До Менделя многие ученые пытались понять закономерности, лежащие в основе наследственности. Однако отсутствие четкой методики исследований гибридных поколений привело к обескураживающим результатам. В одном случае признаки потомков соответствовали признакам одного из родителей, в другом – другого, в третьем проявлялись признаки обоих родителей, а иногда возникали признаки, вообще отсутствующие у родительских форм.
Мендель разработал собственную методологию проведения экспериментов, которая позволила избежать ошибок, присутствовавших в работах предшественников и современников. Ее сущность заключалась в следующем
Тема. Закономерности наследования признаков и принципы наследственности
Лекция 3
План
1. Менделизм. Принципы и методы генетического анализа.
2. Доминантность и рецессивность. Единообразие первого поколения.
3. Правило расщепления гибридов второго поколения.
4. Третий закон Менделя.
5. Условия осуществления Менделеевских законов.
6. Множественный аллелизм.
До Менделя многие ученые пытались понять закономерности, лежащие в основе
наследственности. Однако отсутствие четкой методики исследований гибридных поколений
привело к обескураживающим результатам. В одном случае признаки потомков соответствовали
признакам одного из родителей, в другом – другого, в третьем проявлялись признаки обоих
родителей, а иногда возникали признаки, вообще отсутствующие у родительских форм.
Мендель разработал собственную методологию проведения экспериментов, которая
позволила избежать ошибок, присутствовавших в работах предшественников и современников. Ее
сущность заключалась в следующем:
1. Для скрещивания брались растения, различающиеся по одному признаку или очень
немногим, и учитывалось наследование каждого отдельного признака.
2. При анализах рассматривалось в отдельности каждое из растений и также в отдельности
высевались семена от каждого из гибридных растений.
3. Им был применен математический анализ изучения частоты проявления альтернативных
признаков цветки красные – цветки белые, семена гладкие – семена морщинистые).
Закономерности, выявленные Менделем, легли в основу принципов генетического анализа,
до сих пор используемого в генетических исследованиях.
Приступим к закономерному изложению закономерностей наследования Г. Менделя.
Одним из основных объектов опытов М. был горох. Эта культура – самоопылитель, поэтому,
используя ее, легко в опытах проводить скрещивания, получать гибридное потомство и наблюдать
за ним. Мендель отбирал для своих экспериментов сорта, различающиеся по окраске семенной
кожуры (серая, прозрачная), окраска незрелого боба (зеленая, желтая), длине стебля (длинный,
короткий), типу семян (гладкие, морщинистые), окраске семян (желтые, зеленые), расположению
соцветий (пазушные, верхушечные). Первым его открытием было выявление того, что из 2
альтернативных признаков при скрещивании в первом поколении проявляется только один. Так,
при скрещивании растений с незрелыми зелеными бобами с растением, незрелые бобы которого
были желтыми, в первом поколении у всех растений незрелые бобы были зелеными. Подобные
результаты были получены М. и по другим признакам. Так в F1 появились только такие признаки,
как серая окраска семян кожуры, лущильный тип боба, гладкие семена, желтая окраска семян,
длинный стебель.
Признаки, которые проявились у гибридов в первом поколении, Мендель назвал
доминантами (преобладающими), а альтернативные рецессивными (отсутствующими). Явление
единообразия всех особей первого поколения по изучаемому признаку было названо полным
доминированием. В дальнейшем были обнаружены факты, свидетельствующие о том, что полное
доминирование неуниверсальное явление. Так при скрещивании львиного зева с красными цветками
с линией, образующей белые цветки, все растения в первом поколении сформировали розовые
цветки. Наследование признаков подобного типа было названо неполным доминированием.
Аналогичное наследование было выявлено и у андалузских кур с черным оперением и кур с белым
«забрызганным» оперением все потомки в первом поколении имели голубое оперение.
