Работа представлена на страницах машинописного текста. Содержит таблиц. Список литературы включает наименований на русском и на иностранных языках.
Ключевые слова: ОГУРЕЦ, ГЕНЕТИЧЕСКИЙ АНАЛИЗ, ТИП ЦВЕТКА, ПРИЗНАКИ КАЧЕСТВА ПЛОДА.
Объект исследования – огурец (Cucumis sativus L.)
Цель исследования: проведение генетического анализа выбранных признаков, результаты которого открывают возможность планомерного конструирования будущего сорта на основе объединения желательных признаков.
Федеральное агентство по образованию
Государственное образовательное учреждение
Высшего профессионального образования
«КУБАНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ»
Кафедра генетики, микробиологии и биотехнологии
Допустить дипломника к защите в ГАК
«___»_______________________2011 г.
Научный руководитель – др биол. наук,
проф. Ю. А. Волчков________________
ДИПЛОМНАЯ РАБОТА
ГЕНЕТИЧЕСКИЙ КОНТРОЛЬ ТИПА ЦВЕТКА И ПРИЗНАКОВ
КАЧЕСТВА ПЛОДА ОГУРЦА
Работу выполнила_______________________________________Борисова О. А.
Факультет биологический
Специальность 050102 Биология
Нормоконтролер, канд. биол. наук, доц.______________________Г. Г. Вяткина
Краснодар 2011
РЕФЕРАТ
Работа представлена на страницах машинописного текста. Содержит
таблиц. Список литературы включает наименований на русском и на
иностранных языках.
Ключевые слова: ОГУРЕЦ, ГЕНЕТИЧЕСКИЙ АНАЛИЗ, ТИП ЦВЕТКА,
ПРИЗНАКИ КАЧЕСТВА ПЛОДА.
Объект исследования – огурец (Cucumis sativus L.)
1Цель исследования: проведение генетического анализа выбранных признаков,
результаты которого открывают возможность планомерного конструирования
будущего сорта на основе объединения желательных признаков.
Материал: родительские формы, альтернативные по изучаемым признакам
и специально укомплектованные по итогам объединения семей с однотипным
расщеплением, значительные по объему (~300 растений) выборки гибридов.
Методы исследования: учет признаков визуальный или
органолептический; статистическая обработка данных с помощью метода хи
квадрат. Доказана моногенность по четырем признакам из пяти изученных и
дигенное взаимодействие по типу рецессивного эпистаза для признака тип
опушения завязи.
Область применения результатов – генетика и селекция огурца.
СОДЕРЖАНИЕ
Введение …………………………………………………………………………….4
1 Обзор литературы……………………………………………………………….6
1.1
1.2
2 Материал и методы исследования……………………………………………
3 Генетический контроль тип цветка и признаков качества плода огурца
3.1 Сравнительностатистический анализ отношения расщепления в отдельных
опытах по гибридизации с целью формирования репрезентативного материала
для генетического анализа
3.2 Итоги генетического анализа расщепления F2 гибридов от скрещивания
фенотипически различающихся форм огурца
2Заключение………………………………………………………………….
Выводы………………………………………………………………………
Библиографический список………………………………………………..
ВВЕДЕНИЕ
Цель селекции новых сортов и гибридов огурца состоит в повышении
содержания питательных веществ, витаминов, улучшении консистенции
мякоти, товарного вида, лежкости и технологических качеств плодов.
Новые сорта и гибриды должны превосходить существующие по
урожайности, скороспелости, иметь высокие вкусовые качества, долгое время
сохранять зеленую окраску плода и отличаться комплексной устойчивостью ко
всем наиболее вредоносным болезням, распространенным в данной зоне. Для
этого и проводится сложная работа по объединению многих хозяйственно и
биологически ценных признаков в одном генотипе. Поэтому селекционер,
кроме биологии растения и технологии выращивания, должен знать
закономерности наследования и изменчивости признаков, методы подбора и
получения исходного материала, системы скрещиваний, способствующие
усилению ценных признаков у будущих сортов, приемы отбора по отдельным
биологическим и хозяйственно полезным признакам, теорию иммунитета и
методики заражения и оценки растений огурца на устойчивость к болезням и
вредителям. [Юрина; Корганова,1985]
Одним из наиболее важных направлений селекции огурца на Крымской
ОСС ВИР, где выполнена экспериментальная часть моей работы, является
3получение сортов с обоеполым типом цветка (так называемых гермафродитных
форм), обладающих одновременно некоторыми желательными признаками
плода; к ним относятся матовая кожица плода, белошипость, зеленая окраска
зеленца, простое опушение, отсутствие горечи.
Генетический контроль этих признаков отчасти уже описан в литературе,
однако его необходимо повторить и на гибридном материале, с которым
работает станция, с целью убедиться, что разнообразие гибридов определяется
теми же, отчасти уже известными, генами. Генетический анализ перечисленных
признаков и входил в задачу работы.
С учетом того, что генетический анализ проводился не на данных
специально спланированных генетических опытов, а на данных, полученных
селекционерами, первую и наиболее трудоемкую, как выяснилось, задачу
работы составило комплектование достаточных по объему выборок гибридных
растений на основе объединения результатов опыта
1 Обзор литературы
1.1 Происхождение огурца
Систематическое положение
Царство:
Отдел:
Класс:
Порядок: Тыквоцветные
Растения
Покрытосеменные
Двудольные
4Семейство: Тыквенные
Род: Огурец
Вид: Огурец обыкновенный
Латинское название
Cucumis sativus L.
Огурцы — самая древняя овощная культура. Их возделывают уже около 6
тысяч лет. Родиной огурцов считается Индия. Здесь и сейчас можно встретить
дикие огурцы. Они, как лианы, обвивают деревья в лесах. Огурцами оплетены
заборы индийских селений.
Древним египтянам и грекам огурцы были известны как культурное
растение. В Древней Греции медики приписывали огурцам жаропонижающее
действие. У римских императоров Августа и Тиберия ни один обед не
обходился без огурцов.
В I веке нашей эры огурцы начали выращивать в Китае и Японии. Местные
земледельцы до сегодняшнего дня умело выращивают огурцы. Сначала в ящиках
на крышах подрастает рассада, а затем ее густо высаживают на хорошо
унавоженных маленьких огородах и подвязывают к кольцам. Цепляясь усиками
за подпорки, растения поднимаются вверх. И со шпалер свисают огурцы длиной
1,5 метра. Этот сорт огурцов выращивается и у нас в теплицах, но в отличие от
других видов растений, огурцы тем ценнее, чем они меньше.
[ http://www.pokupaem.com/2007/08/18/ogurec_istorija_ogurca.html]
Огурцы с давних пор являются любимым овощным растением и в России.
История говорит о том, что при царе Алексее Михайловиче под Москвой для
дворцовых нужд на огородах ежегодно выращивали десятки тысяч огурцов.
Согласно этимологическому словарю Фасмера, название заимствовано из
(незрелый). Этот овощ,
(огурец), которое восходит к
ἄωρος
ср.греч.
ἄγουρος
поедаемый в незрелом виде, нарочито противопоставляется дыне —
которую едят в зрелом виде.
πέπων
,
5Название этого растения на санскрите созвучно с именем легендарного
индийского князя, который имел шестьдесят тысяч детей, и связано с
многосемянностью плода. [http://ru.wikipedia.org/wiki/Cucumis_sativus]
В 1885 г. А. Декандоль высказал предположение, что диким родичем и
предком огурца Хардвиком в Гималаях вид Cucumis hardwickii Royle,
описанный в 1839 г. Гипотеза Декандоля ожидала подтверждения более 100 лет.
Обычно считается, что культура огурца возникла в глубокой древности. Однако
все древние описания этого растения указывают не на огурец, а на огуречную
дыню – аджур. Достоверные же известия о культуре огурца относятся к средним
векам.
Исторические, лингвистические, археологические и генетические методы
не дали ответа на вопрос о происхождении огурца. Наиболее логично
предположить, что эволюция этого вида, в том числе и изменение его
хромосом, происходило в крайних условиях существования.
В Индии была найдена дикая C. hardwickii Royle форма огурца, которая
не совпадала с морфологическим описанием, но имела 14 хромосом и мелкие
горькие плоды. Эту форму объединили с огурцом Хардвика в подвид под
общим таксономическим названием Ssp. hardwickii (Royle) Fil. Таким образом,
наличие дикого родича и происхождение огурца из Индии было окончательно
подтверждено.
Дикая форма огурца была исходной при его окультивировании. Кроме
дикой и вполне культурных форм сохранились и промежуточные
полукультурные формы с негорькими плодами, имеющими ничтожно полезную
массу. [Филов ,1961].
1.2 Биологические особенности
Огурец (Cucumis sativus, сем.Сucurbitaceae) однолетнее травянистое
растение семейства тыквенных.
Растения огурца в молодом возрасте (до цветения) прямостоячие. По мере
разрастания плети у большинства сортов приобретают стелющуюся форму.
6Листья на стеблях расположены поочередно. Корни различной длины от 30 до
100 см.
Огурцы перекрестно опыляющиеся растения. Однако у некоторых
тепличных длинноплодных сортов плоды могут образовываться и без опыления
цветков (партенокарпия), что представляет большой интерес для
селекционеров.
По особенностям цветения сорта и формы огурцов очень разнообразны.
Плод огурцов ложная ягода, имеющая от 3 до 5 семенных камер.
[Мещеров, Босс,1967] . Ценятся огурцы в основном за их вкусовые качества.
Питательными веществами плоды огурцов бедны. В них содержится 9495
% воды и всего 12% сахаров. Низка и общая калорийность огурцов: 1 кг. их
содержит лишь 3% калорий, необходимых человеку в течение суток.
Содержание витаминов А и С невысокое. Но в них относительно большое
содержание минеральных соединений щелочного характера, нейтрализующие
кислые соединения. [Ткаченко,1963]. Такая нейтрализация необходима для
полного усвоения белков.
Основными отличительными признаками сортов огурцов являются
размеры, форма, окраска поверхности и опушение плода.
