммируются
аллели независимы
аллели обладают сходным
фенотипическим эффектом
доминантные аллели одного гена
подавляют проявление доминантного
аллеля другого признака
гомозиготные рецессивные аллели одного
гена подавляют проявление доминантной
или рецессивной аллели другого типа
действие полимерных генов суммируется
Например, у ряда пород кур (черные испанские, орпингтоны, польские и др.) при действии
геновослабителей черной пигментации происходит ослабление и нарушение стандартной окраски
оперения.
Ген Li – ослабитель коричневой окраски оперения, сцепленный с полом, превращает все
участки оперения коричневой окраски в бледножелтые. Рецессивный аутосомный мутантный ген
лавандовой окраски lav превращает черную окраску оперения в серую, а красную в палевую.
У кроликов известен ген модификатор Н, усиливающий голубую масть у венской голубой
породы.
У крупного рогатого скота генымодификаторы контролируют пеструю окраску у группы
чернопестрых пород.
Модели расщепления при взаимодействии неаллельных генов.
Таблица 5
Матеем. модель
расщеп ления
Тип взаимодействия аллелей разных генов
Полимерия
1 : 4 : 6 : 4 : 1
Летальные гены
Известны случаи, когда один ген может оказывать влияние на несколько признаков, в том
числе на жизнеспособность. У человека и других млекопитающих определенный рецессивный ген
вызывает образование внутренних спаек легких, что приводит к смерти при рождении. Другим
примером служит ген, который влияет на формирование хряща и вызывает врожденные уродства,
ведущие к смерти плода или новорожденного.
У кур, гомозиготных по аллелю, вызывающему “курчавость” перьев, неполное развитие
перьев влечет за собой несколько фенотипических эффектов. У таких кур теплоизоляция
недостаточна, и они страдают от охлаждения. Для компенсации потери тепла у них появляется ряд
структурных и физиологических адаптаций, но эти адаптации малоэффектны и среди таких кур
высока смертность.
Воздействие летального гена ясно видно на примере наследования окраски шерсти у мышей.
У диких мышей шерсть обычно серая, типа агути; но у некоторых мышей шерсть желтая. При
скрещиваниях между желтыми мышами в потомстве получаются как желтые мыши, так и агути в
отношении 2:1. Единственное возможное объяснение таких результатов состоит в том, что желтая
окраска шерсти доминирует над агути и что все желтые мыши гетерозиготны. Атипичное
менделевское отношение объясняется гибелью гомозиготных желтых мышей до рождения. При
вскрытии беременных желтых мышей, скрещенных с желтыми же мышами, в их матках были
обнаружены мертвые желтые мышата. Если же скрещивались желтые мыши и агути, то в матках
беременных самок не оказывалось мертвых желтых мышат, поскольку при таком скрещивании не
может быть потомства, гомозиготного по гену желтой шерсти.
6