И, наконец, в потомстве могут одновременно проявляться признаки обоих родителей. Этот
тип наследования получил название кодоминирование. Его примером может служить наследование
групп крови у человека (в системе АВ0). Если один из родителей имеет группу крови А (АА), а
1другой (ВВ), то в крови детей присутствуют антигены, характерные как для группы А, так и для
группы В. Наличие этих антигенов определяют специальной антигенной реакцией. Применительно к
растениям – это наследование различных типов запасных белков, выявляемых методом
электрофореза (глиадины, глютеины и гордеины).
Следует отметить, что явление доминирования, открытое Менделем, не такое простое, как
может показаться на первый взгляд. Было установлено, что в ряде случаев доминирование может
видоизменяться под влиянием внешних условий, возраста, пола, особенностей самого организма и
других, часто не установленных факторов. Так, у дурмана пурпурная окраска доминирует над
зеленой, если растения выращивают в полевых условиях. Однако при выращивании этих же
гибридов в теплице, гибриды первого поколения отличаются значительно более светлой окраской
стебля, чем родительская форма с пурпурным стеблем. Имеется множество и других примеров,
свидетельствующих о случаях видоизменения доминирования. Из вышеизложенного видно, что
самым важным открытием Менделя было установление факта единообразия гибридов первого
поколения. Это явление и получило в дальнейшем название первого закона Менделя. При этом не
важно, имеет ли исследователь дело с фактом полного, либо неполного доминирования или
случаем кодоминирования. Во всех этих вариантах непреложным фактом остается единообразие
особей первого поколения.
Правило расщепления гибридов второго поколения.
Под расщеплением гибридов понимают закономерности распределения среди потомков
второго поколения особей с доминантными и рецессивными признаками.
Высевая семена, собранные с гибридных растений гороха первого поколения, Мендель
получил во втором поколении растения с признаками обоих родителей. Следовательно, в отличие
от первого поколения, второе не было единообразным. Растений с доминантными признаками было
в 3 раза больше, чем с рецессивными. Причем эта закономерность наблюдалась по всем
изучавшимся признакам.
Высевая в дальнейшем семена, собранные с каждого из растений второго поколения,
Мендель обнаружил, что некоторые растения с доминантными признаками в третьем поколении не
расщеплялись, и все их потомство было единообразным. Причем соотношение особей с
доминантными и рецессивными признаками, как и во втором поколении, было 3 : 1. Гибридные
растения второго поколения с рецессивными признаками во всех случаях давали
нерасщепляющееся потомство. Соотношение растений с доминантным признаком, не давших
расщепляющегося потомства, к растениям с рецессивным признаком соответствовало 1 : 2 : 1.
Мендель писал: «…теперь ясно, что гибриды по двум различающимся признакам образуют семена,
из которых половина дает вновь гибридную форму, тогда как другая дает растения, которые
остаются константными и в равных долях содержат доминирующий и рецессивный признаки».
Таким образом, Мендель впервые показал, что внешний вид растения (фенотип) не всегда
отражает наследственные задатки (генотип). Изучая последующие гибридные поколения, Мендель
показал, что от поколения к поколению число гибридных форм уменьшается, а число
неращепляющихся особей возрастает. Явление расщепления во втором и последующих поколениях,
открытое Менделем, приложило к случаям неполного доминирования и кодоминирования. Однако
в этих случаях, потомство второго поколения может быть сгруппировано не в два, а в три класса: ¼
особей с признаками первого родителя (Р1), ¼ с признаками второго (Р2) и ½ потомства составляют
гибридные особи с выраженностью признака, идентичной первому поколению. Особи, несущие
признаки родительских форм, при пересевах не расщепляются.
Искусственное получение гибридов путем гибридизации и факты их расщепления во втором
и последующих поколениях позволили Менделю прийти к выводу, что подобные результаты могут
быть объяснены только, если предположить, что половые гаметы (зачатковые и пыльцевые клетки
по М.) несут постоянные и обособленные единицы наследственности – факторы, названные в 1909г.
