Поделиться

Содержание
стр.
Введение…………………………………………………………………………...3
Аннотация…………………………………………………………………………5
Глава 1. Радиоактивное загрязнение растений………………………………….6
1.1.Поступление РН в растения в зависимости от их физико-химических свойств……………………………………………………………………………..6
1.2. Поступления РН в растения из разных типов почв………………………...8
Глава 2. Влияние радионуклидов на растения в лесах………………………..13
2.1. Радиорезистентность высших растений…………………………………...13
2.2. Чувствительность растений к хроническому облучению………………..14
2.3. Влияние радионуклидов на популяции растений………………………...16
2.4. Генетические последствия облучения растений………………………….17
2.5. Радиоэкология горелого леса………………………………………………19
Глава 3. Лесная радиология……………………………………………………..21
Заключение……………………………………………………………………….24
Список литературы………………………………………………………………25
Введение
Лес - одна из наиболее чувствительных к ионизирующей радиации природных экосистем. Его поражение наблюдается даже при небольших дозах облучения, по сравнению с естественными и культурными биогеоценозами других типов. Особенно неустойчивы к ионизирующей радиации хвойные леса. Лиственные леса более устойчивы к облучению, чем хвойные. В ряду деревья - кустарники - полукустарники - травы устойчивость к облучению повышается. Населяющие лес млекопитающие и птицы по чувствительности к ионизирующим излучениям близки или даже превосходят древесные растения (в том числе хвойные).
Самые радиоустойчивые организмы в лесу – это мелкие беспозвоночные, обитающие в подстилке и верхних слоях почвы, водоросли, мхи и лишайники, микроорганизмы. Они выдерживают без признаков лучевого повреждения дозы в 1000-10000 Гр. Дождевые черви, пропускающие через кишечник почву, содержащую радионуклиды, оказываются наиболее чувствительными организмами среди беспозвоночных [1].
Степень лучевого повреждения определяется поглощённой дозой в различных компонентах леса – прежде всего в древесном ярусе – фазой развития растений (весной и летом, то есть в период активного роста, древесные растения повреждаются в 1,5-2 раза сильнее, чем осенью и зимой), экологическими факторами (например, засуха способствует усилению поражающего влияния ионизирующих излучений на лес).
Прекращение облучения или снижение мощности дозы приводит к пострадиационному восстановлению лесного биогеоценоза. Небольшие дозы ионизирующих излучений оказывают стимулирующее действие на растения (облучение семян древесных растений применяют для ускорения их роста и развития), более высокие - мутагенное, что используется в радиационном мутагенезе древесных и кустарниковых пород.
Источником облучения леса могут быть радиоактивные вещества, поступающие в биосферу в результате ядерных взрывов, аварий на предприятиях атомной промышленности, а также с радиоактивными отходами, сбрасываемыми в окружающую среду.
Наибольшую опасность представляют биогенно-подвижные радионуклиды 90Sr (стронций), 129I (йод), 137Cs (цезий) и др. Распределение радионуклидов в лесу зависит от их физико-химических свойств и путей поступления радиоактивных веществ в лесные насаждения, а также от типа леса и структуры насаждений [8].
При поступлении радионуклидов из воздуха основная масса их первоначально задерживается на поверхности крон и стволов деревьев, а позднее - в лесной подстилке, на поверхности почвы. Самоочищение надземных частей деревьев от радиоактивных веществ происходит медленнее в хвойных лесах, где «фильтр» из хвои действует круглогодично, чем в лиственных, где важное значение имеет листопад. Основное количество радионуклидов в лесу локализуется в растениях, роль животных в этом отношении менее существенна.
Лиственные породы накапливают больше радионуклидов, чем хвойные. Повышенные концентрации радиоактивных веществ характерны для лесной подстилки, верхних горизонтов почвы, зелёных частей древесных и травянистых растений. Концентрация радионуклидов в древесине, незначительна. С пищевыми продуктами из леса (мясо промысловых животных, грибы, ягоды) радионуклиды могут поступать в организм человека.
Цель работы: Изучить влияние радиоактивного излучения на лесную растительность.
Задачи:
1. Выявить причины и последствия радиоактивного загрязнения растений.
2. Рассмотреть и проанализировать влияние радионуклидов на растения в лесах.
Аннотация
В работе рассмотрены вопросы влияния радиации на лесные сообщества, дана характеристика радиоактивного загрязнения растений, рассмотрены пути загрязнения, поступления РН в растения из разных типов почв, физико-химические свойства радионуклидов. В работе выявлено, что загрязнение растений радиоактивными веществами зависит от специфики слоя почвы, то есть от типа и свойств почв, на которых растут растения. Дана характеристика механического состава различных лесных почв, минеральной части почвы, кислотности почв, карбонатности и влажности почв. В работе рассмотрены вопросы влияния радионуклидов на растения в лесах. Изучена чувствительность растений к хроническому облучению, генетические последствия облучения растений, влияние радионуклидов на популяции растений, вопросы и значение лесной радиологии.
