В данной работе описано понятие ДНК и самой генетики. Так же объясняется принцип действия генов. Так же определение генов на клеточном уровне. Данная работа знакомит с основными законами Менделя о теории генетики, о специальном наследовании генов как животных, так и человека. Разбирается генетика пола. Так же разбирается наследование признаков, сцепленных с полом.
Министерство образования Рязанской области
Областное государственное бюджетное профессиональное
образовательное учреждение
«Касимовский нефтегазовый колледж»
Реферат
по дисциплине
«Биология»
Тема: «Основы генетики»
Выполнил:
___________
Проверил:
____________Касимов 2018 год.Содержание:
Введение
1. Гибридологический метод изучения наследования признаков Г.Менделя.
2. Законы Менделя.
3. Сцепленное наследование генов.
4. Взаимодействие генов.
5. Генетика пола. Наследование признаков, сцепленных с полом.
ЛитератураВведение.
Генетика
ДНК молекулярная основа наследственности. Каждая молекула ДНК является
двойной цепочкой из нуклеотидов (полинуклеотидов, соединяющихся посредине, образуя
двойную спираль).
Генетика (с стард.грец. genetikos порождать и genesisрождение) область
биологии; наука о генах, наследственности и вариативности организмов. Тот факт, что
живые существа наследуют черты своих родителей, был использован еще в доисторические
времена для улучшения производительности злаковых культур и животных путем
селективного разведения. Однако, современная генетика, которая стремиться понять
процесс наследования, реально началась только с работы Грегора Менделя в середине
девятнадцатого века. Несмотря на то, что он не знал физических основ наследственности,
Мендель заметил, что организмы наследуют черты через особые дискретные единицы
наследственности, которые сегодня мы называем генами.
Гены являются участками в ДНК молекула, состоящая из цепи четырех различных
типов нуклеотидов последовательность которых и есть генетической информацией,
которую наследуют организмы. В подавляющем большинстве случаев ДНК присутствуют в
форме двойных основ, с нуклеотидами на каждой основе, комплементарно дополняющих
друг друга. Каждая основа выступает в качестве шаблона для создания новой основы
партнера это физический метод копирования генов, которые могут быть унаследованы.
Последовательности нуклеотидов в генах транслируются клетками для выработки
цепи аминокислот и синтеза белков порядок аминокислот в белке соответствует порядку
нуклеотидов в гене. Эта связь между последовательностью нуклеотидов и
последовательность аминокислот называется генетическим кодом.Аминокислоты в белке
определяют, каким образом они составляют трехмерную форму, которая, в свою очередь,
отвечает за функцию белка. Белки выполняют практически все функции необходимые для
жизнедеятельности клеток. При изменение последовательности оснований в том участке
молекулы ДНК, которая несет генетическую информацию (гене) может изменить
последовательность аминокислот белка, изменив его форму и функции, что часто приводит
к негативному влиянию на функционирование клеток и организма в целом. Причем в
большинстве случаев происходит изменение только одного нуклеотида. Часто это
вызывает какоето определенное генетическое заболевание, которое может передаваться
по наследству.
Хотя генетические особенности играют чрезвычайно большую роль в появлении,
развитии, функционировании и поведении организмов, конечный результат (совокупностьпризнаков и особых свойств организма) является сочетанием генетических факторов с
условиями в которых происходит развитие организма. Например, наследственность играет
значительную роль в формировании такого признака, как рост человека, однако,
особенности питания и другие условия (например физическая активность, специфические
упражнения и т.д.), могут в значительной степени модулировать указанный признак в
зависимости от условий.1. Гибридологический метод изучения наследования
признаков Г.Менделя.
Генетика – наука о наследственности и изменчивости организмов. Развитие
генетики – характерная черта биологии ХХ в. Генетика изучает законы наследственности и
изменчивости, лежащие в основе эволюции органического мира и деятельности человека по
созданию новых сортов культурных растений и пород домашних животных, как это
установил еще Ч. Дарвин.
Вспомните, как Дарвин определял наследственность. Наследственность это
свойство организма передавать свои признаки и особенности развития следующим
поколениям. Благодаря наследственности все особи в пределах вида сходны между собой.
Наследственность позволяет животным, растениям и микроорганизмам сохранять из
поколения в поколение характерные черты вида, породы, сорта.
Наследование признаков осуществляется через размножение. При половом
размножении новые поколения возникают в результате оплодотворения. Материальные
основы наследственности заключены в половых клетках. При бесполом или вегетативном
размножении новое поколение развивается или из одноклеточных спор, или из
многоклеточных образований. И при этих формах размножения связь между поколениями
осуществляется через клетки,
наследственности.
в которых заключены материальные основы
Вспомните, как Дарвин определял изменчивость. Изменчивость – свойство
организмов приобретать новые признаки в процессе индивидуального развития. Благодаря
изменчивости особи в пределах вида различаются между собой.
Следовательно, наследственность и изменчивость – два противоположных, но
взаимосвязанных свойства организма.
однородность вида, а изменчивость, наоборот, делает вид неоднородным.
Благодаря наследственности сохраняется
Различия между особями одного вида могут зависеть от изменения материальных
основ наследственности организма. Изменчивость определяется и внешними условиями.
Каждому известно, что проявление свойств породы во многом зависит от условий
содержания и кормления.Изменчиво
сть одуванчика, выращенного из одного корня
На рисунке показан результат опыта, проведенного над одуванчиком, корень
одуванчика разрезали пополам. Одну половину его высадили на равнине в условиях
высокой влажности. Выросло растение с крупными листьями, длинными цветоносами.
Другую половину посадили в горах. Выросло маленькое растение с мелкими листьями, с
очень коротким цветоносом. А между тем наследственность у них одинаковая.
Совокупность генов, которую организм получает от родителей, составляет его
генотип. Совокупность внешних и внутренних признаков – это фенотип.
Из приведенного примера становится ясно, что фенотип развивается в результате
взаимодействия генотипа и условий внешней среды.
Гибридологический метод изучения наследственности. Основные закономерности
передачи признаков в ряду поколений при половом размножении были впервые
установлены чешским ученым Грегором Менделем и опубликованы в 1865 г. Его
исследования долгое время не были правильно оценены. Лишь в 1900 г. они были как бы
переоткрыты и подтверждены несколькими учеными и стали основой вновь возникшей
отрасли биологии генетики.