В. Иогансеном генами. При этом, по М., эти единицы наследственности в неизмененном виде
передаются от одного поколения к другому. Мендель ввел буквенные обозначения для факторов,
2определяющих альтернативные признаки. При этом фактор, детерминирующий доминантный
признак, он обозначил заглавной буквой латинского алфавита, а рецессивный – строчной. В.
Иогансен (1926г.) предложил именовать отдельный фактор термином «аллель». М. совершенно
правильно считал, что половые клетки содержат только по одному фактору (аллелю), а гибридные
растения по два. Например, сорта гороха с гладкими семенами несут аллели АА, а с морщинистыми
аа. В первом случае каждая из половых клеток несет аллель А+А=АА и а+а=аа. При скрещивании
растений с гладкими семенами с растениями с морщинистыми семенами будет наблюдаться
следующая картина А+а=Аа. Формы, у которых в зиготе объединяются два идентичных аллеля АА
или аа, получили название гомозиготных, а объединяющих два разных аллеля Аа – гетерозиготных.
М. пришел к выводу, что гибридные растения образуют два типа половых клеток (женские и
мужские): один с аллелем А, а другой с аллелем – а. Оба типа половых клеток образуются с
равными частотами и имеют ровные вероятности на соединения в зиготе, что легко представить,
используя решетку Р. Паннета (англ. Генетик, предложивший производить записи в виде решетки).
Так гибрид первого поколения Аа продуцирует гаметы А и а, образующиеся с частотами ½ и ½. В
данном случае решетка Пеннета имеет 4 клетки. Если сверху написать мужские гаметы, а слева
женские, получим:
Ж М
А
а
А
АА
Аа
а
Аа
Аа
В каждой из клеток записыв. результат соединения женской и мужской гамет, т. е. тип
получаемой зиготы. Расщепление по генотипу 1:2:1, по фенотипу 3:1. Прямое доказательство
гипотезы М. о том, что именно половые клетки несут «постоянные и обособленные единицы
наследственности».
Проверить образование того или иного типа гамет можно путем анализирующего
скрещивания, что и используют в генетическом анализе. Анализирующим называется скрещивание
гибридной формы с родительской, рецессивной по изучаемому признаку. Поскольку Р дает гаметы
только одного типа, выявление в потомстве от анализирующего скрещивания двух типов особей
свидетельствует о том, что гибридная форма продуцирует гаметы 2 типов, а их соотношение (1:1) –
о том, что распределение доминантных и рецессивных генов происходит с равной частотой.
М. не знал о существовании хромосом, о локализации в них генов и о механизме клеточного
деления (мейоза). Значительно позже было установлено, что гены, отвечающие за одну пару
признаков находятся в одинаковых точках (локусах) гомологичных хромосом и что в результате
мейоза гомологичные хромосомы расходятся в гаметы. А поскольку одна половая клетка содержит
только одну из 2 гомологичных хромосом, в нее попадает только один из пары генов. При
соединении в зиготе гены смешиваются и в дальнейшем такими же отдельностями передаются
потомкам. Это правило получило название закона чистоты гамет (закон расщепления).
Закон независимого комбинирования – гены, определяющие различные признаки,
наследуются независимо друг от друга.
Тригибридное скрещивание.
Работает также закон независимого комбинирования. Рассмотрим на примере.
Семена первого поколения по фенотипу будут гладкими и желтыми, а цветки красными.
Растения первого гибридного поколения образуют восемь типов гамет в равном соотношении,
поскольку вероятность попадания в гамету доминантного или рецессивного гена из пары
альтернативных признаков равна ½ × ½ × ½ × ½ = 1/8.
Таким образом, мы имеем следующие типы гамет:
1/8 АВС 1/8 АВс
1/8 АвС 1/8 авС
В свою очередь, это определяет большое разнообразие зигот, как это видно из рис. 1.