Фазы зеленца плоды достигают через 812 дней после оплодотворения
завязи. Скорость прохождения растениями огурцов отдельных фаз развития
находится в большой зависимости от внешних условий. Первыми, как правило,
зацветают мужские цветки, но бывает одновременное зацветание мужских и
женских цветков. [Мещеров, Босс, 1967] .
1.3 Наследование качественных признаков у огурца
Для ускоренной селекционной работы важное значение имеет изучение
изменчивости и наследования основных хозяйственно ценных признаков. В
настоящее время в связи с развитием новых направлений в селекции
необходимы глубокие знания генетических свойств каждого селектируемого
признака. Вопросы эти, однако, разработаны еще крайне недостаточно.
Имеющиеся в литературе данные по наследованию у огурца касаются главным
7образом морфологических признаков, которые наиболее легко прослеживаются
в потомстве и наследуются сравнительно просто. Изучением наследования
признаков у огурца занимались R. Wellington (1913), W. J. Strong (1931), Н.Н.
Ткаченко (1935),A. E. Hutchins ( 1935, 1940), C. F. Poole (1944), W.S. Barham
(1953) .O Shifriss (1961), E. Kooistra (1968, 1969), B. Borghi (1972), R. B.
Robinson (1973), P. B. Whitaker (1974), Т. Р. Стрельникова (1977) и др.
В литературе по вопросу наследования признаков у огурца встречаются
всевозможные символы для разных генов. R. B. Robinson , P. B. Whitaker, V. M.
Munger (1976) внесли некоторые поправки в генетическую символику данной
культуры, предложили по некоторым генам новые символы или ввели символы
первых авторов, изучавших наследование соответствующих признаков.
Таблица Наследование признака у огурца
Символ
гена
Признак
Плод
Источник
Bt
bi
Горечь в плодах(bitter fruit)
W.S. Barham,1953
Отсутствие горечи в плодах и всех
J. M. Andeweg, J. M. De
частях растений( bitter free)
D(d)
Матовая поверхность плода(dull skin of
Brugh , 1959
W. J. Strong,1931
W
а
F(Acr,
AcrF,
D, st)
fruit)
Белая окраска зеленца рецессивна по
отношению к зеленой (white immature
fruit colour) рецессивна по отношению
к зеленой.
Половые типы
Андрокист (androecious)образует
первоначально тычиночные цветки,если
рецессивен по F
F. D. Cochran ,1937
Kubicki, 1969
Женский (Female) высокая, степень
Н. Н. Ткаченко, 1935
проявления женского пола;
Взаимодействует с а и m ;
Сильно модифицируется условиями
среды
gy, g
Женский(gynoecious) рецессивный ген
Н. Н. Ткаченко, 1935
8высокой степени проявления женского
Kubicki, 1974
пола
InF
Усилитель женского пола у F растений
Kubicki,1969
m(a,g,mo
)
(Intensifer of female sex expression)
Андромонокийст (andromonoecious)
Растения андромононоцийные, если
m+, моноцийные++, гиноцийные +F,
гермафрофитныеmF
J. T. Rosa,1928
m2(h)
Андромонойкист2, (andromonoecious)
Kubicki,1974
Tr
обоеполые цветки с нормальными
завязями
Тримонойкист образует мужские.
Обоеполые и женские цветки в
онтогенезе последовательно
Kubicki,1969
Эти сведения касаются в основном признаков, контролируемых
преимущественно одним геном; двумятремя пол и признаки устойчивости к
некоторым болезням и др. [Стрельникова, Маштакова,1984]
1.4 Пол у огурца и его значение в селекции
В растительном мире у всех известных высших цветковых растений
женский пол рецессивен, мужской доминантен. У огурцов наоборот: мужской
пол явно рецессивен, женский доминантен.
Изучение пола у огурцов, кроме теоретического, имеет большое
практическое значение, так как со степенью его проявления тесно связана
продуктивность растений, а также возможность производства гибридных семян.
Еще Корренсом Т.[Corrense, 1932] была предложена следующая схема
проявления пола у огурца:
Схема Корренса:
Исходная форма – гермафродитная.
Промежуточные формы андромоноция, тримоноция, гиномоноция. Конечные
формы андроция, моноция, гиноция.
Данная схема показывает эволюцию проявления пола у огурца.
1. Все цветки имеют функциональные тычинки и пестики.
2. Часть цветков гермафродитные, часть тычиночные.
3. Часть цветков тычиночные, часть пестичные, часть гермафродитные.
94. Часть цветков пестичные, часть гермафродитные.
5. Все цветки тычиночные.
6. Часть цветков тычиночные, часть пестичные.
7. Все цветки пестичные.
Основываясь на работах обобщающего характера[Стрельникова,1977],
таких как Пангало К.И. (1943), в которой дается первая сводка сведений о
половых типах у тыквенных; наиболее фундаментальных работах о половых
типах у огурца Мещерова Э. Т. (1986) и Кубицкого Б. (1969), можно считать
достаточно твердо установленными следующие основные половые типы у
растений огурца:
Обычный однодомный раздельнополый;
Феминизированный однодомный (псевдоженский);
Женский из частично двудомных форм;
Обоеполо мужской (андромоноция);
Обоеполый (гермафродитный).
Обычные однодомные раздельнополые растения образуют
преимущественно мужские (тычиночные) цветки и в значительно меньшем
количестве женские пестичные. В каждом узле располагаются цветки одного
пола мужские или женские.
Псевдоженский тип образует больше женских узлов, которые в
большинстве преобладают над мужскими. Недостатком этого полового типа
является не только его наследственная неустойчивость, но и то, что мужские
образуются на верхних ярусах растения.
Характерной особенностью частичнодвудомных форм является
преобладание в их составе растений женского типа. Они или совсем лишены
мужских цветков или образуют их в небольшом количестве узлов и только в
нижней части главного стебля. Характерной особенностью частично
двудомных форм является то, что женский тип растений у них доминантен.
Другая, меньшая часть растений частично двудомного сорта отнесена к
мужскому типу.
10Растения промежуточного типа, содержащиеся в небольшом количестве
(1020 %) в материнских формах, выведенных в результате гибридизации
частично двудомных сортов с обычными однодомными раздельнополыми
сортами, образуют преимущественно женские цветки, но отличаются от
растений женского типа большим количеством мужских цветков, размещенных
в разных ярусах главного стебля и боковых побегов.
На растениях типа андромоноция образуются обоеполые и мужские
цветки, причем последние количественно преобладают.
Картина наследования половых особенностей осложняется действием генов
модификаторов. Вопросам наследования пола сейчас уделяется большое
внимание. Ткаченко Н. Н. [Ткаченко, 1935] отметил, что различие между
женским и мужским типом растений зависит от одной пары факторов женский
тип F доминирует над мужским f. Было также установлено , что обоеполый тип
рецессивен по отношению к обычным формам. Позже Ткаченко Н. Н. высказал
предположение , что наследование признаков пола у огурца идет более сложным
путём.
Шифрис О. [Jhifriss ,1961] также отметил доминирование женского. Он
ввел символ Acr (усилитель женского пола). Ген «g» контролирует образование
пестичных цветков. Шифрис высказал предположение, что гены «Acr» « G» в
различном сочетании рецессивных и доминантных аллелей обуславливают
проявление основных половых типов. Им было выделено четыре половых типа:
1. Моноция ( + )
♂
2. Андомоноция ( +обоеполые)
3. Гиноция (
4. Гермафродит ( обоеполые) AcrAcrgg
♀ ♂ acracrGG
) ♀ AcrAcrGG
acracrgg
По мнению Шифриса, ген «Acr» ускоряет физиологический процесс,
регулируемый полигенами. Дифференциация пола в период развития, таким
образом, по его данным, зависит от генов различных типов цветков, полигенов,
дозы Acr и среды.
Количественный анализ особенностей пола у огурца, численное
соотношение мужских и женских узлов позволил Кубицкому Б. [Kubickii B,
111969]обнаружить гены, взаимодействующие с локусами acr в определении
выраженности пола.
Ген «F» увеличивает выраженность женского пола независимо от
аллельного состава по локусу «acr».
Получены и описаны мужские растения, которые дают только мужские
цветки. Их можно поддержать только с помощью индолил уксусной кислоты.
Установлено.что мужской пол обусловлен одним рецессивным геном «а». Этот
ген проявляет свое действие при наличии гена «acr». Ген «acr F» эпистатичен по
отношению к гену «а». В результате такого взаимодействия растения с генами:
аcr+ acr+ aa – мужские,
acrF acrFaaженские,
acr Facr+aa женские.
Если скрестить гетерозиготные растения с мужскими, можно получить
двудомные растения.
Кубицкий Б. [Kubickii B, 1974] считает,что половые тип у огурца
контролируется четырьмя парами генов: «Mm», «Ff», « Gg», « Hh».
Он также описывает новую форму огурца с женским типом цветения, имеющую
рецессивный характер наследования.
Кооистра Е.[ Kooistra ,1967] предлагает различать генетически «сильные»
и генетически «слабые» женские линии. Он отмечает , что проявление женского
пола в F1 трудно предсказать, основываясь лишь на выраженности женского
пола исходных форм.
Мещеров Э. Т. и Залькалн А.А.[Мещеров, Залькалн , 1967] в популяции
частично женских сортов выделяют генотипически женские растения,
неспособные образовывать мужские цветки под влиянием внешних условий.
Авторы также отмечают, что при опылении генотипически женских растений
пыльцой обычных мужских форм в F1 они наблюдали полностью
фенотипически женское потомство. Позже Мещеров Э.Т.[Мещеров,1975]
отмечает, что у таких растений женский пол обусловлен доминантными генами,
а у растений, образующих мужские цветки под действием раствора
гиббереллина или внешних условий, рецессивными.
12Ткаченко Н. Н. [Ткаченко ,1968]высказывает мнение, что попытки
установить количество генов, контролирующих пол, пока преждевременны. По
его мнению, все семь хромосом гаплоидного набора принимают участие в
определении пола. Он отмечает, что на соотношение растений различных
половых типов в потомстве большое влияние оказывает степень выраженности
женской или мужской наследственной тенденции.