Мендель проводил опыты на горохе. У этого растения много разных сортов,
отличающихся друг от друга хорошо выраженными
наследственными признаками. Имеются, например, сорта
с белыми и пурпурными цветками, с высоким и низким
стеблем, с желтыми и зелеными семенами, с гладкими и
морщинистыми семенами и т. п. Каждая из указанных
особенностей наследуется в пределах данного сорта У
гороха обычно происходит самоопыление, хотя возможно
и перекрестное опыление.
Мендель
применил
гибридологический метод
исследования – скрещивание различающихся по
определенным признакам родительских форм – и проследил проявление изучаемыхпризнаков в ряду поколений. Мендель шел аналитическим путем: из большого
многообразия признаков растений он вычленял одну или несколько пар противоположных
друг другу признаков и прослеживал проявление их в ряду следующих друг за другом
поколений. Характерной чертой опытов Менделя был точный количественный учет
проявления изучаемых признаков у всех особей. Это позволило ему установить
Анализ
определенные количественные закономерности в наследственности.
закономерностей наследственности Мендель начал с моногибридного скрещивания –
скрещивания родительских форм, наследственно различающихся лишь по одной паре
признаков.
Единообразие первого поколения гибридов.
Гибриды – организмы, получающиеся в результате скрещивания особей,
различающихся наследственными зачатками. Условно принято обозначать родительское
поколение латинской буквой P (лат. parentale – родительский), первое поколение гибридов
– F1, второе F2 (лат. filiale – дочерний) и т. д.
Если скрестить растения гороха с желтыми и зелеными семенами, то у всех
полученных в результате этого скрещивания растений первого поколения гибридов семена
будут желтыми. Противоположный признак (зеленые семена) как бы исчезает. В этом
проявляется установленное Менделем правило единообразия первого поколения гибридов.
Признак желтой окраски семян как бы подавляет проявление противоположного признака
(зеленая окраска) и все семена у гибридов F1, оказываются желтыми (единообразными).
Явление преобладания признака получило название доминирования, а преобладающий
признак называют доминантным. В рассматриваемом примере желтая окраска семян
доминирует над зеленой. Противоположный, внешне исчезающий признак (зеленая окраска)
называют рецессивным.
Первый закон Менделя. В потомстве от первого поколения гибридов (во втором
поколении – F2) наблюдается расщепление: появляются растения с признаками обоих
родителей в определенных численных соотношениях. Желтых семян оказывается примерно
в три раза больше, чем зеленых. Соотношение семян гороха с доминантными и
рецессивными признаками близко к отношению 3:1. В опыте Менделя были получены
следующие количественные отношения: желтых – 6022, зеленых – 2001. Аналогичные
результаты дали опыты по изучению других пар признаков. Оказалось, пурпурная окраска
венчика цветка доминирует над белой и во втором поколении гибридов дает то же
расщепление 3:1; гладкая форма семян доминирует над морщинистой. Рецессивный
признак в первом поколении гибридов не выявляется. В этом проявляется первый закон
Менделя, получивший название закона расщепления: гибриды первого поколения F1 придальнейшем размножении расщепляются; в их потомстве F2 снова появляются особи с
рецессивными признаками, составляющие примерно четвертую часть от всего числа
потомков.
Как будут проявляться признаки в третьем, четвертом и последующих поколениях
гибридов? Для решения этого вопроса Мендель путем самоопыления получил потомство
третьего и последующих поколений.
Ход моногибридного скрещивания
Светлые кружки организмы с доминантным признаком, темные с рецессивным признаком
На рисунке видно, что растения, обладавшие рецессивным признаком, далее в
любом числе поколений не обнаруживали расщепления. В их потомстве никогда не
появлялось растений с доминантным признаком. Иначе себя вели гибриды второго
поколения, обладавшие доминантным признаком. Среди них при анализе потомства,
полученного путем самоопыления, обнаружили две группы. Первая, составляющая 1/3 от
общего числа растений с доминантным признаком, далее не расщепляется. В их потомстве,
в последующих поколениях, обнаруживается только доминантный признак. Иначе ведут
себя другие растения второго поколения, составляющие 2/3 от общего числа растений с
доминантным признаком. В их потомстве проявляется расщепление в том же соотношении
3:1 (3/4 доминантных, 1/4 рецессивных), как и у гибридов второго поколения. Изучение
последующих поколений дает сходный результат. Потомки растений с рецессивным
признаком не расщепляются.
У всех растительных и животных организмов при половом размножении происходит
расщепление в потомстве гибридов. Особи, сходные по внешним признакам, могут
обладать различными наследственными свойствами. Например, среди растений гороха с
желтыми семенами во втором поколении гибридов одни особи при самоопылении
обнаруживают в потомстве расщепление, другие же не расщепляются. Такие особи,
которые не обнаруживают в потомстве расщепления и сохраняют свои признаки в
«чистом» виде, называютгомозиготными по данному признаку (лат. «гомо» – равные,
одинаковые). Особей, которые в потомстве обнаруживают явление расщепления (т. е.являются по наследственным зачаткам гибридными), называют гетерозиготными по
данному признаку (лат. «гетеро» – разный).
Промежуточный характер наследования. В рассмотренных выше примерах
правило единообразия первого поколения гибридов выражалось в том, что все гибриды
внешне были похожи на одного из родителей, т. е. проявлялось доминирование. Это
наблюдается не всегда. Часто признаки у гетерозиготных форм носят промежуточный ха
актер, т. е. доминирование может быть неполным. На рисунке 103 представлены
результаты скрещивания двух наследственных форм растения ночная красавица. Одна из
них обладает красными цветками, другая – белыми. Все гибриды первого поколения имеют
розовые цветки, т. е. они носят промежуточный характер.
Моногибридное скрещивание ночной красавицы
При скрещивании гибридов между собой во втором поколении происходит
расщепление в отношении: один красный, два розовых и один белый (1:2:1). Очевидно, в
этом случае гетерозиготные (гибридные) растения внешне отличаются от гомозиготных
(красных и белых).