В работе использованы источники литературы современных авторов и ученых по теме радиационная экология, радиобиология, в трудах ученых рассмотрены вопросы действия ионизирующих излучений на экологические системы, миграция радионуклидов в лесных биогеоценозах и другие.
В данной работе даны такие определения и понятия, как: радиология, нуклеотиды, радиоактивное загрязнение, радиоактивные частицы, радиочувствительность, радиоэкологические исследования, мутагенное действие излучения, радиорезистентные растения, ионизирующее излучение и другие.
Работа состоит из введения, трех глав, заключения и списка использованной литературы, включает 2 рисунка.
Глава 1. Радиоактивное загрязнение растений
1.1.Поступление РН в растения в зависимости
от их физико-химических свойств
Радиоактивное поражение растений проявляется в торможении роста, снижении урожайности, репродуктивной качества семян, а при больших дозах вызывает гибель растений.
Растения могут загрязняться двумя путями: аэрозольным (не корневой путь) и корневым (почвенный путь поступления).
Особенность не корневого пути поступления заключается в том, что при непосредственном осадке радиоактивных частиц из различных слоев атмосферы происходит загрязнение надземной массы растений всеми РН, что выпадают [2].
Радиоактивные частицы не полностью задерживаются на растениях. Степень задержания радиоактивных частиц растениями характеризуется величиной первоначального задержания отношением количества осевших на растениях радиоактивных частиц от общего их количества, которая выпала из атмосферы на данной площади.
Коэффициент первичного содержания Кпу = ξГ/ξВ, Дж
где ξ,в - плотность выпадений (количества радиоактивности, что выпала на единицу площади посева или травостоя);
ξ,г - плотность радиоактивного загрязнения надземной массы растений (количества радиоактивности в надземной массе с единицы площади посева).
Различные лесные и сельскохозяйственные культуры имеют неодинаковую способность к содержанию радиоактивных осадков из атмосферы, что обусловлено как спецификой морфологического строения растений, так и степенью развития надземной массы.
Коэффициент первичного содержания Кпу может изменяться в очень широких пределах - от нескольких до 95%.
Неодинаковой способностью к содержанию радиоактивных осадков характеризуются не только различные виды сельскохозяйственных и лесных культур, но и различные части и органы одного и того же растения.
Содержимое РН в единице массы зерна зависит от сроков их выпадение [8].
Выпадение аэрозольных частиц 90Sr из атмосферы на некоторые растения очень опасное. При выпадении из атмосферы 137Cs не только механически загрязняет леса, но и интенсивно проникает в ткани наземных органов растений, включается в метаболизм, перемещается внутри растения и накапливается в урожае. Достаточно интенсивно движется внутри растения при попадании на ее поверхность 131И.
Механизм усвоения РН корнями растений для поглощения основных питательных веществ, макро - и микроэлементов.
Отличие заключается в том, что в основном РН во внешней среде имеется в предельно низких концентрациях. Например, стронция 90 содержится 1,4 х 10 - 12г х кг - 1 почвы, а масса 1 Ки 90Sr составляет 7 х 10 – 3 г.137Cs является химическим аналогом калия, 90Sr - кальция, поэтому наблюдается определенное сходство поглощения растениями и передвижения по ним К, Са и их химических аналогов Cs и Sr.
Больше всего поглощается растениями с питательного раствора 137Cs, значительно меньше - 90Sr. Таких РН, как 60Со, 106Ru, 144Ce, 147Рт, поступает из водного раствора в наземную массу растений в 10 раз меньше, чем Cs и Sr.
В целом накопления РН и их содержимое на единицу массы сухого вещества в процессе роста растений имеет такую же закономерность, как и накопления биологически важных элементов: с возрастом растения в надземных органах увеличивается абсолютное количество РН и снижается содержание на единицу массы сухого вещества [3].
1.2. Поступления РН в растения из разных типов почв
Загрязнение растений зависит от специфики слоя почвы, то есть от типа и свойств почв, на которых растут растения.
Среди физико-химических характеристик грунта выделяют 10 параметров, что считают наиболее значимыми при определении поведения радионуклидов в почве и перехода их в растения. В общем виде влияние почвы проявляется в снижении биологической подвижности радионуклидов при увеличении содержания в почве обменных катионов, органического вещества, ила, минералов, емкости поглощения. Направленность влияния кислотности, карбонатов и влажности почв зависит от физико-химических свойств радионуклидов. В количественном отношении влияние различных свойств почвы проявляется в разной степени. Для большинства накопления радионуклидов в растениях в зависимости от изменения почвенных характеристик может колебаться в среднем в 10-20 раз, а для некоторых радионуклидов в 100-200 раз [2].