Первоисточником для создания современных материнских форм
гетерозисных гибридов служил японские частично двудомные сорта. [Ткаченко,
1935]. К настоящему времени благодаря труду отечественных и зарубежных
селекционеров создано большое количество материнских форм.
В качестве исходного материала и при выведении материнских форм следует
брать такие сорта, у которых женский пол прочнее генетически закреплен. От
этого в сильной степени зависит успех дела по созданию гетерозисных
гибридов. [Юрина, 1985]
Первоисточником обоеполости является сорт Лемон из США. Плоды у
него мелкие, почти шаровидные, четырех пятикамерные, слегка чалмовидной
формы, мозаичной окраски.
Обоеполость у огурца контролируется одним рецессивным геном. В
первом гибридном поколении, полученном от скрещивания с моноцией, то есть
раздельнополой формой, все растения имеют признаки этой формы, все
раздельнополые.
На признаках плода и на других признаках растения почти не отражается
присутствие в первом гибридном поколении генов обоеполого родителя. Во
втором поколении происходит расщепление по обычной моногибридной схеме
3 моноции: 1 андромоноций.
Путем насыщающих скрещиваний в течении двухтрех поколений с
растениями женского типа можно повысить у обоеполых сортов их скрытое
свойство усиливать проявление женского пола у материнских форм. Это
скрытое свойство у гермафродитных сортов, повидимому, связано с
превалированием на растении гермафродитных цветков за счет сокращения
13мужских, тем сильнее выражено скрытое свойство усиливать проявление
женского пола у гибридной материнской формы. . [Юрина, 1985]
Гермафродитная форма.
Для усиления женского пола материнских форм при получении
гетерозисных гибридов создают гермафродитные или андромоноцийные формы,
с использованием которых сокращаются затраты на прочистки мужских
растений на материнских сортах. Тройные гибриды, полученные на их основе,
более урожайны и более пластичны по приспособительным возможностям, чем
простые гибриды.
Селекционную работу по выведению этих форм целесообразно по
возможности упростить, применяя метод создания аналогов (В. Кубицкий).
Андромоноцийный или гермафродитный сорт подвергают насыщающим
скрещиваниям в двух трех поколениях с материнской формой женского типа,
сопровождая в каждом поколении соответствующим отбором.
Когда сорт приобретает по внешнему плода признаки материнской формы,
он становится ее гермафродитным аналогом.
При выведении нового обоеполого сорта, предназначенного для усиления
женского пола, исходный обоеполый сорт скрещивается с одним из
гинойкистных. Он должен быть достаточно проверенным в качестве
материнской формы и соответствовать по хозяйственно ценным признакам
избранному направлению селекции. Для этого скрещивания отбираются
растения с наиболее сильно выраженной женской тенденцией генетически
женские. Признак обоеполости у огурца является рецессивным по отношению к
признаку раздельнополости.
Сложная материнская форма (СМФ).
СМФ (женская форма × гермафродитный аналог) получают сначала в
научных учреждениях, а при расширении семеноводства новых районированных
гибридов в семеноводческих хозяйствах или на опытных станциях. Посевы
проводятся в соотношении 2:1 или 3:1, где гермафродитный аналог
используется как отцовская форма.
14На посевах материнской формы тщательно проводят сортовые прочистки,
так как от её чистоты зависит гибридность семян. На рядках материнской
формы получают семена СМФ, которые затем используют в скрещиваниях с
однодомными сортами.
1.5 Горечь
У огурца качество товарного урожая сильно снижается из – за горечи
плодов, характерной почти для всех гибридов и сортов, выращиваемых в нашей
стране в открытом и закрытом грунте.
J. M. Andeweg, J. W. De Brayn (1959) делят все сорта огурца по
образованию в них горьких веществ на три группы:
1) вегетативные части содержат горечь, в плодах при определенных условиях
также проявляется горечь;
2) вегетативные части содержат горечь, а в плодах она не проявляется даже при
неблагоприятных условиях;
3)горечь отсутствует как в вегетативных частях , так и в плодах.
Последняя группа сортов и представляет наибольший интерес как
исходных материал в селекции при создании форм без горечи, отсутствие
которой обусловлено генетически.
Согласно литературным данным(Barham,1953, Andeweg , De Brayn ,
1959,Borghi 1972) отсутствие горечи контролируется рецессивным геном
bi(bitter free),горечь доминантным Bi .Наши исследования также подтвердили,
что отсутствие горечи наследуется рецессивно монофакториально. Оценивали
растения на наличие горечи в фазе сеянцев органолептическим способом.
В первом поколении гибридов, полученных от скрещивания горьких и
негорьких форм, горечь в растениях доминирует. Во втором поколении
гибридов наблюдается расщепление по фенотипу в соотношении, очень близком
к теоретически ожидаемому 3:1,в беккросном потомстве 1:1[Стрельникова,
Маштакова,1984]
В закрытом грунте горечь у огурцов проявляется при неблагоприятных
условиях выращивания (низкие и высокие температуры, повышенная и
пониженная освещенность, сухость воздуха и т.д).
15У горьких форм (BiBi, Bibi)горечь обычно проявляется с момента начала
прорастания семян, наиболее сильно выражена она в фазе семядолей, несколько
меньше в листьях и других вегетативных органах. [Стрельникова, 1977]
1.6Опушение завязи
Завязь по форме может быть яйцевидной, веретеновидной,
цилиндрической и эллипсовидной. Опушение завязи трех типов: простое
волоски (шипики) сидят непосредственно на поверхности завязи; сложное
волоски сидят на пузырчатых вздутиях, смешанное имеют волоски простого и
сложного строения. [Юрина, 1985]
.
2 Материал и методы исследования.
Экспериментальная часть работы выполнена в отделе генетических
ресурсов и селекции овощных культур Крымской опытной станции ВНИИР в
20092010 гг. в период прохождения учебной и специальной производственной
практик.
Личное участие в получении материала для дипломной работы состояло в
выполнении части работы по опылению при постановке скрещиваний, но
главным образом, в описании гибридов F2 в трех различных вариантах опыта и
их выращивания по комплексу признаков, включающих тип цветка, а также
некоторые признаки качества плода. Общее число учтенных признаков пять. В
дальнейшем именно они и подлежали генетическому анализу.
В качестве родительских формродоначальников семейгибридов
выступали сортообразцы, а иногда достаточно сложные гибриды из
селекционной коллекции отдела селекции: сортообразец 788 (во всех
16комбинациях скрещивания он выступал в качестве материнской формы);
748×745;660, [F4 Пасамонте×(723×(791×736) In 2].
Фенотипические характеристики родительских форм по пяти
учитываемым признакам сведены в табл.1. В 2009 г. исследовались три
гибридные комбинации : 788×[748×745], 788×660, 788×[F4
Пасамонте×(723×(791×736) In 2]. Первые две из указанных были повторены в
2010 г.
Таким образом за два года исследования анализу подлежали гибриды пяти
семей. Четыре признака: тип цветка, окраска плода, характер опушения завязи,
тип кожицы плода, описывали визуально, наличие горечи органолептически.
При производстве скрещиваний с последующим описанием расщепления
руководствовались пособием «Методические указания по селекции огурца»
О.В. Юриной: «…Среди отобранных лучших растений подбирают сходные
между собой и искусственно опыляют смесью пыльцы с нескольких цветков
одного растения. Семена собирают с каждого растения отдельно. При
использовании этого метода каждое растение должно иметь свой порядковый
номер для индивидуальной оценки и искусственного скрещивания.
Наиболее удобна следующая схема размещения на делянках питомников в
открытом грунте (в защищенном грунте нумеруются все растения): в каждом
ряду по 10 гнезд, в гнезде по 2 растения, на делянке по 34 ряда. Растения
нумеруют так: в первом ряду с 1 по 10, во втором с 11 по 20 и т.д. пара
растений в гнезде обозначается буквами «а» и «б», причем то растение в гнезде,
которое расположено ближе к началу ряда, т.е. ближе к деляночной этикетке,
отмечают буквой «а». Так как расстояние между гнездами в ряду довольно
большое 70 см, то нетрудно определить порядковый номер каждого растения.
Описание и первичный отбор в питомниках целесообразно проводить
следующим образом: описывать и оценивать каждую делянку по трем сборам
зеленцов; все растения описывать по выраженности пола, а затем по наиболее
интересным делянкам после второго или третьего сбора зеленцов описывать и
оценивать лучшие растения индивидуально, уточняя при этом план проведения
17искусственных опылений. Для лучшей сохранности ценного материала на
первом этапе селекции селекционные питомники следует располагать в
пленочных теплицах.
Отбор по признакам выраженности пола эффективен при выращивании
растении в условиях, когда женский пол проявляется более слабо: в
защищенном грунте при летнеосенней и осеннее зимней культуре. Растения,
отобранные по признаку женского пола в этот период, сохраняет да признак в
потомстве, такие условия позволяют выделить генетические женские
растения.» [Юрина, 1985]
Статистическую обработку данных производили, используя критерий хи
квадрат К. Пирсона, который применяли как при оценке однородностей
расщепления в отдельных вариантах опыта (или семьях), так и при оценке
генетических гипотез о природе расщепления, т.е. при сравнении эмпирического
и теоретически ожидаемого расщеплений. Необходимо указать на важную
особенность применения в первом случае: при сравнении эмпирических
расщеплений на предмет отсутствия между ними статистически достоверных
различий(H0) допускается сравнение Fкритерия как со стандартным значением
3,84 для 5%го уровня значимости, так и 1%го6,64;поскольку 1% уровень при
проверке данной H0 соответствует не повышенной, а обычной надежности
статистического прогноза.
Вычисления выполнены с использованием калькулятора.
3 Результаты и обсуждение
На первом этапе анализа решался вопрос о возможности объединить
данные по делянкам каждой семьи. Для его решения использовали специальную
модификацию критерия χ2, которая позволяет сравнить два и более
эмпирических отношения расщепления.
