Половое размножение характерно для большинства живых организмов. Перекомбинация генов или хромосом в процессе мейоза и оплодотворения является основой наследственной изменчивости. Половой процесс связан с разделением по полу и наличием женских и мужских особей или женских и мужских органов у одного организма, при этом одна особь может производить как женские, так и мужские гаметы.
Лекция № 5.
Тема. Генетика пола. Сцепленное с полом наследование.
Типы определения пола (у животных, у человека и растений).
Наследование сцепленное с полом.
Нерасхождение половых хромосом.
План.
1.
2.
3.
Половое размножение характерно для большинства живых организмов. Перекомбинация
генов или хромосом в процессе мейоза и оплодотворения является основой наследственной
изменчивости. Половой процесс связан с разделением по полу и наличием женских и мужских
особей или женских и мужских органов у одного организма, при этом одна особь может
производить как женские, так и мужские гаметы.
Определение пола может происходить на разных этапах цикла размножения. Выделяют
следующие типы определения пола.
Сингамный – преобладание женской или мужской тенденции развития проявляется в момент
слияния гамет и образования зиготы. Сингамия характерна для большинства растений, птиц, рыб,
млекопитающих.
Прогамный – т. е. происходящий до оплодотворения тип определения пола, встречается у
немногих организмов (коловраток – Rotatoria, первичных кольчецов – Dinophilus, тлей – Phylloxera
vastatrix). У них пол зиготы определяется еще в процессе онтогенеза, при этом у самок в следствие
неравномерного деления цитоплазмы образуются крупные и мелкие яйца, после оплодотворения из
крупных яиц развиваются самки, из мелких – самцы.
Эпигамный – происходящий после оплодотворения, наиболее редкий тип определения пола.
В качестве примера может служить морской червь, у которого очень мелкие самцы обитают в
матке более крупных самок. Если свободноплавающие личинки приклепляются к камням, то они
развиваются в самках, а если попадают на хоботок самки, то превращаются в самцов и мигрируют в
ее половые органы.
Хромосомный механизм определения пола при сингамии.
К настоящему времени установлено, что пол у большинства раздельнополых организмов
детерминируется в основном хромосомным механизмом, однако факторы внешней среды могут
регулировать и видоизменять половой фенотип.
Г. Мендель в своих работах предполагал, что пол наследуется как любой признак при
анализирующем скрещивании моногетерозигот (Аа) с рецессивной гомозиготой (аа). Поскольку
гетерозигота Аа дает гаметы двух типов (А и а) в равном соотношении, а гомозигота (аа) – гаметы
только одного типа – а, то и потомство будет образовываться в таком же равном соотношении (1 :
1), как и исходное развитие (F1 аа +Аа) таким образом, был сделан вывод, что один пол должен
быть гомозиготным ( в последствие его стаои называть гомогаметным), а другой – гетерозигтным
(гетерогаметным).
Первые доказательства этого предположения были получены в опытах Донкастера (1906) с
бабочкой пяденицей крыжоновноковой и Корренса (1907) в скрещивании двудомных и однодомных
растений и рода переступень из семейства тыквенных.
Диплоидный набор хромосом возникает при оплодотворении, когда гаплоидные гаметы
материнского и отцовского организмов образуют зиготу. Гаплоидные наборы хромосом у гамет
точно соответствуют друг другу и в зиготе образуют пары гомологичных хромосом. Однако при
цитологическом изменении клеток женского и мужского организмов клопов рода Protenor было
обнаружено неравное распределение хромосом. У самцов при мейозе в клетки сперматоцитов
второго порядка одни клетки имели 6 хромосом, а другие 7, т. е. одна хромосома была непарной.
Ее назвали Ххромосомой, а все другие аутосомами (А). В соматических клетках самцов
содержится 13 хромосом, одна из которых Ххромосома. Таким образом, сперматозоиды самца
могут быть двух типов и содержать 6А+Х либо 6А хромосом.
1В соматических клетках самок имеется 14Х(12А+2Х) и соответственно образуются
яйцеклетки одного типа – 6А+Х. В сперматозоидах у другого клопа (Lygaeus turcicus) хромосомы
одной из пар различались по форме и размеру. При том одна из хромосом была похожа на пару
хромосом женского пола, и за ней оставили название Ххромосомы, а другая, негомологичная
хромосома получила название Ухромосома.
При напечатании в схемах Ххромосому иногда обозначают прямой чертой, а Ухромосому
– неполной стрелкой, т. е. схему скрещивания можно записать как ХХ ХУ.
Цитологическое изучение хромосомных наборов мужского и женского пола у многих
организмов показало, что один пол имеет двеодинаковые Ххромосомы, а другой – гетероморфную
пару из Х и У хромосом, либо только одну Ххромосому.
Пол, имеющий пару ХХ, назвали – гомогаметным, т. к.в результате мейоза у него образуются
гаметы одного типа, апол с ХУ или ХО – гетерогаметным.
У разных групп организмов гомо или гетерогаметным может быть различный пол.