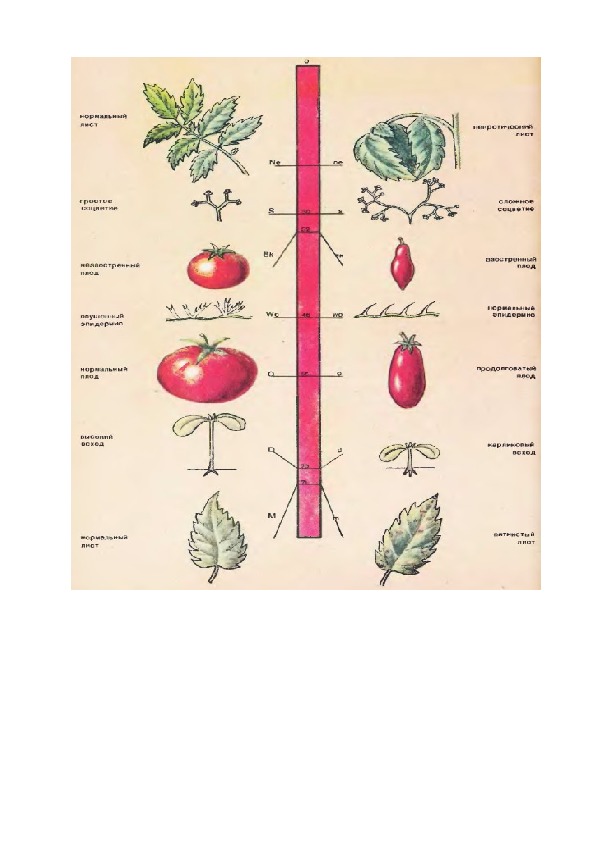
Генетическая карта фрагмента хромосомы томата (буквами обозначены названия генов, цифрами место их локализации)2. Законы Менделя
Моногибридное скрещивание. Первый закон Менделя.
В опытах Менделя при скрещивании сортов гороха, которые имели желтые и
зеленые семена, все потомство (т.е. гибриды первого поколения) оказалось с желтым
семенами. При этом не имело значения, из какого именно семена (желтого или зеленого)
выросли материнские (отцовские) растения. Итак, оба родителя в равной степени способны
передавать свои признаки потомству.
Аналогичные результаты были обнаружены и в опытах, в которых во внимание
брались другие признаки. Так, при скрещивании растений с гладкими и морщинистым
семенами все потомство имело гладкие семена. При скрещивании растений с пурпурными и
белыми цветками у всех гибридов оказались лишь пурпурные лепестки цветков и т. д.
Обнаруженная закономерность получила название первый закон Менделя, или закон
единообразия гибридов первого поколения. Состояние (аллель) признака, проявляющегося
в первом поколении, получило название доминантного, а состояние (аллель), которое в
первом поколении гибридов не проявляется, называется рецессивным. «Задатки»
признаков (по современной терминологии гены) Г. Мендель предложил обозначать
буквами латинского алфавита. Состояния, принадлежащие к одной паре признаков,
обозначают одной и той же буквой, но доминантный аллель большой, а рецессивный
маленькой.
Второй закон Менделя.
При скрещивании гетерозиготных гибридов первого поколения между собой
(самоопыления или родственное скрещивание) во втором поколении появляются особи как
с доминантными, так и с рецессивными состояниями признаков, т.е. возникает
расщепление, которое происходит в определенных отношениях. Так, в опытах Менделя на
929 растений второго поколения оказалось 705 с пурпурными цветками и 224 с белыми. В
опыте, в котором учитывался цвет семян, с 8023 семян гороха, полученных во втором
поколении, было 6022 желтых и 2001 зеленых, а с 7324 семян, в отношении которых
учитывалась форма семени, было получено 5474 гладких и 1850 морщинистых. Исходя из
полученных результатов, Мендель пришел к выводу, что во втором поколении 75% особей
имеют доминантное состояние признака, а 25% рецессивное (расщепление 3:1). Эта
закономерность получила название второго закона Менделя, или закона расщепления.
Согласно этому закону и используя современную терминологию, можно сделать
следующие выводы:
а) аллели гена, находясь в гетерозиготном состоянии, не изменяют структуру друг
друга;б) при созревании гамет у гибридов образуется примерно одинаковое число гамет с
доминантными и рецессивными аллелями;
в) при оплодотворении мужские и женские гаметы, несущие доминантные и
рецессивные аллели, свободно комбинируются.
При скрещивании двух гетерозигот (Аа), в каждой из которых образуется два типа
гамет (половина с доминантными аллелями А, половина с рецессивными а),
необходимо ожидать четыре возможных сочетания. Яйцеклетка с аллелью А может быть
оплодотворена с одинаковой долей вероятности как сперматозоидом с аллелью А, так и
сперматозоидом с аллелью а; и яйцеклетка с аллелью а сперматозоидом или с аллелью А,
или аллелью а. В резульатате получаются зиготы АА, Аа, Аа, аа или АА, 2Аа, аа.
По внешнему виду (фенотипу) особи АА и Аа не отличаются, поэтому расщепление
выходит в соотношении 3:1. По генотипу особи распределяются в соотношении
1АА:2Аа:аа. Понятно, что если от каждой группы особей второго поколения получать
потомство только самоопылением, то первая (АА) и последняя (аа) группы (они
гомозиготные) будут давать только однообразное потомство (без расщепления), а
гетерозиготные (Аа) формы будут давать расщепление в соотношении 3:1.
Таким образом, второй закон Менделя, или закон расщепления, формулируется
так: при скрещивании двух гибридов первого поколения, которые анализируются по одной
альтернативной паре состояний признака, в потомстве наблюдается расщепление по
фенотипу в соотношении 3:1 и по генотипу в соотношении 1:2:1.
Третий закон Менделя, или закон независимого наследования признаков.
Изучая расщепления при дигибридном скрещивании, Мендель обратил внимание на
следующее обстоятельство. При скрещивании растений с желтыми гладкими (ААВВ) и
зелеными морщинистыми (ааbb) семенами во втором поколении появлялись новые
комбинации признаков: желтые морщинистое (Ааbb) и зеленые гладкие (ааВb), которые не
встречались в исходных формах. Из этого наблюдения Мендель сделал вывод, что
расщепление по каждой признаку происходит независимо от второго признака. В этом
примере форма семян наследовалась независимо от их окраски. Эта закономерность
получила название третьего закона Менделя, или закона независимого распределения
генов.
Третий закон Менделя формулируется следующим образом: при скрещивании
гомозиготных особей, отличающихся по двум (или более) признаках, во втором поколении
наблюдаются независимое наследование и комбинирование состояний признаков, если
гены, которые их определяют, расположенные в разных парах хромосом. Это возможно
потому, что во время мейоза распределение (комбинирования) хромосом в половыхклетках при их созревании идет независимо и может привести к появлению потомства с
комбинацией признаков, отличных от родительских и прародительский особей.