Признаки материнской формы (Р1) отцовского (Р2)
1/8 Авс 1/8 аВс
1/8 аВС 1/8 авс
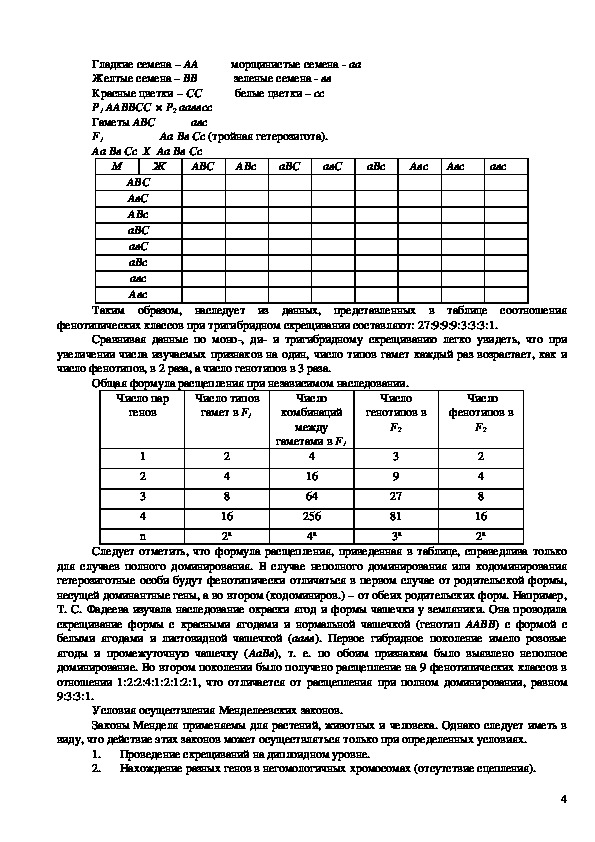
3Гладкие семена – АА морщинистые семена аа
Желтые семена – ВВ зеленые семена вв
Красные цветки – СС белые цветки – сс
Р1 ААВВСС × Р2 ааввсс
Гаметы АВС авс
F1 Аа Вв Сс (тройная гетерозигота).
Аа Вв Сс Х Аа Вв Сс
М
Ж АВС
АВс
аВС
авС
аВс
Авс
Авс
авс
АВС
АвС
АВс
аВС
авС
аВс
авс
Авс
Таким образом, наследует из данных, представленных в таблице соотношения
фенотипических классов при тригибридном скрещивании составляют: 27:9:9:9:3:3:3:1.
Сравнивая данные по моно, ди и тригибридному скрещиванию легко увидеть, что при
увеличении числа изучаемых признаков на один, число типов гамет каждый раз возрастает, как и
число фенотипов, в 2 раза, а число генотипов в 3 раза.
Общая формула расщепления при независимом наследовании.
Число
Число пар
Число
генов
Число типов
гамет в F1
комбинаций
генотипов в
между
гаметами в F1
4
16
64
256
4n
F2
3
9
27
81
3n
Число
фенотипов в
F2
2
4
8
16
2n
1
2
3
4
n
2
4
8
16
2n
Следует отметить, что формула расщепления, приведенная в таблице, справедлива только
для случаев полного доминирования. В случае неполного доминирования или кодоминирования
гетерозиготные особи будут фенотипически отличаться в первом случае от родительской формы,
несущей доминантные гены, а во втором (кодоминиров.) – от обеих родительских форм. Например,
Т. С. Фадеева изучала наследование окраски ягод и формы чашечки у земляники. Она проводила
скрещивание формы с красными ягодами и нормальной чашечкой (генотип ААВВ) с формой с
белыми ягодами и листовидной чашечкой (аавв). Первое гибридное поколение имело розовые
ягоды и промежуточную чашечку (АаВв), т. е. по обоим признакам было выявлено неполное
доминирование. Во втором поколении было получено расщепление на 9 фенотипических классов в
отношении 1:2:2:4:1:2:1:2:1, что отличается от расщепления при полном доминировании, равном
9:3:3:1.
Условия осуществления Менделеевских законов.
Законы Менделя применяемы для растений, животных и человека. Однако следует иметь в
виду, что действие этих законов может осуществляться только при определенных условиях.
1.
2.