Рассмотрим для примера расщепление на делянках по признаку тип цветка
(таблица 3).
18Номер
семьи
(делянки)
1
(1,2)
2
(3, 4, 5)
3
(6, 7, 8, 9)
Суммарно
по семье
20: 4
14: 0
18: 6
24: 6
13: 6
22: 9
19: 2
18: 5
13: 3
161: 41
21,4: 2,5
12, 4: 1,5
18,1: 5,9
22,6: 7,4
14,3: 4,7
28,8: 7,6
22,3:5,9
24,5:6,5
19,4: 5,1
На основе
модели 3: 1
166,5: 55,5
34: 4
55: 18
72: 19
161: 41
4,41;
˂
р 0,01
6,16;
˂
р 0,05
6,27;
˂
р 0,05
3,18;
˂
р 0,05
Таблица 3. Результаты эмпирических отношений расщепления по признаку
«тип цветка: » (раздельнополый: обоеполый) на различных делянках
выращивания гибридов огурца трех семей F2.
Отношения расщепления
эмпирическое
Теоретически
ожидаемое
Суммарно
по семье
χ 2; pH0
Из последней колонки таблицы, где приведены значения χ2 и вероятность
0гипотезы о сходстве отношения расщепления (рН0) видно, что во всех
случаях объединение данных по разных делянкам возможно.
Суммарное по семье отношение 161:41 статистически достоверно не
отличается от теоретически ожидаемого , на основе модели 3:1, 166,5:55,5. Это
означает, что расщепление определяется одним геном, женский тип цветка при
этом доминирует.
Данные таблицы 4 также указали на возможность объединения материала
по делянкам. Расщепление по окраске плода (зеленый белый) также
моногенно, зеленая окраска доминирует.
Таблица 4. Результаты сравнения эмпирических отношений расщепления по
признаку «окраска плода» (зеленый: белый) на различных делянках
выращивания гибридов огурца трех семей F2
19Номер
семьи
(делянки)
1
(1,2)
2
(3, 4, 5)
3
(6, 7, 8, 9)
Суммарно
по семье
Отношения расщепления
эмпирическое
Теоретически
ожидаемое
Суммарно
по семье
χ 2; pH0
23: 1
14: 0
18: 6
27: 3
16: 3
25: 6
16: 5
17: 6
12:4
168: 34
37: 1
61: 12
70: 21
168: 34
0,68;
˂
р 0,01
2,25;
˂
р 0,05
0,42;
˂
р 0,05
0,015,
˂
р 0,05
23,31: 0,6
13,69: 0,4
20,1: 3,9
25,0:4,9
15,9: 3,1
23,8: 7,1
16,1: 4,8
17,5: 5,2
11,9: 3,6
На основе
модели 3:1
167,3: 33,6
Данные таблицы 5 также позволили объединить делянки по семьям (показать
р›0,01), но суммарное по семье отношение расщепление указало на
расщепление по 2м генам, взаимодействующим по типу рецессивного эпистаза.
Соответствующая математическая модель 9:7. Значение χ2 =0,97(показать)
подтверждает такой дигенный контроль генетического признака.
Таблица 5. Результаты сравнения эмпирических отношений расщепления по
признаку «тип опушения» (простое: сложное) на различных делянках
выращивания гибридов огурца трех семей F2
20Номер
семьи
(делянки)
1
(1,2)
2
(3, 4, 5)
3
(6, 7, 8, 9)
Суммарно
по семье
Отношения расщепления
эмпирическое
Теоретически
ожидаемое
Суммарно
по семье
χ 2; pH0
17: 7
9: 5
14: 10
24: 6
13: 6
15: 16
10: 11
10: 13
9: 7
121: 81
26: 12
51: 22
44: 47
121:81
16,4: 9,7
7,6: 4,4
16,8: 7,3
20,9: 9.0
13,3: 5,7
13,2: 14,1
10,1:10,8
11:11,7
7,5: 7,5
На основе
модели 3:1
168,34: 80,2
0,2;
˂
р 0,01
2,9;
˂
р 0,05
0,82;
˂
р 0,05
13,32,;
˂
р 0,05
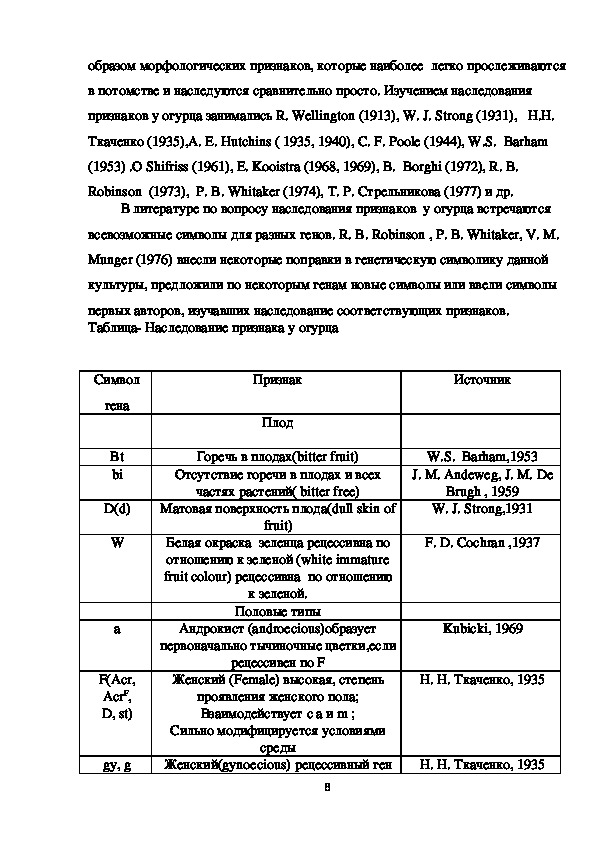
Данные таблицы 6 указали на моногенность признака поверхность плода,
матовая доминирует.
Таблица 6. Результаты сравнения эмпирических отношений расщепления по
признаку «поверхность плода» (матовая: глянцевая) на различных делянках
выращивания гибридов огурца трех семей F2
Номер
семьи
(делянки)
1
(1,2)
2
(3, 4, 5)
3
(6, 7, 8, 9)
Суммарно
по семье
Отношения расщепления
эмпирическое
Теоретически
ожидаемое
Суммарно
по семье
16: 8
11:3
21:3
30:1
18: 1
23: 8
20:1
20: 3
16: 0
175: 28
27: 11
69: 5
79: 12
175: 28
17,0:6,9
9,9: 4,1
22,1: 1,6
29,0: 2,1
17,9: 1,3
26,9: 4,1
18,2: 2,8
19,7: 3
13,4 :1,2
На основе
модели 3:1
21
174,1: 27,1
χ 2;
pH0
0,65;
˂
р 0,01
1,96;
˂
р 0,05
7,32;
˂
р 0,05
0,03,
˂
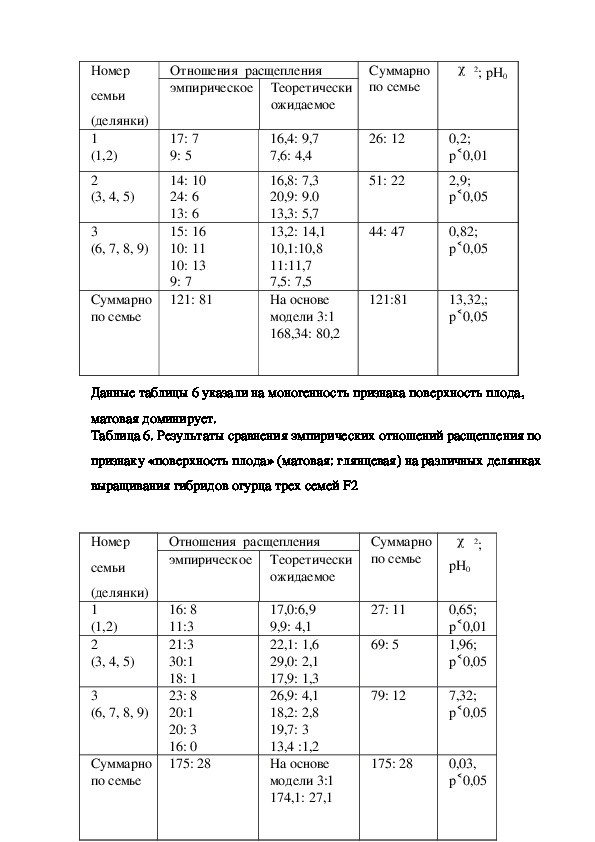
р 0,05Единственное затруднение представил анализ признака наличие горечи.
Делянки по семье 3 объединить не удалось, χ2=21,8 при р 0,05(показать). В
семье выделено два совершенно разных типа расщепления: 0:54 ,13:24. Причина
такого результата остается пока неясной. Суммарное по двум первым семьям
˂
отношение расщепления 90:21(показать) оказалось моногенным, модель 3:1, где
отсутствие горечи признак рецессивный.