Для записи скрещиваний нередко используют специальные решетки, которые
предложил английский генетик Пеннет (решетка Пеннета). Ими удобно пользоваться при
анализе полигибридних скрещиваний. Принцип построения решетки состоит в том, что
сверху по горизонтали записывают гаметы отцовской особи, слева по вертикали гаметы
материнской особи, в местах пересечения вероятные генотипы потомства.3. Сцепленное наследование генов
Хромосомная теория наследственности (Т. X. Морган и
др.)
Доказано, что количество наследственных признаков организма значительно
превышает число хромосом гаплоидного набора. Так, в гаплоидном наборе классического
объекта генетических исследований мухидрозофилы есть только четыре хромосомы, но
число наследственных признаков и, соответственно, генов которые их определяют,
несомненно, значительно больше. Это означает, что в каждой хромосоме находится много
генов. Поэтому вместе с признаками, которые наследуются независимо, должны
существовать и такие, которые наследуются сцеплено друг с другом, так как они
определяются генами, расположенными в одной хромосоме. Такие гены образуют группу
сцепления. Количество групп сцепления в организмах определенного вида равно
количеству хромосом в гаплоидном наборе (например, у дрозофилы 1пара = 4, у человека
1пара = 23).
Экспериментальные исследования явления сцепленного наследования провел
выдающийся американский генетик Т. X. Морган со своими сотрудниками. Их результаты
обосновали предложенную учёными хромосомную теорию наследственности.
Следует отметить, что Т. X. Морган, как в свое время Г. Мендель, удачно избрал
для исследований мухудрозофилу, которая впоследствии стала классическим объектом
для генетических экспериментов. Дрозофил легко содержать в лабораториях, у них
высокая плодовитость, быстрая смена поколений (при оптимальных условиях содержания
новое поколение возникает каждые полторыдве недели), небольшое число хромосом, что
упрощает наблюдения.
Явление сцепленного наследования Т. X. Морган установил в процессе следующего
опыта. Самцов дрозофилы, гомозиготных за доминантными аллелями окраски тела (серое)
и формой крыльев (нормальные), скрестили с самками, гомозиготными за
соответствующими рецессивными аллелями (черное тело недоразвитые крылья).
Генотипы этих особей обозначили соответственно ЕЕУУ и ееуу. Все гибриды первого
поколения имели серое тело и нормальные крыла, то есть были гетерозиготными по обеим
парами аллелей (генотип ЕЕУУ). Затем гибридов скрестили с особями, гомозиготными за
соответствующими рецессивными аллелями (анализирующее скрещивание).
Теоретически можно было ожидать два варианта расщепление. Если бы гены, что
обусловливают окрас тела и форму крыльев, содержались в негомологичных хромосомах,
то есть наследовались независимо, расщепление должно быть таким: 25% особей с серымтелом и нормальными крыльями, 25% с серым телом и недоразвитыми крыльями, еще
25% с черным телом и нормальными крыльями и 25% с черным телом и недоразвитыми
крыльями (то есть в соотношении 1:1:1:1). Если бы эти гены размещались в одной
хромосоме и наследовались сцеплено, то было бы получено 50% особей с серым телом и
нормальными крыльями и 50% с черным телом и недоразвитыми крыльями (т.е. 1:1).
На самом деле 41,5% особей имели серое тело и нормальные крылья, 41,5%
черное тело и недоразвитые крылья, 8,5% серое тело и недоразвитые крылья и 8,5%
черное тело и нормальные крылья, то есть расщепление приближалось к соотношению
фенотипов 1:1 (как в случае сцепленного наследования), но вместе с тем проявились все
четыре варианта фенотипа (как в случае независимого наследования). На основании этих
данных Т. X. Морган предположил, что гены, определяющие окраску тела и форму
крыльев, расположены в одной хромосоме, но в процессе мейоза при образовании гамет
гомологические хромосомы могут обмениваться участками, т.е. имеет место явление,
получившее название перекрёст хромосом, или кроссинговер .
Кроссинговер обмен участками гомологичных хромосом в процессе клеточного
деления, преимущественно в профазе первого мейотического разделения, иногда в митозе.
Опытами Т. Моргана, К. Бриджеса и А. Стертеванта было показано, что нет абсолютно
полного сцепления генов, при котором гены передавались бы всегда вместе. Вероятность
того, что два гена, локализованные в одной хромосоме, не разойдутся в процессе мейоза,
колеблется в пределах 10,5. В природе преобладает неполное сцепление, обусловленное
перекрёстком гомологичных хромосом и рекомбинацией генов. Цитологическая картина
кроссинговера была впервые описана датским ученым Ф. Янсенсом .
Кроссинговер проявляется только тогда, когда гены находятся в гетерозиготном
состоянии (АВ / ав). Если гены находятся в гомозиготном состоянии (АВ / АВ или аВ/аВ),
обмен идентичными участками не дает новых комбинаций генов в гаметах и в поколении.
Частота (процент) перекрёстка между генами зависит от расстояния между ними: чем
дальше они располагаются друг от друга, тем чаще происходит кроссинговер. Т. Морган
предложил расстояние между генами измерять кроссинговером в процентах, по формуле:
N1/N2 X 100 = % кроссинговера,
где N1 общее число особей в F;
N2 суммарное число кроссоверных особей.
Отрезок хромосомы, на котором осуществляется 1% кроссинговера, равна одной
морганиде (условная мера расстояния между генами). Частоту кроссинговера используют
для того, чтобы определить взаимное расположение генов и расстояние между ними. Для
построения генетической карты человека пользуются новыми технологиями, кроме того
построены цитогенетические карты хромосом.Различают несколько типов кроссинговера: двойной, множественный (сложный),
неправильный, неровный.
Кроссинговер приводит к новому сочетанию генов, вызывает изменение фенотипа.
Кроме того, он наряду с мутациями является важным фактором эволюции организмов.
Указанные доказательства были положены в основу хромосомной теории
наследственности:
1.
Гены размещаются в хромосомах по длине в линейном порядке;
различные хромосомы содержат неодинаковое число генов; набор генов каждой из
негомологичних хромосом уникальный.
2.
2. Аллельные гены занимают определенные и идентичные локусы
(места) гомологичных хромосом.
3.
3. Гены, расположенные в одной хромосоме, образуют группу
сцепления, благодаря чему имеет место сцепления некоторым признакам, которые
вместе (сцепленные) передаются потомкам. Количество групп сцепления равно
гаплоидному набору хромосом. Сцепление не абсолютное.