Различные типы почв имеют закономерное сочетание основных почвенных показателей. Так, черноземы характеризуются повышенным содержанием гумуса, обменных катионов, большой емкостью поглощения, повышенным содержанием минералов, а дерново-подзолистые почвы, наоборот, отличаются небольшим количеством питательных веществ, незначительной емкостью обмена, низким рН, низким содержанием гумуса.
В связи с этим выявляется зависимость между поступлением радионуклидов в растения и типом почвы. Накопление радионуклидов растениями снижается в ряде почв: аллювиально-шаровые, дерново-подзолистые, серые лесные, сероземы, каштановые, черноземы.
Емкость катионного обмена и содержимое обменных катионов. Емкость катионного обмена характеризует способность почвы сорбировать катионы. Радионуклиды, сорбированные в почве по обменному типу, являются наиболее доступными для растений. Прочность сорбции радионуклидов в почве тем выше, чем больше емкость катионного обмена.
Рядом с обменным поглощением радионуклидов почвой существует необменные поглощения - фиксация.
Емкость катионного обмена и сумма обменных катионов в почве меняются в зависимости от реакции среды, состав органических и минеральных компонентов и природы грунтовых минералов. Увеличение щелочности почв способствует росту их емкости поглощения. Вследствие высокой поглотительной способности гумусовых кислот (до 400-930 мг экв/100 г), содержащие большое количество органического вещества и минералов, отличаются наибольшей емкостью поглощения (черноземы, черноземно-луговые почвы) [3].
Эти почвы характеризуются наиболее высокой сорбции радионуклидов и минимальной их биологической подвижностью.
Механический состав почв. Сорбционная способность почв возрастает с увеличением степени дисперсности грунтовых частиц мелкодисперсных фракции имеют высокую способность по сравнению с крупными фракциями.
В пределах одного типа почв в зависимости от гранулометрического состава накопления радионуклидов растениями может изменяться в 10 раз.
Более высокая сорбция радионуклидов мелкими фракциями почвы обусловлена как большой удельной поверхностью глинистых и илистых частиц, так и различием их свойств.
С уменьшением размера гранулометрических фракций повышается содержание в них гумуса и обменных катионов, а также увеличивается емкость катионного обмена.
Кроме того, различия в свойствах гранулометрических элементов различной степени дисперсности объясняется особенностями минералогического состава. Предпочтительными минералами фракций мелкого песка и крупной пыли является кварц и полевые шпаты, в средней и мелкой пыли увеличивается содержание слюд и гидрослюд. В илистой фракции преобладают слюды, гидрослюды и минералы [4].
Минеральная часть почвы. Минеральная часть грунта составляет от 55 до 97 % массы грунта. В почве присутствуют первичные минералы, представленные крупнопесчаными долями, и вторичные минералы, которые преобладают в глинистых и коллоидных фракциях почвы.
В группу первичных минералов относятся кварц, апатит, полевые шпаты, слюды (мусковит, биотит) и др., к вторичным - минералы группы каолинита, вермикулита, вторичных гидрослюд и др. Вторичные минералы отличаются высокой поглощаемостью [1].
Органическое вещество почвы. Важное влияние на миграцию радионуклидов в почве и поглощения их растениями создает органическое вещество. Для большинства радионуклидов увеличение содержания гумуса в почве является фактором, снижающим их поступление в растения [4].
Поведение радионуклидов связано с органическим веществом почв специфической природы - гуминовыми и фульвокислотами. Способность гуминовых кислот адсорбировать ионы, а также образовывать прочные сложные комплексы с радионуклидами влияет на сорбцию их в почве и поступление в растения.
Ряд радионуклидов, например трансурановых, характеризуется высокой мобильностью. Исключительная роль органического вещества в поступлении в растения радиоизотопов вследствие образования йод-гумусовых соединений.
Кислотность почвы. Кислотность почв неоднозначно влияет на биологическую подвижность в них радионуклидов. Для 90Sr, 137Cs при увеличении кислотности возрастает интенсивность поступления радионуклидов в растения. При повышении рН 59Fe, 60Co, 65Zn и 115mCd переходят из ионной формы в разные гидролизные и комплексные соединения, что снижает их доступность для растений. Для многих радионуклидов зависимость поведения от кислотности почв сложная - для некоторых из них характерны два и более пики максимумов подвижности [1].
Кислотность влечет и косвенное влияние на сорбцию грунтами радионуклидов, изменяя емкость катионного обмена.