Таблица 7. Результаты сравнения эмпирических отношений расщепления по
признаку «вкус» (горький: без горечи) на различных делянках выращивания
гибридов огурца трех семей F2
Номер
семьи
(делянки)
1
(1,2)
2
(3, 4, 5)
3
(6, 7, 8, 9)
Суммарно
по семье
Отношения расщепления
эмпирическое
Теоретически
ожидаемое
Суммарно
по семье
15: 9
8: 6
20: 4
29: 1
18: 1
0: 31
7: 14
0: 23
6: 10
103: 99
23: 15
67: 6
13: 78
103: 99
14,5: 9,4
8,5: 5,5
22,1: 2,0
27,5: 2,5
17,4: 1,6
4,4: 26,5
3,0: 18,0
3,2: 19,5
2,2: 13,3
На основе
модели 1:1
102,8: 98,3
χ 2;
pH0
0,11;
˂
р 0,01
3,42;
˂
р 0,05
21,8;
˂
р 0,05
3,89,
˂
р 0,05
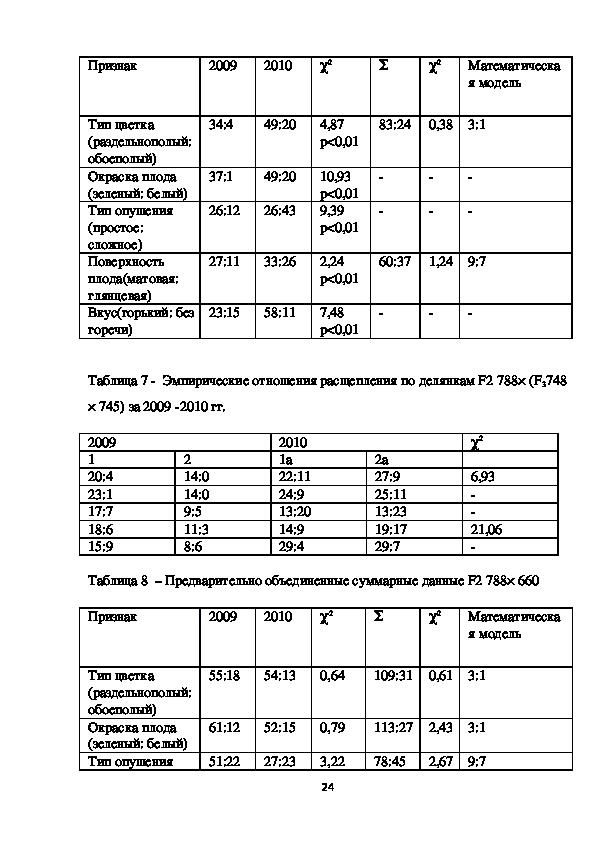
Таблица 4 Объединение данных по делянкам F2 788× 660 (2010 г.)
Признак
Отношение
расщепления
χ2
22Тип цветка
(раздельнополый:
обоеполый)
Окраска плода
(зеленый: белый)
Тип опушения
(простое: сложное)
Поверхность
плода(матовая:
глянцевая
Вкус(горький: без
горечи)
26:7
28:6
27:6
25:9
16:0
11:23
17:16
17:17
29:4
28:6
0,09
p < 0,01
0,68
p<0,01
20,04
р<0,01
0,02
р<0,01
0,4
р<0,01
Таблица 5 Объединение данных по делянкам F2 788× (F3748 × 745); 2010 г.
Признак
Тип цветка
(раздельнополый:
обоеполый)
Окраска плода
(зеленый: белый)
Тип опушения
(простое: сложное)
Поверхность
плода(матовая:
глянцевая)
Вкус(горький: без
горечи)
Отношение
расщепления
22:11
27:9
24:9
25:11
13:20
13:23
14:9
19:17
29:4
29:7
χ2
0,58
р<0,01
0,09
р<0,01
0,08
0,37
р<0,01
0,69
р<0,01
Данные χ2 таблицы 4 и 5 указали на возможность объединения материала по
делянкам, кроме признака «тип опушения» в семье F2 788× 660, где χ2=20,04,
р<0,01. Прошлогодний расчет указал на дигенный контроль признака по типу
рецессивного эпистаза.
Таблица 6 – Предварительно объединенные суммарные данные F2 788× (F3748 ×
745) за 2009 и 2010 гг.
23Признак
2009
2010
χ2
Σ
χ2
Математическа
я модель
Тип цветка
(раздельнополый:
обоеполый)
Окраска плода
(зеленый: белый)
Тип опушения
(простое:
сложное)
Поверхность
плода(матовая:
глянцевая)
Вкус(горький: без
горечи)
34:4
49:20
37:1
49:20
26:12
26:43
27:11
33:26
23:15
58:11
4,87
р<0,01
10,93
р<0,01
9,39
р<0,01
2,24
р<0,01
7,48
р<0,01
83:24
0,38
3:1
60:37
1,24
9:7
Таблица 7 Эмпирические отношения расщепления по делянкам F2 788× (F3748
× 745) за 2009 2010 гг.
2009
1
20:4
23:1
17:7
18:6
15:9
2
14:0
14:0
9:5
11:3
8:6
2010
1а
22:11
24:9
13:20
14:9
29:4
2а
27:9
25:11
13:23
19:17
29:7
χ2
6,93
21,06
Таблица 8 – Предварительно объединенные суммарные данные F2 788× 660
Признак
2009
2010
χ2
Σ
χ2
Математическа
я модель
Тип цветка
(раздельнополый:
обоеполый)
Окраска плода
(зеленый: белый)
Тип опушения
55:18
54:13
0,64
109:31
0,61
3:1
61:12
52:15
0,79
113:27
2,43
3:1
51:22
27:23
3,22
24
78:45
2,67
9:7(простое:
сложное)
Поверхность
плода(матовая:
глянцевая)
Вкус(горький: без
горечи)
69:5
34:33
31,91
67:6
57:10
1,55
124:16
6,41
15:1
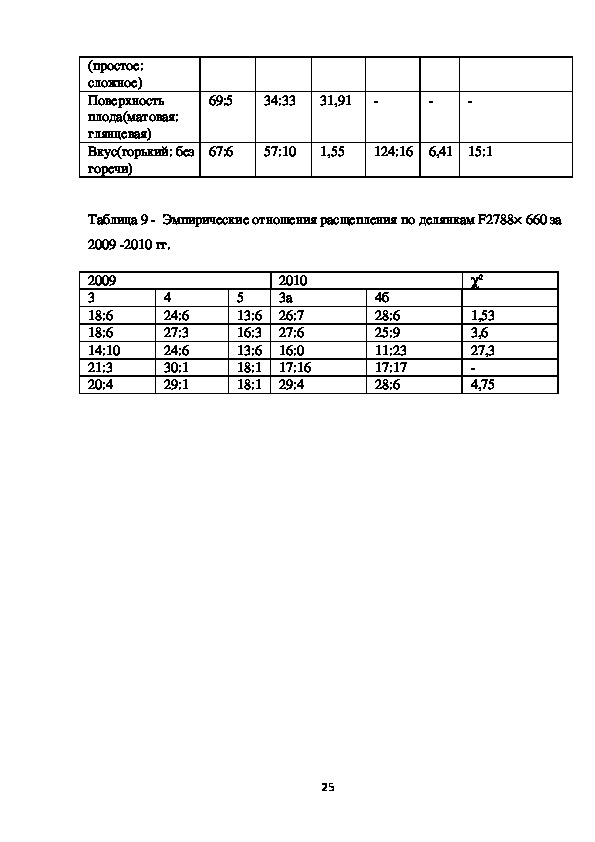
Таблица 9 Эмпирические отношения расщепления по делянкам F2788× 660 за
2009 2010 гг.
2009
3
18:6
18:6
14:10
21:3
20:4
4
24:6
27:3
24:6
30:1
29:1
2010
3а
26:7
27:6
16:0
17:16
29:4
5
13:6
16:3
13:6
18:1
18:1
4б
28:6
25:9
11:23
17:17
28:6
χ2
1,53
3,6
27,3
4,75
254. ЗАКЛЮЧЕНИЕ
Таким образом, по четырём признакам из пяти изученных доказана
моногенность генетического контроля, исключение составляет опушение
кожицы плода, которое контролируется по типу двойного рецессивного
эпистаза.
Когда генетический контроль признаков, желательные варианты которых
требуется соединить в будущей селекционной форме, открывается
возможность построить селекцию по принципу планомерного конструирования
будущего сорта на основе данных генетического анализа. Решение такой задачи
представляется не только возможным, но и не очень сложным. Особенно, если
желательные варианты признака являются рецессивными. Отбор рецессивных
гомозигот проводится совершенно однозначно.
СПИСОК ЛИТЕРАТУРЫ
261.Даскалов Х., Михов А. [и др]. Гетерозис и его использование в овощеводстве.
Пловдив,1975. С.137139.
2.Ерёмин В.Г., Гасанова Т. А., Подорожный В.Н. Крымская опытно
селекционная станция (к 75летию образования). Крымск,2010.283 с.
Жимулёв И. Ф. Общая и молекулярная генетика. Новосибирск, 2002. 457 с.
3.Медведев А. А. Сорта и гибриды огурца для летних сроков
посева//Сохранение и использование генофонда в селекции овощных и плодово
ягодных культур на юге России (тезисы докладов и выступлений на
международной научно практической конференции). Крымск,2000. С.4648
4.Мещеров Э. Т. Огурцы / Мещеров Э. Т., Босс Г. В. Л., 1967.
5.Мещеров Э.Т. Основные задачи селекции и новые методы семеноводства
гетерозисных гибридов огурцов// Методы ускорения селекции овощных
культур. Л., 1975 с. 7475
6. Прохоров И. А., Крючков А. В. Селекция и семеноводство овощных культур,
М.,1997 с. 262
7.Ткаченко Н. Н. Огурцы М., 1963г.
8.Ткаченко Н. Н. Предварительные итоги генетического изучения огурцов//
Труды по прикладной ботанике, генетике и селекции. Серия II, 9, 1935, с. 311
355
9.Ткаченко Н. Н. Методы выведения частично двудомных форм//Гетерозис в
овощеводстве. М., 1968 с. 166171.
10.Ткаченко Н. Н. Генетика гибридных огурцов//Научные труды Крымской
опытной станции. Крымск, 1973г., с 34.
2711.Стрельникова Т. Р. Использование гетерозиса в селекции тепличных огурцов.
Кишинев, 1977, с. 2234
12. Стрельникова Т. Р., Маштакова А. Х., Гусева Л. И. Селекция гетерозисных
гибридов огурца. Кишинев, 1984
13.Шевелуха В. С. Методические указания по селекции и семеноводству
гетерозисных гибридов огурца М.,1985. С.717.
14. Corrence T. Nodal sequence of flower type in cucumber. 1932, Proc. Amer. Soc.
Hort. Sci. 29. P. 477479
15. Kooistra E. Euphytica.1967,P. 117
16. Kubicki B . Proc. XIX Jnternat, Hort, Congr 3.1974, P.475485
17. Kubicki B. Investigations on sex determination in cucumber (Cucumis sativis
L) .V Genes controlling intensity of femaleness. Genetica Polonica , VI
Androecism. Genetica Polonica, 1969, P.8799.