4.
4. Во время мейоза, которое происходит только при образовании
гамет, диплоидные число хромосом уменьшается вдвое (гаплоидные число). Это
соответствует закону расщепления, согласно которому генетический материал
обоих родителей поразному комбинируется в гаметах.
5.
5. Согласно закону независимого распределения родительские и
материнские наборы не сцепленных генов расщепляются независимо друг от друга.
Если не сцепленные гены расположены в разных хромосомах, то во время мейоза
материнские и отцовские хромосомы должны распределиться между гаметы
случайно.
6.
6. Между генами гомологичных родительских и материнских групп
реципрокные
сцепления могут происходить,
рекомбинации. Этому соответствует образование хиазм при конъюгации
благодаря кроссинговеру,
гомологичных хромосом в мейозе (генетический кроссинговер).
7.
7. Сила сцепления между генами обратно пропорциональна
расстоянию между ними. Чем ближе расположены гены в одной хромосоме, тем
сильнее их сцепление, тем меньше будет возникать рекомбинаций между ними, и
наоборот. Расстояние между генами измеряется в процентах кроссинговера. Один
процент
морганиди.
кроссинговера
одной
соответствует
8. Каждый биологический вид характеризуется специфическим набором хромосом
– кариотипом.4. Взаимодействие генов
Взаимодействие аллельных генов
В состав генотипа входит большое количество генов, функционирующих и
взаимодействуют как целостная система. Г. Мендель в своих опытах обнаружил только
одну форму взаимодействия между аллельными генами полное доминирование одной
аллели и полную рецесивнисть другой. Генотип организма нельзя рассматривать как
простую сумму независимых генов, каждый из которых функционирует вне связи с
другими. Фенотипное проявления того или иного признака являются результатом
взаимодействия многих генов.
Различают две основных группы взаимодействия генов: взаимодействие между
аллельными генами и взаимодействие между неаллельнимы генами. Однако следует
понимать, что это не физическое взаимодействие самих генов, а взаимодействие первичных
и вторичных продуктов, которые обусловят тот или иной признак. В цитоплазме
происходит взаимодействие между белками ферментами, синтез которых опрелятся
генами, или между веществами, которые образовываются под влиянием этих ферментов.
Возможны следующие типы взаимодействия:
1)
для образования определенного признака необходимо взаимодействие
двух ферментов, синтез которых опрелятся двумя неаллельнимы генами;
2)
фермент, что был синтезирован с участием одного гена, полностью
подавляет или инактивирует действие фермента, что был образован другим
неаллельным геном;
3)
два ферменты, образование которых контролируется двумя
неаллельми генами, влияющими на один признак или на один процесс так, что их
совместное действие приводит к возникновению и усилению проявления признака.
Взаимодействие аллельных генов.
Гены,
которые занимают идентичные
(гомологические) локусы в гомологичных хромосомах, называются аллельными. У каждого
организма есть по два аллельных гена.
Известны такие формы взаимодействия между аллельными генами: полное
доминирование, неполное доминирование, кодоминированием и сверхдоминирование.
Основная форма взаимодействия полное доминирование, которое впервые описано
Г. Менделем. Суть его заключается в том, что в гетерозиготном организме проявление
одной из аллелей доминирует над проявлением другой. При полном доминировании
расщепления по генотипу 1:2:1 не совпадает с расщеплением по фенотипу 3:1. В
медицинской практике с двух тысяч моногенных наследственных болезней почти вполовины имеет место доминированое проявления патологических генов над нормальными.
В гетерозигот патологический аллель проявляется в большинстве случаев признаками
заболевания (доминантный фенотип).
Неполное доминирование форма взаимодействия, при которой у гетерозиготного
организма (Аа) доминантный ген (А) не полностью подавляет рецессивный ген (а),
вследствие чего проявляется промежуточный между родительскими признак. Здесь
расщепление по генотипу и фенотипу совпадает и составляет 1:2:1
При кодоминировании в гетерозиготных организмах каждый из аллельных генов
вызывает формирование зависимого от него продукта, то есть оказываются продукты
обеих аллелей. Классическим примером такого проявления является система групп крови,
в частности система АBО, когда эритроциты человека несут на поверхности антигены,
контролируемые обеими аллелями. Такая форма проявления носит название
кодоминированием.
Сверхдоминирование когда доминантный ген в гетерозиготном состоянии
проявляется сильнее, чем в гомозиготном. Так, у дрозофилы при генотипе ААнормальная
продолжительность жизни; Аа удлиненная триватисть жизни; аа летальный исход.
Множественный алелизм
У каждого организма есть только по два аллельных гена. Вместе с тем нередко в
природе количество аллелей может быть более двух, если какой то локус может находится
в разных состояниях. В таких случаях говорят омножественные аллели или множественный
аллеломорфизм.
Множественные аллели обозначаются одной буквой с разными индексами,
например: А, А1, А3 ... Аллельные гена локализуются в одинаковых участках
гомологичных хромосом. Поскольку в кариотипе всегда присутствуют по две
гомологичных хромосомы, то и при множественных аллелях каждый организм может иметь
одновременно лишь по два одинаковых или различных аллели. В половую клетку (вместе с
различием гомологичних хромосом) попадает только по одному из них. Для
множественных аллелей характерное влияние всех аллелей на один и тот же признак.
Отличие между ними заключается лишь в степени развития признака.
Второй особенностью является то, что в соматических клетках или в клетках
диплоидных организмов содержится максимум по две аллели из нескольких, поскольку они
расположены в одном и том же локусе хромосомы.
Еще одна особенность присуща множественным аллелям. По характеру
доминирования аллеломорфные признаки размещаются в последовательном ряду: чаще
нормальный, неизмененный признак доминирует над другими, второй ген ряда рецессивныйотносительно первого, однако доминирует над следующими и т.д. Одним из примеров
проявления множественных аллелей у человека есть группы крови системы АВО.
Множественный алелизм имеет важное биологическое и практическое значение,
поскольку усиливает комбинативну изменчивость, особенно генотипического.
Взаимодействие неалельних генов
Известно много случаев, когда признак или свойства детерминируются двумя или
более неалельнимы генами, которые взаимодействуют между собой. Хотя и здесь
взаимодействие условно, потому что взаимодействуют не гены, а контролируемые ими
продукты. При этом имеет место отклонение от менделивских закономерностей
расщепления.