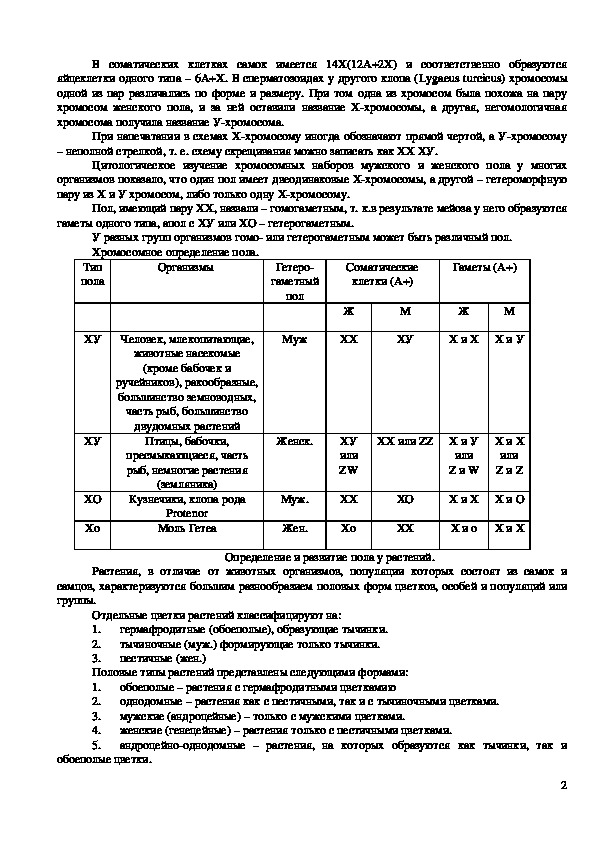
Хромосомное определение пола.
Тип
пола
Организмы
Гетеро
гаметный
пол
Соматические
клетки (А+)
Гаметы (А+)
ХУ
Человек, млекопитающие,
животные насекомые
Муж
(кроме бабочек и
ручейников), ракообразные,
большинство земноводных,
часть рыб, большинство
двудомных растений
Птицы, бабочки,
пресмыкающиеся, часть
рыб, немногие растения
(земляника)
Кузнечики, клопа рода
Protenor
Моль Гетеа
ХУ
ХО
Хо
Женск.
Муж.
Жен.
Ж
ХХ
ХУ
или
ZW
ХХ
Хо
М
ХУ
Ж
М
Х и Х Х и У
ХХ или ZZ Х и У
или
Z и W
Х и Х
или
Z и Z
ХО
ХХ
Х и Х Х и О
Х и о
Х и Х
Определение и развитие пола у растений.
Растения, в отличие от животных организмов, популяции которых состоят из самок и
самцов, характеризуются большим разнообразием половых форм цветков, особей и популяций или
группы.
гермафродитные (обоеполые), образующие тычинки.
тычиночные (муж.) формирующие только тычинки.
пестичные (жен.)
Отдельные цветки растений классифицируют на:
1.
2.
3.
Половые типы растений представлены следующими формами:
1.
2.
3.
4.
5.
обоеполые – растения с гермафродитными цветкамию
однодомные – растения как с пестичными, так и с тычиночными цветками.
мужские (андроцейные) – только с мужскими цветками.
женские (генецейные) – растения только с пестичными цветками.
андроцейнооднодомные – растения, на которых образуются как тычинки, так и
обоеполые цветки.
2генецейнооднодомные (М и Ж).
тримоноцейные – образующие цветки трех типов (М, Ж и МЖ).
6.
7.
В 1906 году в опытах Корренса по скрещиванию мужских и женских растений брионии
двудомной и однодомной было установлено, что женские растения гомогаметны, а мужские
гетерогаметны.
Мох – печеночник стал первым растением, у которого цитологически обнаружили половые
хромосомы. Растения мха гаплоидны, диплоидны спорангий и его ножка. Ален (1917) установил,
что женские и мужские гаплоидные растения мха имеют по 7 одинаковых хромосом, а одна пара
различается у мужских растений маленькая точечная Jхромосома, а у женских длинная Х
хромосома. После оплодотворения и соединения двух гаплоидных наборов развивается сапрофит
14 А+ХУ. После мейоза из одной материнской клетки образуется 4 споры, две из которых
получают набор 7А+Х, а две другие – 7А+У и соответственно развиваются 1:1 женские и мужские
растения в соотношении.
Наследование, сцепленное с полом.
При анализе наследования ряда признаков у раздельнополых организмов выяснилось, что
некоторые из них передаются своеобразно и явно зависят от пола.
В 1909 г. Т. Морганом с сотрудниками было доказано, что такие признаки находятся в
половых хромосомах.
Изучая наследование признаков Т. Морган установил у дрозофилы наличие связи
определенных генов с половыми хромосомами и таким образом заложил фундамент хромосомной
теории наследственности. Кроме генов, определяющих пол, половые хромосомы содержат гены,
влияющие на разные признаки, не имеющие отношения к дифференциации пола. При передаче
таких генов и наблюдается явление так называемого наследования, сцепленного с полом.
В своих первых опытах Морган использовал мутацию белых глаз. Дикие дрозофилы имеют
красные глаза. Красный цвет глаз доминирует над белым.
При скрещивании гомозиготной красноглазой самки с белоглазым самцом в первом
поколении все мухи имели красные глаза, а во втором поколении происходило расщепление в
равном соотношении на красноглазых и белоглазых мух только среди самцов, а все самки во втором
поколении были красноглазые. Соотношение по полу 1:1.
Р Ж красн. Х М белогл.
F1 все красногл.
F2 Ж красногл., ½ М красн. И ½ М белогл.
Если М красн. Х Ж белогл.
F1 Ж красн., М белогл.
F2 ½ Ж красн. И ½ Ж белогл.
½ М красн. И ½ М белогл.
В реципрокном скрещивании результаты отличались тем, что уже в F1 наблюдалось
расщепление по цвету глаз, причем все самки были красноглазые, а самцы белоглазые, т. е. похожи
на матерей. Такое наследование называется крисскросс, т. е. крест –накрест. В F2 такое
расщепление становится понятным, если допустить, что гены, определяющие окраску глаз,
находятся в Ххромосоме, а в Ухромосоме их нет.
3