18. Shifriss O. Sex control in cucumbers. Journal of Heredity , 1961, P. 512
19.Огурец обыкновенный/ Википедия свободная энциклопедия. URL:
http://ru.wikipedia.org/wiki/Cucumis_sativus(дата обращения: 15.03.2011)
20. Видова О. Главный огуречник страны/ URL:
http://www.roskrup.ru/articles/articles.php?id=257(дата обращения:27.04.2011)
21. Биометрический анализ качественных признаков в селекции растений/
URL:http://www.library.timacad.ru/download/genetics/9.pdf
05.2011)
(дата обращения: 5.
22.Значения критерия Пирсона (хи - квадрат) при разных уровнях
значимости/ URL: http://www.vertcha.ru/Files/hi.pdf (дата
обращения:5.05.2011)
283.1 Сравнительностатистический анализ отношений расщепления в отдельных
опытах по гибридизации с целью формирования репрезентативного материала
для генетического анализа
Огромный опыт генетического анализа отдельных признаков самых
различных объектов убедил генетиков в целесообразности его проведения на
гибридном материале большого объёма. Так называемая теорема «больших
чисел», перефразированная применительно генетикоаналитическим
исследованиям, звучит так: частоты разных фенотипических, а, следовательно, и
генотипических классов расщепления, тем точнее соответствует их вероятности,
чем больше число изученных гибридов. Первым из генетиков в этом убедился
уже Г. Мендель, 1926 [цит. по Жимулёву, 2002]. Лишь суммировав перед
анализом гибридов F2 частоты одноименных классов по нескольким семьям, он
получил отношение расщепления хорошо соответствующее ожидаемой
математической модели расщепления.
Следует особо подчеркнуть, что проведение генетического анализа на
материале, полученном селекционерами, значительно отличается от того,
которое выполняется на данных специально спланированного эксперимента.
Действительно, селекционеры планируют систему скрещиваний и описывают
гибриды с одной единственной целью отобрать среди них так называемые
селекционноценные формы. Полноценное проведение генетического анализа не
входит в их задачу. Поэтому генетик, обратившись к селекционному материалу,
должен проделать значительную подготовительную работу для того, чтобы
правильно объединить или, наоборот, разделить полученные гибридные семьи, а
также учесть их различие, обусловленное выращиванием гибридов в несколько
различных условиях, например, на разных делянках селекционного поля, в
разные годы и т.д.
С такой подготовительной работы и было начато данное исследование.
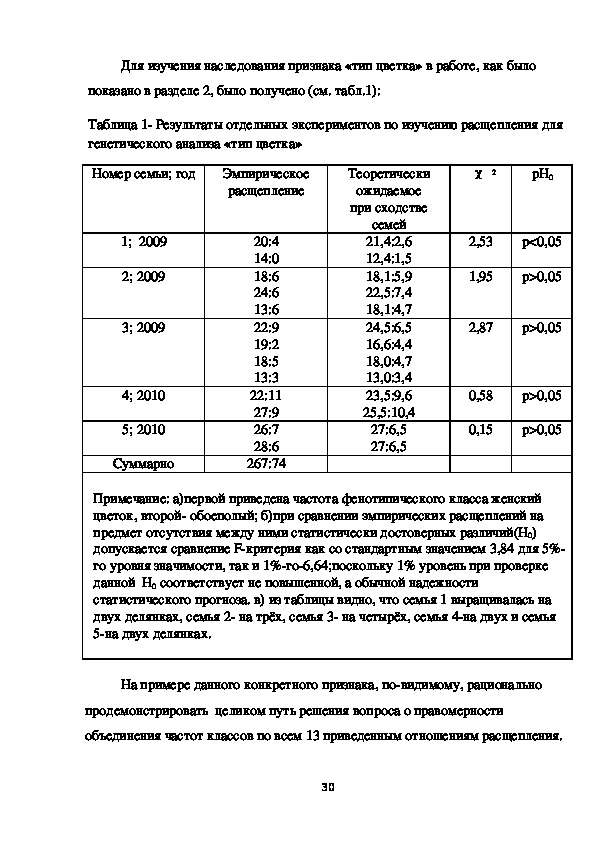
29Для изучения наследования признака «тип цветка» в работе, как было
показано в разделе 2, было получено (см. табл.1):
Таблица 1 Результаты отдельных экспериментов по изучению расщепления для
генетического анализа «тип цветка»
Номер семьи; год
Эмпирическое
расщепление
Теоретически
ожидаемое
при сходстве
1; 2009
2; 2009
3; 2009
4; 2010
5; 2010
Суммарно
20:4
14:0
18:6
24:6
13:6
22:9
19:2
18:5
13:3
22:11
27:9
26:7
28:6
267:74
семей
21,4:2,6
12,4:1,5
18,1:5,9
22,5:7,4
18,1:4,7
24,5:6,5
16,6:4,4
18,0:4,7
13,0:3,4
23,5:9,6
25,5:10,4
27:6,5
27:6,5
χ 2
pH0
2,53
1,95
р<0,05
р>0,05
2,87
р>0,05
0,58
0,15
р>0,05
р>0,05
Примечание: а)первой приведена частота фенотипического класса женский
цветок, второй обоеполый; б)при сравнении эмпирических расщеплений на
предмет отсутствия между ними статистически достоверных различий(H0)
допускается сравнение Fкритерия как со стандартным значением 3,84 для 5%
го уровня значимости, так и 1%го6,64;поскольку 1% уровень при проверке
данной H0 соответствует не повышенной, а обычной надежности
статистического прогноза. в) из таблицы видно, что семья 1 выращивалась на
двух делянках, семья 2 на трёх, семья 3 на четырёх, семья 4на двух и семья
5на двух делянках.
На примере данного конкретного признака, повидимому, рационально
продемонстрировать целиком путь решения вопроса о правомерности
объединения частот классов по всем 13 приведенным отношениям расщепления.
30При сравнении семей с одинаковым расщеплением мы исходили из старой
истины: различными могут быть только неверные решения, а верное – одно.
Сравнение двух или более распределений на предмет их сходства или
различия строится на определении вероятности H0гипотезы об отсутствии
различий. Для этого в качестве теоретически ожидаемой частоты классов
выбирается одно и то же число, а именно сумма частот соответствующего
класса по всем сопоставляемым распределениям. Поскольку эта сумма
естественно больше частоты любого из классов в конкретном распределении,
устанавливается их соответствия путем умножения на поправочный
коэффициент. Он равен доле конкретного распределения, вносимой в
суммарный результат. После этого вычисляются теоретически ожидаемые
частоты и величина χ 2 , которая оценивается при df =(a1) (k1), где а число
классов в расщеплении, k число сопоставляемых распределений.
Для двух делянок семьи 1 (2009 г. учета) соответствующая таблица имеет
вид(см.таблица 2):
Таблица 2 Пример сравнения двух эмпирических распределений на предмет
наличия статистически достоверных различий
Эмпирическое
распределение
20:4
14:0
∑ 34:4
Поправочные
коэффициенты
34/38=0,89
4/38=0,11
Суммы по классам
Теоретически ожидаемые
частоты
24
14
38
Теоретически
ожидаемое:
24×0,89=21,3
24×0,11=2,6
21,3:2,6
12,4:1,5
χ 2 =2,53
р<0,05
Н0 принимается
Абсолютно аналогичным образом сопоставлены расщепления на трёх
делянках в семье 2, четырёх делянках, где выращивалась семья 3, двух
делянках семей 4 и 5.
31Судя по фактическим значениям критерия χ 2 во всех случаях есть
статистические основания объединить материал по делянкам всех семей (см.
табл. 1).
Оставалось сравнить отношения расщепления в гибридах пяти семей,
полученных в результате скрещивания разных родительских форм в различные
годы.
Таблица 3, построенная по принципу табл. 2, приводится:
Таблица 3 Результаты сравнения эмпирических отношений расщепления в
тринадцати вариантах эксперимента, подлежащих генетическому анализу
признака «тип цветка» огурца
Эмпирическое распределение в различных
Сумма
Теоретически определяемые
34
4
∑ 38
55
18
73
семьях
72
19
91
49
20
69
48
13
61
по
классам
258
74
332
28,4
8,1
69,7
20,0
56,8
54,2
16,3
15,5
χ 2=4,67< χ 2
05=9,49
49,0
14,1
Поправочные коэффициенты
38/332
=0,11
73/332
=0,22
91/332
=0,27
69/332
=0,21
р>0,05
Н0 принимается
61/332
=0,19
Результаты сравнения пяти сопоставляемых отношений расщепления
отражен в фактическом значении χ 2 , которое уступает стандартному для
5%го уровня значимости (см.табл. 3). Это означает, что все 13 сопоставляемых
отношений расщепления по признаку «тип цветка» статистически правомерно
могут быть объединены, и генетическому анализу может подлежать суммарное
по делянкам, семьям и годам отношения расщепления: 264 женских: 74
обоеполых.
Методы и путь сравнения результатов разных экспериментов по
скрещиванию на примере альтернативных классов: женский цветок обоеполый
цветок изложены достаточно подробно для того, чтобы при исследовании
32расщепления по другим изучаемым признакам можно было бы ограничиться
приведением только финальных таблиц.