Различают четыре основных типа взаимодействия генов: комплементарность,
эпистаз, полимерию и модифицирующее действие (плейотропия).
Комплементарность это такой тип взаимодействия неаллельних генов, когда один
доминантный ген дополняет действие другого неаллельного доминантного гена, и они
вместе определяют новый признак, который отсутствует у родителей. Причем
соответственный признак развивается только в присутствии обоих неаллельних генов.
Например, сера окраска шерсти у мышей контролируется двумя генами (А и В). Ген А
детерминирует синтез пигмента, однако как гомозиготы (АА), так и гетерозиготы (Аа)
альбиносы. Другой ген В обеспечивает скопления пигмента преимущественно у основания
и на кончиках волос. Скрещивания дигетерозигот (АаВЬ х АаВЬ) приводит к расщеплению
гибридов в соотношении 9:3:4. Числовые соотношения при комплементарном
взаимодействии могут быть как 9:7; 9:6:1 (видоизменение менделивского расщепления).
Примером комплементарного взаимодействия генов у человека может быть синтез
защитного белка интерферона. Его образование в организме связано с комплементарным
взаимодействием двух неаллельних генов, расположенных в разных хромосомах.
Эпистаз это такое взаимодействие неаллельных генов, при котором один ген
подавляет действие другого неаллельного гена. Угнетение могут вызывать как
доминантные, так и рецессивные гены (А> В, а> В, В> А, В> А), и в зависимости от этого
розличают эпистаз доминантный и рецессивный. Подавляющий ген получил
названиеингибитора или супрессора. Геныингибиторы в основном не детерминируют
развитие определенного признака, а лишь подавляют действие другого гена.
Ген, эффект которого подавляется, получил название гипостатичного. При
епистатичном взаимодействияи генов расщепление по фенотипу в F2 составляет 13:3;
12:3:1 или 9:3:4 и др. Окрас плодов тыквы, масть лошадей определяются этим типом
взаимодействия.Если генсупрессор рецессивный, то возникает криптомерия (греч. хриштад
тайный, скрытый). У человека таким примером может быть "Бомбейский феномен". В этом
случае редкий рецессивный аллель "х" в гомозиготном состоянии (мм) подавляет
активность гена jB (определяющий В (III) группу крови системы АВО). Поэтому женщина с
генотипом jв_хх, фенотипно имеет I группу крови 0 (I).
Полигенное наследования количественных признаков
плейотропия
экспрессивность и пенетрантность генов
Большинство количественных признаков организмов определяется несколькими
неаллельнимы генами (полигенами). Взаимодействие таких генов в процессе формирования
признака называется полимерным. В этом случае две или более доминантных аллели в
равной степени влияют на развитие одной и того же признаки. Поэтому полимерные гены
принято обозначать одной буквой латинского алфавита с цифровым индексом, например:
А1А1 и а1а1. Впервые однозначные факторы были выявлены шведским генетиком
НильсонЭле (1908 г.) при изучении наследования цвета в пшеницы. Было установлено, что
этот признак зависит от двух полимерных генов, поэтому при скрещивании доминантних и
рецессивных дигомозигот окрашенной (А1А1, А2 А2) с бесцветной (а1а1, а2а2) в F, все
растения дают окрашенные семена, хотя они светлее, чем родительские экземпляры,
которые имеют красное семя. В F, при скрещивании особей первого поколения
проявляется расщепление по фенотипу в соотношении 15:1, потому бесцветным является
лишь рецессивные дигомозиготы (а1а1 а2а2). В пигментированных экземплярах
интенсивность цвета очень отличается в зависимости от количества полученных ими
доминантних аллелей: максимальная в доминантных дигомозигот (А1А1, А2 А2) и
минимальная у носителей одного из доминантных аллелей.
Важная особенность полимерии суммация действия неаллельних генов на развитие
количественных признаков. Если при моногенном наследовании признака возможны три
варианта "доз" гена в генотипе: АА, Аа, аа, то при полигенных количество их возрастает до
четырех и более. Суммация "доз" полимерных генов обесчивает существования
непрерывных рядов количественных изменений.
Биологическое значение полимерии заключается еще и в том, что признаки,
кодируемые этими генами, более стабильны, чем те, которые кодируются одним геном.
Организм без полимерных генов был бы очень неустойчивым: любая мутация или
рекомбинация приводила бы к резкой изменчивости, а это в большинстве случаев имеет
неблагоприятный характер.У животных и растений есть много полигенных признаков, среди них и ценные для
хозяйства: интенсивность роста, скороспелость, яйценоскость, количество молока,
содержание сахаристых веществ и витаминов и т.п.
Пигментация кожи у человека определяется пятью или шестью полимерными
генами. В коренных жителей Африки (негроидной расы) преобладают доминантные аллели,
у представителей европеоидной расы рецессивные. Поэтому мулаты имеют
промежуточную пигментацию, но при браках мулатов у них возможно появление как более,
так и менее интенсивно пигментированных детей.
Многие морфологические, физиологические и патологические особенности
человека определяются полимерными генами: рост, масса тела, величина артериального
давления и др. Развитие таких признаков у человека подчиняется общим законам
полигенного наследования и зависит от условий среды. В этих в случаях наблюдается,
например, склонность к гипертонической болезни, ожирению и др. Данные признаки при
благоприятных условиях среды могут не проявиться или проявиться незначительно. Эти
полигенные признаки отличаются от моногенных. Изменяя условия среды можно
обеспечить профилактику ряда полигенных заболеваний.
Плейотропия
Плейотропное действие генов это зависимость нескольких признаков от одного
гена, то есть множественное действие одного гена. В дрозофилы ген белого цвета глаз
одновременно влияет на цвет тела, длины, крыльев, строение полового аппарата, снижает
плодовитость, уменьшает продолжительность жизни. У человека известна наследственная
болезнь арахнодактилия ("паучьи пальцы"очень тонкие и длинные пальцы), или болезнь
Марфана. Ген, отвечающий за эту болезнь, вызывает нарушение развития соединительной
ткани и одновременно влияет на развитие нескольких признаков: нарушение строения
хрусталика глаза, аномалии в сердечнососудистой системе.
Плейотропное действие гена может быть первичным и вторичным. При первичной
плейотропии ген проявляет свой множественный эффект. Например, при болезни Хартнупа
мутация гена приводит к нарушению всасывания аминокислоты триптофана в кишечнике и
его реабсорбции в почечных канальцах. При этом поражаются одновременно мембраны
эпителиальных клеток кишечника и почечных канальцев с расстройствами
пищеварительной и выделительной систем.