Карбонатность почвы. Влияние карбонатности почв на доступность радионуклидов растениям связана с изменением кислотности, соотношением различных фракций органических веществ и состав обменных катионов, в частности, содержания обменного Са. Увеличение карбонатности почв снижает накопление 90Sr в сельскохозяйственных и лесных культурах в 1,1-3 раза, а для 137Cs в этих же условиях поступление в растения увеличивается в 1,3-4 раза.
Влажность почвы. Сведения о роли почвенной влаги в миграции радионуклидов в системе почва - растения довольно противоречивы.
Отмечено увеличение перехода радионуклидов в растения с ростом содержания влаги в почве, так и отсутствие влияния влажности. При различных режимах увлажнение может расти общий вынос радионуклидов за счет увеличения биомассы растений [1].
Неоднозначность имеющихся данных о роли влажности связана с тем, что различные радионуклиды поглощаются растениями из почвы в зависимости от режима ее увлажнения по-разному. Кроме того, влияние влажности почв на подвижность радионуклидов зависит от свойств почв и биологических особенностей растений.
Распределение радионуклидов в профиле почвы. Распределение радионуклидов в пределах корнеобитаемого слоя почвы влияет на их поступление в растения. В первый период после попадания радионуклидов на грунтово-растительный покров с воздуха они локализуются в верхней части почвы. Много радионуклидов характеризуются слабой подвижностью и в течение длительного периода времени задерживаются в слое почвы 0-5 см. Наиболее интенсивно переходят в растения радионуклиды при сосредоточении их в луговой дернине. Так, в первый год после внесения 90Sr на дернину его содержание в растениях будет в 2 - 8 раз выше, чем на 3-й год [7].
Равномерное размещение радионуклидов в пахотном слое при вспахивании снижает переход их в растения [7].
Наиболее высокие уровни загрязнения стронцием наблюдаются на дерново-подзолистых почвах, меньше - на серых лесных почвах и сероземах и самые низкие - на черноземах. Аналогичная зависимость установлена и для цезия.
Большое разнообразие почв в нашей стране определяет значительную разницу в поведении РН в почвах и накоплении их в растениях. Поэтому концентрация РН в растениях на различных почвах в различных ґрунтово-климатических зонах страны при одном и том же уровне загрязнения может отличаться в 10 раз [8].
Существует прямо пропорциональная зависимость между плотностью загрязнения местности и накоплением РН в растительной продукции.
Размер накопления РН в растениях зависит от их видовых и сортовых особенностей. Растения, которые содержат больше кальция, накапливают 90Sr больше, а растения, отличающиеся высоким содержанием калия, накапливают больше 137Cs.
Глава 2. Влияние радионуклидов на растения в лесах
2.1. Радиорезистентность высших растений
Все высшие растения, диплоидное и полиплоидные эукариоты, относятся к IV кариотаксону, что отличается максимальной надежностью генома, которая составляет около 105 эв. Такая надежность генома обусловлена интенсивными процессами репарации ДНК, поэтому благодаря резко повышенной по сравнению с микроорганизмами содержания ДНК в ядре их радиочувствительность не столь высокой. В зависимости от содержания ДНК в ядре (который варьирует в них от 10 до 1000 пг) Do для высших растений колеблется от единиц до сотен грей [4].
Однако это показатель радиорезистентности клеток высших растений. Чтобы вызвать гибель ростков, а тем более взрослых растений, нужны поглощенные дозы излучения в десятки и сотни раз больше, особенно в случае хронического воздействия [6].
Некоторые представления о радиорезистентность высших растений при хроническом облучении дают эксперименты на гамма-полях, в том числе с размещением гамма-источников в лесах. Такие работы проводились в Великобритании (А. Воробья), США (Г. Вудвелл), Пуэрто-Рико (Г. Одум и Г. Пиджин), России (М. В. Тимофеев-Ресовський).
Эти исследования доказали, что наименее радиорезистентными из высших растений являются хвойные, прежде всего сосна: иглы ее начинают желтеть, а затем опадают уже по мощности поглощенной дозы излучения около 0,1 Гр/сутки (10 советов/сутки) или 35 Гр (3500 советов) в год. Почти такую же чувствительность к облучению проявляют сосны в случае выпадения радионуклидов. Так, для аварии на Южном Урале в 1957 г., а также на Чернобыльской АЭС характерный «рыжий» лес - большие массивы соснового леса с хвоей. Опадание хвои - следствие влияния не столько инкорпорированного, сколько внешнего облучения (например, от радиоактивного облака, возникшей во время аварии и содержала много короткоживущих радионуклидов).