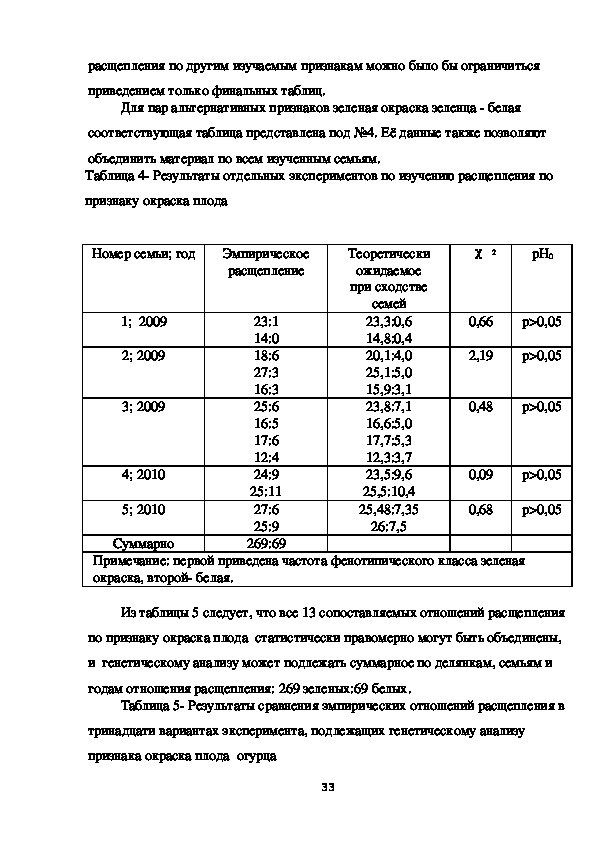
Для пар альтернативных признаков зеленая окраска зеленца белая
соответствующая таблица представлена под №4. Её данные также позволяют
объединить материал по всем изученным семьям.
Таблица 4 Результаты отдельных экспериментов по изучению расщепления по
признаку окраска плода
Номер семьи; год
Эмпирическое
расщепление
1; 2009
2; 2009
3; 2009
4; 2010
5; 2010
Суммарно
23:1
14:0
18:6
27:3
16:3
25:6
16:5
17:6
12:4
24:9
25:11
27:6
25:9
269:69
Теоретически
ожидаемое
при сходстве
семей
23,3:0,6
14,8:0,4
20,1:4,0
25,1:5,0
15,9:3,1
23,8:7,1
16,6:5,0
17,7:5,3
12,3:3,7
23,5:9,6
25,5:10,4
25,48:7,35
26:7,5
χ 2
pH0
0,66
2,19
р>0,05
р>0,05
0,48
р>0,05
0,09
0,68
р>0,05
р>0,05
Примечание: первой приведена частота фенотипического класса зеленая
окраска, второй белая.
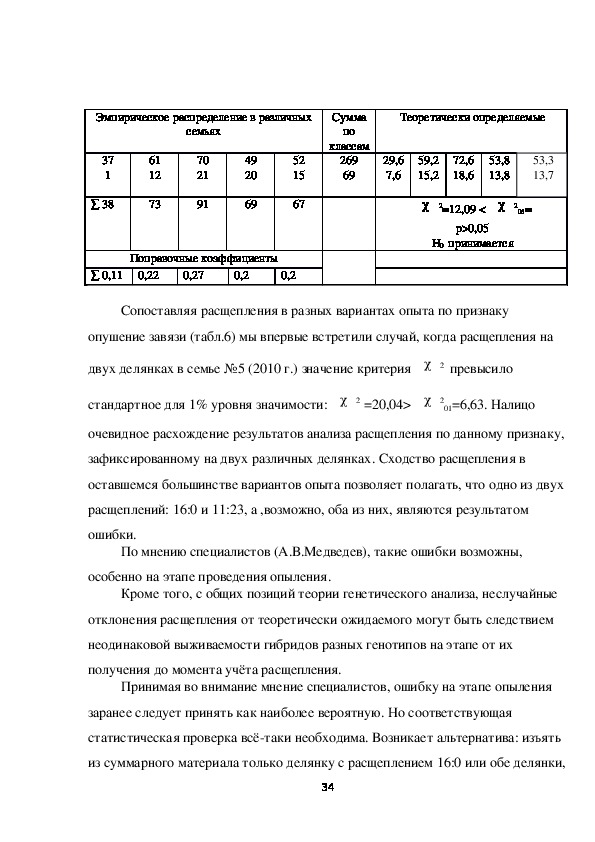
Из таблицы 5 следует, что все 13 сопоставляемых отношений расщепления
по признаку окраска плода статистически правомерно могут быть объединены,
и генетическому анализу может подлежать суммарное по делянкам, семьям и
годам отношения расщепления: 269 зеленых:69 белых.
Таблица 5 Результаты сравнения эмпирических отношений расщепления в
тринадцати вариантах эксперимента, подлежащих генетическому анализу
признака окраска плода огурца
33Эмпирическое распределение в различных
семьях
37
1
∑ 38
61
12
73
70
21
91
49
20
69
52
15
67
Сумма
по
классам
269
69
Теоретически определяемые
29,6
7,6
59,2
15,2
72,6
18,6
53,8
13,8
53,3
13,7
χ 2=12,09 < χ 2
05=
р>0,05
Н0 принимается
Поправочные коэффициенты
∑ 0,11
0,22
0,27
0,2
0,2
Сопоставляя расщепления в разных вариантах опыта по признаку
опушение завязи (табл.6) мы впервые встретили случай, когда расщепления на
двух делянках в семье №5 (2010 г.) значение критерия χ 2 превысило
стандартное для 1% уровня значимости: χ 2 =20,04> χ 2
01=6,63. Налицо
очевидное расхождение результатов анализа расщепления по данному признаку,
зафиксированному на двух различных делянках. Сходство расщепления в
оставшемся большинстве вариантов опыта позволяет полагать, что одно из двух
расщеплений: 16:0 и 11:23, а ,возможно, оба из них, являются результатом
ошибки.
По мнению специалистов (А.В.Медведев), такие ошибки возможны,
особенно на этапе проведения опыления.
Кроме того, с общих позиций теории генетического анализа, неслучайные
отклонения расщепления от теоретически ожидаемого могут быть следствием
неодинаковой выживаемости гибридов разных генотипов на этапе от их
получения до момента учёта расщепления.
Принимая во внимание мнение специалистов, ошибку на этапе опыления
заранее следует принять как наиболее вероятную. Но соответствующая
статистическая проверка всётаки необходима. Возникает альтернатива: изъять
из суммарного материала только делянку с расщеплением 16:0 или обе делянки,
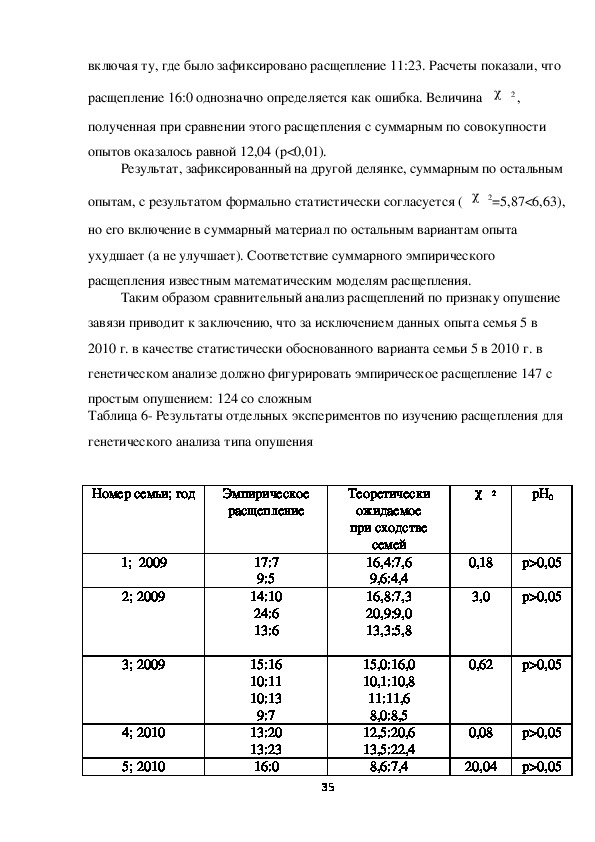
34включая ту, где было зафиксировано расщепление 11:23. Расчеты показали, что
расщепление 16:0 однозначно определяется как ошибка. Величина χ 2 ,
полученная при сравнении этого расщепления с суммарным по совокупности
опытов оказалось равной 12,04 (р<0,01).
Результат, зафиксированный на другой делянке, суммарным по остальным
опытам, с результатом формально статистически согласуется ( χ 2=5,87<6,63),
но его включение в суммарный материал по остальным вариантам опыта
ухудшает (а не улучшает). Соответствие суммарного эмпирического
расщепления известным математическим моделям расщепления.