Проведение скрещиваний на диплоидном уровне.
Нахождение разных генов в негомологичных хромосомах (отсутствие сцепления).
4Одновременное созревание мужской и женской половых клеток всех типов,
обеспечивающее равновероятное их соединение при оплодотворении.
Отсутствие селективности при оплодотворении гаметами всех типов.
Равновероятная выживаемость мужских и женских гамет всех типов.
Отсутствие селективности и выживаемости зигот всех возможных генотипов.
Проведение экспериментов в условиях, не препятствующих нормальному развитию
4.
5.
6.
7.
8.
3.
Отсутствие у изучаемых организмов нарушений процесса мейоза, а, следовательно, и
равновероятное образование гамет всех возможных типов.
изучаемых признаков.
Обеспечение в эксперименте получения сравнительно большого числа особей.
9.
Таков перечень основных условий, при котором экспериментатор может быть уверен в
отсутствии препятствий в проявлении Менделеевских законов.
Генетические обозначения. Множественные аллели.
Мендель был первым, кто ввел буквенные обозначения для генов. Пока для каждого из
изучающихся видов было открыто небольшое число генов, не имело принципиального значения,
какой буквой обозначался данный ген. Однако с открытием все новых и новых генов при
одинаковом их буквенном значении возникла путаница. В настоящее время генетически чаще всего
обозначался первой буквой английского слова, описывающего признак, который данный ген
контролирует. Различие между аллелями одного гена возникают путем мутаций. Как правило,
доминантный ген мутирует в рецессивный А а. Но известны и обратные мутации: а А. Таким
образом, возникли пары аллельных генов. Например, аллель wx у кукурузы контролирует развитие
восковидного эндосперма, а аллель Wx – нормального крахмального эндосперма. У ячменя
доминантный ген Li определяет развитие лигул у листьев, а его рецессивный аллель li – отсутствие
лигул (безлигульность).
Множественный аллелизм.
Известно много примеров, когда в результате мутирования возникает не 2, а много аллелей
одного гена. Но при этом каждый диплоидный организм всегда несет в генотипе только 2 любых
аллеля одного гена. Классический пример множественного аллелизма – серия аллелей,
определяющая окраску меха у кролика.
Аллель Генотип Фенотип (окраска меха)
С=С+ С+С+;С+СсhС+СhС+Сa серая окраска
Ссh Ссh Ссh шиншилла (пятн.)
Сh Сh Сh; Сh Сa светлая шиншилла
Сa Сa Сa гималайский альбинос (белый)
При этом аллель С = С+ (обозначение дикого типа) доминантен по отношению ко всем
остальным аллелям, аллель Ссh проявляет неполное доминирование по отношению к аллелям Сh, Сa,
поэтому у гетерозиготных особей мех более светлый, чем шиншиллы. Аллель Сh доминантен по
отношению к аллелю Сa.
У ряда видов растений в процессе эволюции выработался механизм, препятствующий
самоопылению. Так, у табаков выявлена серия аллелей гена S: S1, S2, S3, S4, S5...Sn. При этом в
случае попадания на рыльце пестика прорастает только пыльца, не несущая аллелей, идентичных
аллелям генотипа данного растения, а пыльца, несущая ген S3, S4, S5 прорастает свободно, в
результате чего происходит оплодотворение и завязывание семян.
В ряде случаев некоторые из серий множественных аллелей влияют на комбинативное
проявление признака на различных частях организма. Примером таких аллелей является серия
аллелей гена R, влияющего на проявление антоциана у кукурузы:
Rr – растение окрашено, алейрон ограничен.
Rg – растение не окрашено, алейрон окрашен.
rr растение окрашено, алейрон не окрашен.
rg – растение и алейрон не окрашены.
5Присутствие окраски – признак доминирующий над ее отсутствием. Однако растение с
генотипом Rg rr окрашено, и алейрон окрашен, как это имело место у особей с генотипом Rr Rr или
Rr – подобный тип доминирования получил название мозаичного доминирования.
6