Корневая система деревьев в почве, что сорбируется основную массу радионуклидов, пострадала относительно мало, как и клетки камбия, что способны пробуждаться и давать новые ростки с нормальной хвоей [6].
2.2. Чувствительность растений к хроническому облучению
Для того чтобы дерево или кустарник погибли в результате радиоактивного загрязнения, нужно не только инактивировать часть клеток их точек роста, т.е. кончиков корней и побегов, но и уничтожить часть клетки камбия, способных пробуждаться и давать начало новым точкам роста. Благодаря такому защитному барьеру кривые выживаемости деревьев и кустарников имеют обычно большие плечи и наклонности, обусловленные уровнем Dо для их клеток, вследствие чего они становятся подобными кривых выживаемости простейших. Поэтому сведения об уровне Do в случае облучения деревьев и других растений дают значительно заниженные оценки их радиорезистентности [6].
Многочисленные эксперименты с острого и хронического облучения семян различных растений показали, что предельной дозой, ниже которой в каких растений не удается наблюдать других последствий облучения, кроме лучевого стимулирования, является доза 10 Гр., что примерно соответствует поверхностной активности радионуклидов 3,7 х 1014 Бк/км2 (104 Ки/км2) в пересчете на хроническое облучение в течение года. Эта оценка учитывает результаты изучения растений и в лабораторных экспериментах, и в природных условиях.
Наиболее чувствительны к воздействию радионуклидов является семена, ростки и деревья сосны, другие древесные значительно устойчивее, еще более радоорезистентни кустарники и травы. Поэтому травы (по крайней мере некоторые виды их) могут выдерживать хроническое облучение дозой до 10 Гр/сутки, что соответствует поверхностной активности радионуклидов на загрязненной территории до 3,7 х 1014 Бк/км2 (104 Ки/км2) и более.
Хронический влияние градиента гамма-излучения на дубово-сосновый лес показано на рис. 1. Мощность дозы 0,02 - 0,05 Г/сутки или 2 - 5 советов/сутки (что составляет 10 - 20 Гр/год, или 1000 - 2000 советов/год), не влияет на растения такого леса. В случае мощности дозы около 0,1-0,2 Гр/сутки (10 - 20 советов/сутки) начнет реагировать сосна, в которой впоследствии буреет и опадает хвоя [6].
Мощность поглощенной дозы излучения 0,05-1 Гр/сутки, або5-100 рад/сутки (то есть 10 - 3,5 • 102 Гр/год), приводит к снижению видового разнообразия леса и увеличение количества насекомых-паразитов, что предпочитают подавленным растениям. Мощность поглощенной дозы излучения, которая превышает 1 Гр/сутки (100 рад/сутки), или 3,5 • 102 Гр/год, приводит к гибели высших рослый, которые практически вымирают при 5 Гр/сутки (500 советов/сутки), или 3,5 • 104 Гр/год.
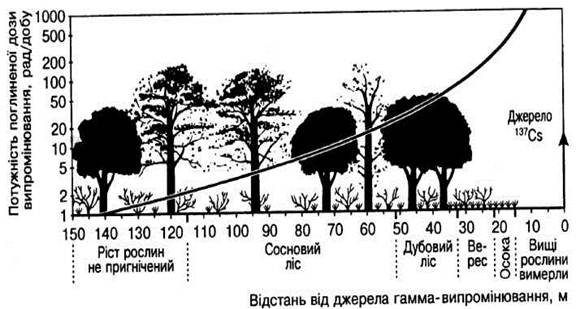
Рис. 1. Влияние градиента хронического гамма-излучения на дубово-сосновый лес (за И.А. Одумом, 1975)
Наиболее радиорезистентними из лесных растений есть мелкие кустарники и травы, такие как верес, осока и другие, в которых значительная часть растения размещена под землей и после прекращения облучения дает корневые ростки, что быстро отрастают. Рекордсменом в этом отношении является травянистое растение Panicum sanguinale, которое образует в случае хронического облучения по мощности поглощенной дозы 10 Гр/сутки (1000 советов/сутки) вместо обычной смеси многих видов разнотравья и злаков чистый травостой [6].
2.3. Влияние радионуклидов на популяции растений
Таким образом, можно обоснованно считать, что поверхностная активность радионуклидов территории примерно до уровня 3,7 • 103 Бк/км2 (103 Ки/км2) никак не будет влиять на проживание на ней микроорганизмов и растительность. Это нижняя граница радиорезистентности этих организмов.
На самом деле их радиорезистентность еще выше.
Во-первых, эффект хронического облучения намного меньше, чем острого (а приведенный выше материал касается в основном острого облучения).
Во-вторых, при использовании данных о радионуклеотидном загрязнение мы учитывали только внешнее облучение от радионуклидов, внесенных в почву или воду, а не дополнительное внутреннее облучение за счет инкорпорированных радионуклидов [8].