При вторичной плейотропии есть один первичный фенотипний проявление гена,
вслед за которым развивается ступенчатый процесс вторичных изменений, приводящих к
множественным эффектам. Так, при серповидно клеточной анемии у гомозигот
наблюдается несколько патологических признаков: анемия, увеличенная селезенка,поражение кожи, сердца, почек и мозга. Поэтому гомозиготы с геном серповидно
клеточной анемии гибнут, как правило, в детском возрасте. Все эти фенотипные
проявления гена составляют иерархию вторичных проявлений. Первопричиной,
непосредственным фенотипним проявлением дефектного гена является аномальный
гемоглобин и эритроциты серповидной формы. Вследствие этого происходят
последовательно другие патологические процессы: слипание и разрушение эритроцитов,
анемия, дефекты в почках, сердце, мозге эти патологические признаки вторичны.
При плейотропии, ген, воздействуя на какой то один основнй признак, может также
менять, модифицировать проявление других генов, в связи с чем введено понятие о генах
модификаторах. Последние усиливают или ослабляют развитие признаков, кодируемых
"основным" геном.
Показателями зависимости функционирования наследственных задатков от
характеристик генотипа является пенетрантность и экспрессивность.
Рассматривая действие генов, их аллелей необходимо учитывать и
модифицирующее влияние среды, в которой розвивается организм. Если растения примулы
скрещивать при температуре 1520 ° С, то в F1 согласно менделивской схеме, все
поколения будут иметь розовые цветы. Но когда такое скрещивание проводить при
температуре 35 °С, то все гибриды будут иметь цветы белого цвета. Если же осуществлять
скрещивания при температуре около 30 ° С, то возникает разное соотношение (от 3:1 до
100%) растений с белыми цветами.
Такое колебание классов при расщеплении в зависимости от условий среды
получило название пенетрантность сила фенотипного проявления. Итак, пенетрантность
это частота проявления гена, явление появления или отсутствия признака у организмов,
одинаковых по генотипу.
Пенетрантность значительно колеблется как среди доминантных, так и среди
рецессивных генов. Наряду с генами, фенотип которых появляется только при сочетании
определенных условий и достаточно редких внешних условий (высокая пенетрантность), у
человека есть гены, фенотипное проявление которых происходит при любых соединениях
внешних условий (низкая пенетрантность). Пенетрантностью измеряется процентом
организмов с фенотипным признаком от общего количества обследованных носителей
соответствующих аллелей.
Если ген полностью, независимо от окружающей среды, определяет фенотипное
проявление, то он имеет пенетрантность 100 процентов. Однако некоторые доминантные
гены проявляются менее регулярно. Так, полидактилия имеет четкое вертикальное
наследования, но бывают пропуски поколений. Доминантная аномалия преждевременноеполовое созревание присуще только мужчинам, однако иногда может передаться
заболевания от человека, который не страдал этой патологией. Пенетрантностью
указывает, в каком проценте носителей гена оказывается соответствующий фенотип. Итак,
пенетрантность зависит от генов, от среды, от того и другого. Таким образом, это не
константное свойство гена, а функция генов в конкретных условиях среды.
Экспрессивность (лат. ехргеssio выражение) это изменение количественного
проявления признака в разных особейносителей соответствующего аллелей.
При доминантных наследственных заболеваниях экспрессивность может колебаться.
В одной и той же рсемье могут проявляться наследственные болезни от легких, едва
заметных до тяжелых: различные формы гипертонии, шизофрении, сахарного диабета и т.д.
Рецессивные наследственные заболевания в пределах семьи проявляются однотипно и
имеют незначительные колебанийния экспрессивности.5. Генетика пола.
Пол
совокупность признаков, по которым производится специфическое
разделение особей или клеток, основанное на морфологических и физиологических
особенностях,
комбинирование в потомках наследственных задатков родителей.
позволяющее осуществлять в процессе полового размножения
Морфологические и физиологические признаки, по которым производится
специфическое разделение особей, называется половым.
Признаки, связанные с формированием и функционированием половых клеток,
называется первичными половыми признаками. Это гонады (яичники или семенники), их
выводные протоки, добавочные железы полового аппарата, копулятивные органы. Все
другие признаки, по которым один пол отличается од другого, получили название
вторичных половых признаков. К ним относят: характер волосяного покрова, наличие и
развитие молочных желез, строение скелета, тип развития подкожной жировой клетчатки,
строение трубчатых костей и др.
Наследование признаков, сцепленных с полом.
Морган и его сотрудники заметили, что наследование окраски глаз у дрозофилы
зависит от пола родительских особей, несущих альтернативные аллели. Красная окраска
глаз доминирует над белой. При скрещивании красноглазого самца с белоглазой самкой в
F1, получали равное число красноглазых самок и белоглазых самцов. Однако при
скрещивании белоглазого самца с красноглазой самкой в F1 были получены в равном числе
красноглазые самцы и самки . При скрещивании этих мух F1, между собой были получены
красноглазые самки, красноглазые и белоглазые самцы, но не было ни одной белоглазой
самки. Тот факт, что у самцов частота проявления рецессивного признака была выше, чем
у самок, наводил на мысль, что рецессивный аллель, определяющий белоглазость,
находится в Х хромосоме, а Y хромосома лишена гена окраски глаз. Чтобы проверить
эту гипотезу, Морган скрестил исходного белоглазого самца с красноглазой самкой из F1.
В потомстве были получены красноглазые и белоглазые самцы и самки. Из этого Морган
справедливо заключил, что только Х хромосома несет ген окраски глаз. В Y – хромосоме
соответствующего локуса вообще нет. Это явление известно под названием наследования,
сцепленного с полом.
Гены, находящиеся в половых хромосомах, называют сцепленными с полом. В Х
хромосоме имеется участок, для которого в Yхромосоме нет гомолога. Поэтому у особей
мужского пола признаки, определяемые генами этого участка, проявляются даже в том
случае, если они рецессивны. Эта особая форма сцепления позволяет объяснить
наследование признаков, сцепленных с полом.При локализации признаков как в аутосоме, так и в Х b Yхромосоме наблюдается
полное сцепление с полом. У человека около 60 генов наследуются в связи с Х
хромосомой, в том числе гемофелия, дальтонизм (цветовая слепота), мускульная
дистрофия, потемнение эмали зубов, одна из форм агаммглобулинемии и другие.