Коэффициенты перехода и накопления могут быть очень значительными, и дозы, полученные растениями вследствие действия радионуклидов, должны быть существенно больше, чем было учтено нами во время рассмотрения влияния радионуклидов в растения (как и на животных, живущих на загрязненных ими территориях).
Фактически человека интересует не столько характер изменений в состоянии здоровья отдельных представителей биоты, живущих на загрязненных территориях, сколько то, какой вред может причинить такое загрязнение популяциям этих организмов, то есть, как повлияет их облучение на способность давать полноценное потомство. Таких данных уже накоплено достаточно, чтобы считать, что популяции микроорганизмов и высших растений более устойчивы к облучению (особенно хронического), чем их отдельные особи.
Учитывая такой популяционный аспект радиоэкологических исследований можно считать вполне обоснованным уровень мощности поглощенной дозы излучения 10 Гр/год или поверхностной активности радионуклидов 3,7 • 1013 Бк/км2 (103 Ки/км2) как нижнюю границу биологически безвредного облучения не только микроорганизмов и растений. При дозах, не превышающих приведенных значений, никаких неблагоприятных последствий для растений ожидать не следует. Независимо от такого облучения растения и микроорганизмы будут нормально выполнять свои трофические функции, иметь такую же производительность и способность, как и без облучения, и играть ту же роль концентраторов и транспортеров радионуклидов [2].
Такое кондиционирования окружающей среды от радионуклидов, которые попали в него, более чем на 99% выполняют именно высшие растения и микроорганизмы, и только около 1% приходится на животных.
2.4. Генетические последствия облучения растений
Генетические последствия облучения микроорганизмов имеют универсальный характер для всей биоты. Относительно генетических последствий облучения, то имеются в виду мутации генов, ведь довольно значительные мутации хромосом летальные для клеток, и закономерности их возникновения в результате облучения такие же, как и закономерности его летальной действия [6].
Нельзя считать мутации генов однотипными. Кроме уже отмеченного разнообразия их структурной основы - от точечных мутаций, обусловленных поражением малого количества основ, к структурным, которые захватывают много тысяч основ, нужно учитывать, что подобные фенотипические последствия могут иметь изменения генов вследствие внутренних рекомбинаций; частота же рекомбинаций может в 10-30 раз превышать частоту настоящих мутаций генов.
Таким образом, под генетическими последствиями облучения понимают интегральный результат, что проявляется в изменении потомства облученных особей независимо от того, вызвано это мутацией генов (точечной или структурной) или хромосом.
Рассмотрим два аспекта мутагенного действия излучения: зависимость ее от мощности дозы и ЛПЭ излучения, а также от уровней интегральной поглощенной дозы излучения. На основании различных данных, полученных как на микроорганизмах, так и на высших растениях, установлено, что мутагенный эффект облучения, отнесенный к единице поглощенной дозы излучения (например, частота мутаций на 1 Гр), с увеличением мощности поглощенной дозы сначала несколько уменьшается, а затем начинает расти.
Механизм такой закономерности и локализация «точки перегиба» на оси мощности дозы окончательно не выяснены. По этому поводу существует лишь одна гипотеза, что связывает характер изменений мутагенного эффекта облучения с индукцией в объектах системы восстановления (SOS-системы), подвергшихся облучению, и обусловленным с этим восстановлением клеток от передмутационных изменений ДНК [5].
На основании общих понятий, объясняют эмпирически установленный факт увеличения генетического эффекта облучения с увеличением ЛПЭ излучения. Однако, механизм этого феномена еще не выяснен.
Относительно количественной зависимости частоты индуцирования мутаций от поглощенной дозы излучения являются данные в основном для одноразового острого облучения в достаточно больших дозах - 1 Гр (100 рад) и больше. В случае хронического воздействия обычно экспериментируют с поглощенными дозами 1 Гр (100 рад) и более на поколения, потому что уменьшение дозы ниже этого уровня предопределяет необходимость столь увеличивать объем выборки (для получения достоверных данных), что это делает невозможным исследования [6].
Можно считать, по мощности поглощенной дозы 0,1 - 1 Гр/год в случае хронического облучения уровень регистрируемых генетических изменений статистически не будет отличаться от контроля. Это утверждение касается и микроорганизмов, и высших растений, и большинства животных. Поэтому такую мощность поглощенной дозы в случае хронического облучения 0,1 - 1 Гр/год можно, считать относительно генетических последствий безопасной для любых представителей биоты.