Наследование таких признаков отклоняется от закономерностей, установленных
Г.Менделем. Ххромосома закономерно переходит от одного пола к другому, при этом
дочь наследует Ххромосому отца, а сын Ххромосому матери. Наследование, при котором
сыновья наследуют признак матери, а дочери признак отца получило, название крисс
кросс (или крестнакрест).
Известны нарушения цветового зрения, так называемая цветовая слепота. В основе
появления этих дефектов зрения лежит действие ряда генов. Краснозеленая слепота
обычно называется дальтонизмом. Еще задолго до появления генетики в конце XVIII и в
XIX в. Было установлено, что цветовая слепота наследуется согласно вполне
закономерным правилам. Так, если женщина, страдающая цветовой слепотой, выходит
замуж за мужчину с нормальным зрением, то у их детей наблюдается очень своеобразная
картина перекрестного наследования. Все дочери от такого брака получат признак отца,
т.е. они имеют нормальное зрение, а все сыновья, получая признак матери, страдают
цветовой слепотой (адальтонизм, сцепленный с Ххромосомой)
Р Ха Ха х Ха y
Ха Ха,y
F1 Ха Ха, Хаy
В том же случае, когда наоборот, отец является дальтоником, а мать имеет
нормальное зрение, все дети оказываются нормальными. В отдельных браках, где мать и
отец обладают нормальным зрением, половина сыновей может оказаться пораженными
цветовой слепотой. В основном наличие цветовой слепоты чаще встречается у мужчин.
Э.Вильсон объяснил наследование этого признака, предположив, что он локализовал в Х
хромосоме и что у человека гетерогаметным (XY) является мужской пол. Становится
вполне понятным, что в браке гомозиготной нормальной женщины (Ха Ха) с мужчиной
дальтоником (Х аy) все дети рождаются нормальными. Однако при этом, все дочери
становятся скрытыми носителями дальтонизма, что может проявиться в последующих
поколениях.
Другим примером наследования сцепленного с полом, может послужить
рецессивный полулетальный ген, вызывающий несвертываемость крови на воздухе
гемофилию. Это заболевание появляется почти исключительно только у мальчиков.
При гемофилии нарушается образование фактора VIII, ускоряющего свертывание
крови. ген, детерминирующий синтех фактора VIII, находится в участке Ххромосомы,недоминантным нормальным и рецессивным мутантным. Возможны следующие генотипы и
фенотипы:
Генотипы
Фенотипы
Хн Хн
Хн Хn
Хнy
Хny
Нормальная женщина
Нормальная женщина (носитель)
Нормальный мужчина
Мужчина гемофилик
В гомозиготном состоянии у женщин ген гемофилии летален.
Особей женского пола, гетерозиготных по любому из сцепленных с полом
называют носителями соответствующего рецессивного гена.
признаков,
Они
фенотипически нормальны, но половина их гамет несет рецессивный ген. Несмотря на
наличие у отца нормального гена, сыновья матерейносителей с вероятностью 50% будут
страдать гемофилией.
Один из наиболее хорошо документированных примеров наследования гемофилии
мы находим в родословной потомков английской королевы Виктории. Предполагают, что
ген гемофилии возник в результате мутации у самой королевы Виктории или у одного из ее
родителей. Среди унаследовавших это врожденное заболевание цесаревич Алексей, сын
последнего русского царя Николая II. Мать цесаревича, царица Александра Федоровна
(Алиса, рис.2), получила от своей бабушки королевы Виктории ген гемофилии и передала
его в четвертом поколении бывшему наследнику царского престола. На рис.2 показано, как
этот ген передавался ее потомкам.
Один из сцепленных с полом рецессивных генов вызывает особый тип мышечной
дистрофии (тип Дюмена). Эта дистрофия проявляется в раннем детстве и постепенно
ведет к инвалидности и смерти ранее 20летнего возраста. Потому мужчины с дистрофией
Дюмена не имеют потомства, а женщины гетерозиготные по гену этого заболевания, вполне
нормальны.Среди доминантных признаков, связанных с Х
хромосомой, можно указать на ген, который вызывает
недостаточность органического фосфора в крови. В
результате, при наличии этого гена, часто развивается
рахит, устойчивый к лечению обычными дозами
витамина А. В этом случае картина сцепленного с
полом наследования заметно отличается от того хода
передачи по поколениям, который был описан для
рецессивных болезней. В браках девяти больных
женщин со здоровыми мужчинами среди детей была
половина больных девочек и половина мальчиков.
Здесь, в соответствии с характером наследование
доминантного гена, в Ххромосомах произошло
расщепление в отношении 1:1:1:1.
Другим примером доминантного
гена,
локализованного в Ххромосоме человека, может
послужить ген, вызывающий дефект зубов, приводящий
к потемнению эмали зубов.
Так как гетерогаметный пол гемизиготен по
сцепленным с полом генам, то эти гены всегда
проявляются в их фенотипе, даже если они рецессивны.
Большинство генов, имеющихся в Ххромосоме, в Y
хромосоме отсутствует,
однако определенную
генетическую информацию она всетаки несет.
Различают два типа такой информации: вопервых, содержащуюся в генах,
присутствующих только в Yхромосоме, и, вовторых, в генах, присутствующих как в Y,
так и в Ххромосоме (гемфрагический диатез).
Yхромосома передается от отца всем его сыновьям, и только им. Следовательно,
для генов, содержащихся только в Yхромосоме, характерно голандрическое наследование,
т.е они передаются от отца к сыну и проявляются у мужского пола.
У человека в Yхромосоме содержатся по крайней мере три гена, один из которых
необходим для дифференциации семенников, второй требуется для проявления антигена
гистосовместимости, а третий оказывает влияние на размер зубов. Yхромосома имеет
немного признаков, среди которых есть патологические. Патологические признакинаследуются по параллельной схеме наследования (100%ое проявление по мужской
линии). К ним относят:
1) облысение;
2) гипертрихоз (оволосенение козелка ушной раковины в зрелом возрасте);
3) наличие перепонок на нижних конечностях;
4) ихтиоз (чешуйчатость и пятнистое утолщение кожи).Литература:
1. Общая биология 1011
Интернет ресурсы:
1. http://vseprogeny.ru/ru_osnovygenetyky.html
2. http://sbio.info/list.php?c=obbosnovgen
3. https://ru.wikipedia.org/wiki