Косвенным подтверждением этого является то, что в случае возникновения при таком облучении и с низкой частотой мутантных особей (дополнительно к спонтанному фона) они, будут объединяться в популяции в результате отбора, известно, что подавляющее большинство спонтанных и индуцированных мутантов отличаются от особей дикого типа (выделенные из природных условий без отбора) несколько пониженной жизнеспособностью. Мутанты с повышенной жизнеспособностью, подвергающихся отбору, будут привлекаться к участию в жизни популяции, не причиняя ей никакого вреда [1].
2.5. Радиоэкология горелого леса
Крупные пожары в 30-километровой зоне ЧАЭС в 1990-1992 гг. охватили площадь (2000-3000 га) загрязненного радионуклидами леса.
Процесс верховой (по кронам деревьев) и низового пожара привел к выгоранию сухой, загрязненной радионуклидами лесной подстилки, которая содержит до 90 % радионуклидов, выпавших на лесные массивы зоны.
Горение лесной подстилки обусловило образование огромного количества аэрозолей, что вызвало интенсивное вторичное ветровое подъема в воздух больших количеств радионуклидов (коэффициент ветрового подъема в горящем лесу достигает 10 -2 м -1). Радионуклиды с аэрозолями и пеплом переносятся на достаточно большие расстояния от мест пожаров.
Так, на полигоне «Буряковка» после массового возгорания лесов радиационный фон на расстоянии 3-5 км повысился в 1,5 раза; возросли также и уровне поверхностного загрязнения почвы [6].
Выгоревший лес - это «прозрачное» скопления голых стволов, стоящих среди черной выгоревшей лесной подстилки. Понятно, что горела лесная подстилка среди стволов, практически лишенных кроны, приводит к резкому увеличению (в 3-5 раз) поверхностного твердого и жидкого стока. Основные пожары в зоне были осенью, и закрепление горелой подстилки свежим травяным покровом не произошло. Поэтому до появления снежного покрова в зоне ЧАЭС в последующие годы весной можно было ожидать повышенного стока радионуклидов по всей территории пожарищ [6].
Восстановление травяного покрова на местах пожарищ длится 3-5 лет, а древесной растительности более 15 - 30 лет. В этот период способность горелого леса удерживать радионуклиды резко уменьшается.
При значительных площадях горелого леса и высоком уровне их загрязнения радионуклидами возможное повышение стока радионуклидов из ландшафта пожарищ в водотоки, в г. Припять и далее в каскад Днепровских водохранилищ.
По оценкам авторов, вследствие пожаров удельная радиоемкость лесных массивов уменьшается в 35 раз. Поэтому для лесов, значительно загрязненных радионуклидами, особенно в 30-километровой зоне ЧАЭС, а также в Украине и Беларуси, следует предусмотреть эффективную систему противопожарных мероприятий. Горения таких лесов может заметно повысить загрязнения воздуха и таким образом потенциально увеличить составляющую дозы излучения для населения поселков и сел за путем ветрового переноса аэрозолей и пепла. Много важных проблем радиоэкологии горелого леса еще требуют детального исследования [7].
Глава 3. Лесная радиология
Раздел радиобиологии, изучающий действие ионизирующих излучений на лес и миграцию радиоактивных веществ (радионуклидов) в лесных биогеоценозах, основная задача которой состоит в исследовании круговорота естественных и искусственных радионуклидов в лесах и влияния ионизирующих излучений, как на отдельные компоненты, так и на лес в целом [3].
Ранние признаки радиационных повреждения хвойных пород, обладающих крупными хромосомами (мишенями радиационного воздействия), обнаруживаются уже при дозах 2-3 Гр (грей - единица поглощённой дозы ионизирующего излучения, соответствует энергии в 1 Дж, поглощённой 1 кг ткани).
Рис.2. Структурная схема миграции в берёзовом лесу.
Степень лучевого повреждения определяется поглощённой дозой в компонентах леса, прежде всего в древесном ярусе, фазой развития растений (весной и летом, т. е. в период активного роста, древесные растения повреждаются в 1,5-2 раза сильнее, чем осенью и зимой), экологическими факторами (например, засуха способствует усилению поражающего влияния ионизирующих излучений на лес). Прекращение облучения или снижение мощности дозы приводит к пострадиационному восстановлению лесного биогеоценоза. Небольшие дозы ионизирующих излучений оказывают стимулирующее действие на растения (облучение семян древесных растений применяют для ускорения их роста и развития), более высокие - мутагенное, что используется в радиационном мутагенезе древесных и кустарниковых пород.
Источником облучения леса могут быть радиоактивные в-ва, поступающие в биосферу в результате ядерных взрывов, аварий на предприятиях атомной промышленности, а также с радиоактивными отходами, сбрасываемыми в окружающую среду. Наибольшую опасность представляют биогенно-подвижные радионуклиды 90Sr, 129I, 137Cs и др [2].
Распределение радионуклидов в лесу зависит от их физико-химических свойств и путей поступления радиоактивных веществ в лесные насаждения, а также от типа леса и структуры насаждений. При поступлении радионуклидов из воздуха (наиболее значимый источник радиационного поражения лесов) основная масса их первоначально задерживается на поверхности крон и стволов деревьев, а позднее - в лесной подстилке, на поверхности почвы. Самоочищение надземных частей деревьев от радиоактивных веществ происходит медленнее в хвойных лесах, где «фильтр» из хвои действует круглогодично, чем в лиственных, где важное значение имеет листопад.
Основное количество радионуклидов в лесу локализуется в растениях, роль животных в этом отношении менее существенна. Лиственные породы накапливают больше радионуклидов, чем хвойные. Повышенные концентрации радиоактивных веществ характерны для лесной подстилки, верхних горизонтов почвы, зелёных частей древесных и травянистых растений.
Концентрация радионуклидов в древесине, как правило, незначительна. С пищевыми продуктами из леса (мясо промысловых животных, грибы, ягоды и т. д.) радионуклиды могут поступать в организм человека.
Наиболее важное направление лесной радиологии, основная задача которой состоит в исследовании круговорота естественных и искусственных радионуклидов в лесах и влияния ионизирующих излучений, как на отдельные компоненты, так и на лес в целом. Учитывая важное значение ионизирующих излучений как экологического фактора, лесную радиоэкологию можно рассматривать также как раздел экологии. В связи с интенсивным развитием атомной энергетики во 2-й половине 20 в. и необходимостью защиты окружающей среды от радиоактивного загрязнения прикладное значение лесной радиологии особенно возросло. Значительный вклад в изучение лесной радиологии внесли А. А. Молчанов, Г. Ф. Хильми (СССР), А. X. Спарроу, Дж. М. Вудвелл, Р. Б. Платт (США) [8].
Заключение
В результате работы выявлено, что: ранние признаки радиационного повреждения хвойных пород, обладающих крупными хромосомами обнаруживаются уже при дозах 2-3 Гр.
Самые радиоустойчивые организмы в лесу - мелкие беспозвоночные, обитающие в подстилке и верхних слоях почвы, а также водоросли, мхи и лишайники, микроорганизмы.
Принято выделять первичные лучевые реакции леса, непосредственно связанные с воздействием ионизирующих излучений, и вторичные, обусловленные опосредованным действием ионизирующих излучений.
Небольшие дозы ионизирующих излучений оказывают стимулирующее действие на растения (облучение семян древесных растений применяют для ускорения их роста и развития), более высокие - мутагенное, что используется в радиационном мутагенезе древесных и кустарниковых пород.
Источником облучения леса могут быть радиоактивные вещества, поступающие в биосферу в результате ядерных взрывов, аварий на предприятиях атомной промышленности, а также с радиоактивными отходами, сбрасываемыми в окружающую среду.
Распределение радионуклидов в лесу зависит от их физико-химических свойств и путей поступления радиоактивных веществ в лесные насаждения, а также от типа леса и структуры насаждений.
Основное количество радионуклидов в лесу локализуется в растениях, роль животных в этом отношении менее существенна.
Список литературы
1. Алексахин Р. М., Нарышкин М. А., Миграция радионуклидов в лесных биогеоценозах, М., 1977.-342с.
3. Куплахмедов Ю.А. и др. Основы радиоэкологии. - К.: Высшая школа, 2003. - 319 с.
4.Лесная энциклопедия: В 2-х т., т.2/Гл.ред. Воробьев Г.И.; Ред.кол.: Анучин Н.А., Атрохин В.Г., Виноградов В.Н. и др. - М.: Сов. энциклопедия, 2006.- 631 с.
7. Сельскохозяйственная радиоэкология/Под ред. Алексахина Р.М., Корнеева Н.А. - М.: Экология, 2002. - 400 с.
8.Тихомиров Ф.А., Действие ионизирующих излучений на экологические системы, М., 2002. – 276с.










![Равномерное размещение радионуклидов в пахотном слое при вспахивании снижает переход их в растения [7]](https://fs.znanio.ru/d5af0e/76/d5/a0280204ec694708162eb40e4c277e380c.jpg)



![Гр/сутки (1000 советов/сутки) вместо обычной смеси многих видов разнотравья и злаков чистый травостой [6]](https://fs.znanio.ru/d5af0e/cc/30/c3045e8b3506a22678828e9633742b1ae7.jpg)









Материалы на данной страницы взяты из открытых источников либо размещены пользователем в соответствии с договором-офертой сайта. Вы можете сообщить о нарушении.