Таким образом сравнительный анализ расщеплений по признаку опушение
завязи приводит к заключению, что за исключением данных опыта семья 5 в
2010 г. в качестве статистически обоснованного варианта семьи 5 в 2010 г. в
генетическом анализе должно фигурировать эмпирическое расщепление 147 с
простым опушением: 124 со сложным
Таблица 6 Результаты отдельных экспериментов по изучению расщепления для
генетического анализа типа опушения
Номер семьи; год
Эмпирическое
расщепление
Теоретически
ожидаемое
при сходстве
1; 2009
2; 2009
3; 2009
4; 2010
5; 2010
17:7
9:5
14:10
24:6
13:6
15:16
10:11
10:13
9:7
13:20
13:23
16:0
семей
16,4:7,6
9,6:4,4
16,8:7,3
20,9:9,0
13,3:5,8
15,0:16,0
10,1:10,8
11:11,6
8,0:8,5
12,5:20,6
13,5:22,4
8,6:7,4
35
χ 2
pH0
0,18
р>0,05
3,0
р>0,05
0,62
р>0,05
0,08
р>0,05
20,04
р>0,05Суммарно
11:23
174:147
18,4:15,6
Примечание: первой приведена частота фенотипического класса простое
опушение, второй сложное
Таблица 7 Результаты сравнения эмпирических отношений расщепления в
тринадцати вариантах эксперимента, подлежащих генетическому анализу
признака опушение завязи
Эмпирическое распределение в различных
семьях
Сумма
по
классам
Теоретически определяемые
∑
Поправочные коэффициенты
χ 2= < χ 2
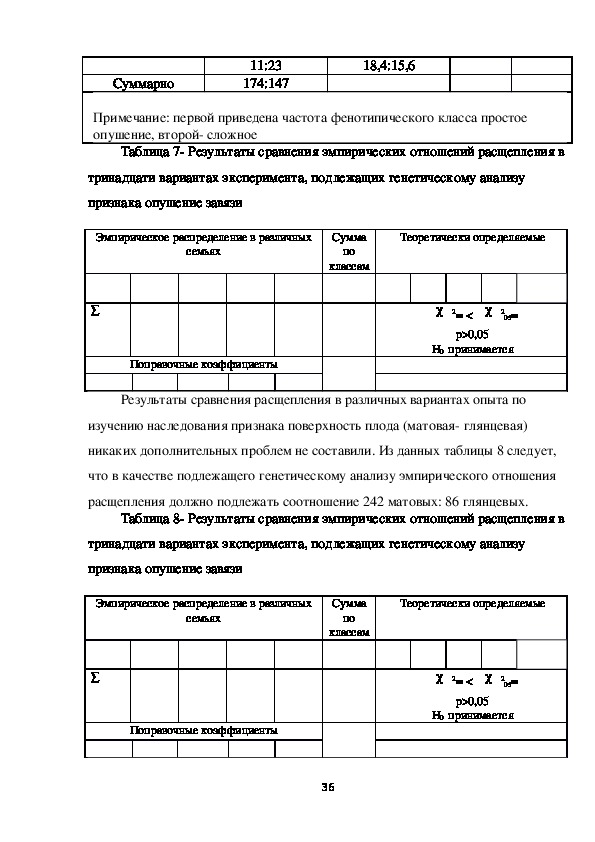
05=
р>0,05
Н0 принимается
Результаты сравнения расщепления в различных вариантах опыта по
изучению наследования признака поверхность плода (матовая глянцевая)
никаких дополнительных проблем не составили. Из данных таблицы 8 следует,
что в качестве подлежащего генетическому анализу эмпирического отношения
расщепления должно подлежать соотношение 242 матовых: 86 глянцевых.
Таблица 8 Результаты сравнения эмпирических отношений расщепления в
тринадцати вариантах эксперимента, подлежащих генетическому анализу
признака опушение завязи
Эмпирическое распределение в различных
Сумма
Теоретически определяемые
семьях
по
классам
∑
Поправочные коэффициенты
χ 2= < χ 2
05=
р>0,05
Н0 принимается
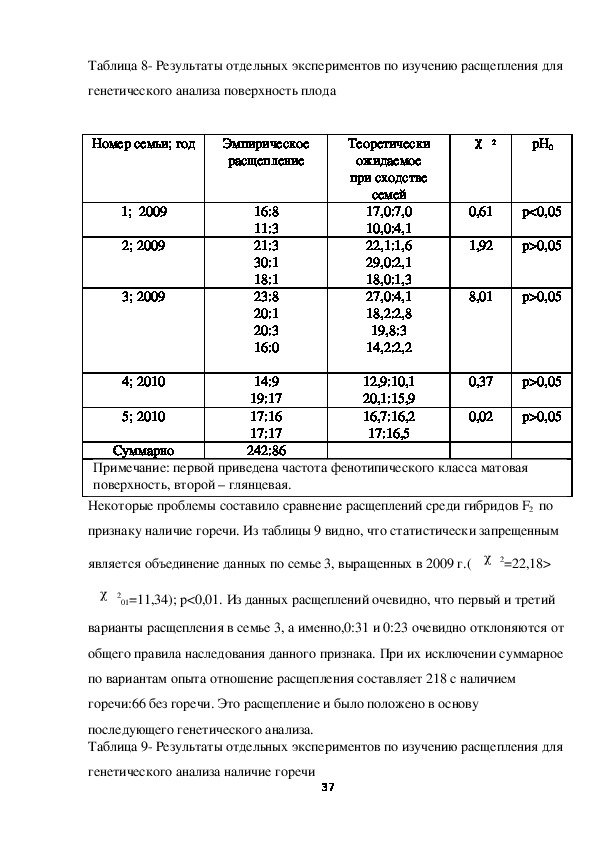
36Таблица 8 Результаты отдельных экспериментов по изучению расщепления для
генетического анализа поверхность плода
Номер семьи; год
Эмпирическое
расщепление
Теоретически
ожидаемое
при сходстве
1; 2009
2; 2009
3; 2009
4; 2010
5; 2010
Суммарно
16:8
11:3
21:3
30:1
18:1
23:8
20:1
20:3
16:0
14:9
19:17
17:16
17:17
242:86
семей
17,0:7,0
10,0:4,1
22,1:1,6
29,0:2,1
18,0:1,3
27,0:4,1
18,2:2,8
19,8:3
14,2:2,2
12,9:10,1
20,1:15,9
16,7:16,2
17:16,5
χ 2
pH0
0,61
1,92
р<0,05
р>0,05
8,01
р>0,05
0,37
0,02
р>0,05
р>0,05
Примечание: первой приведена частота фенотипического класса матовая
поверхность, второй – глянцевая.
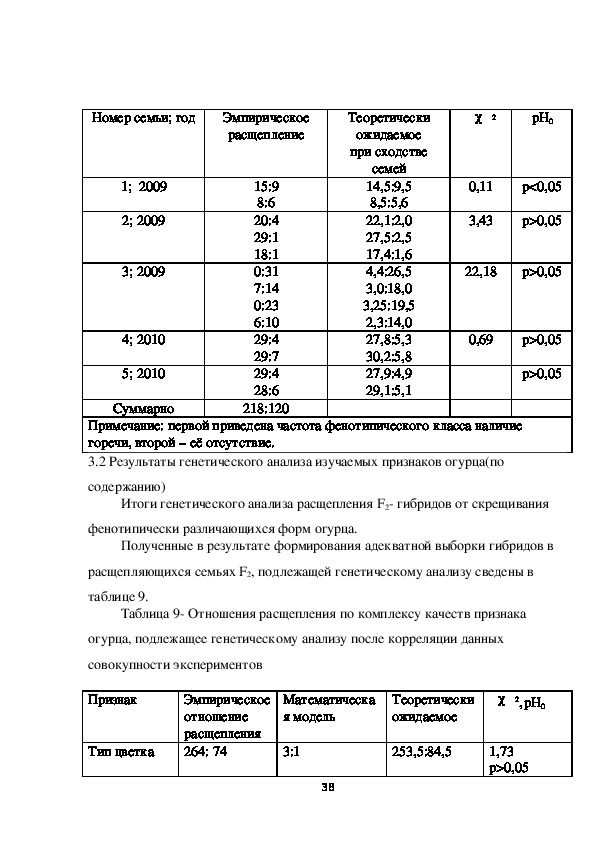
Некоторые проблемы составило сравнение расщеплений среди гибридов F2 по
признаку наличие горечи. Из таблицы 9 видно, что статистически запрещенным
является объединение данных по семье 3, выращенных в 2009 г.( χ 2=22,18>
χ 2
01=11,34); р<0,01. Из данных расщеплений очевидно, что первый и третий
варианты расщепления в семье 3, а именно,0:31 и 0:23 очевидно отклоняются от
общего правила наследования данного признака. При их исключении суммарное
по вариантам опыта отношение расщепления составляет 218 с наличием
горечи:66 без горечи. Это расщепление и было положено в основу
последующего генетического анализа.
Таблица 9 Результаты отдельных экспериментов по изучению расщепления для
генетического анализа наличие горечи
37Номер семьи; год
Эмпирическое
расщепление
Теоретически
ожидаемое
при сходстве
1; 2009
2; 2009
3; 2009
4; 2010
5; 2010
15:9
8:6
20:4
29:1
18:1
0:31
7:14
0:23
6:10
29:4
29:7
29:4
28:6
семей
14,5:9,5
8,5:5,6
22,1:2,0
27,5:2,5
17,4:1,6
4,4:26,5
3,0:18,0
3,25:19,5
2,3:14,0
27,8:5,3
30,2:5,8
27,9:4,9
29,1:5,1
χ 2
pH0
0,11
3,43
р<0,05
р>0,05
22,18
р>0,05
0,69
р>0,05
р>0,05
Суммарно
218:120
Примечание: первой приведена частота фенотипического класса наличие
горечи, второй – её отсутствие.
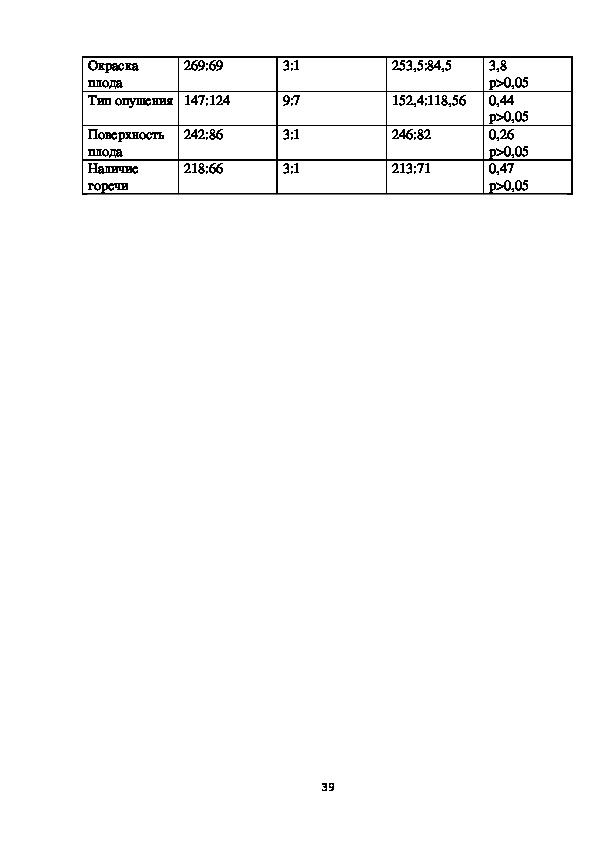
3.2 Результаты генетического анализа изучаемых признаков огурца(по
содержанию)
Итоги генетического анализа расщепления F2 гибридов от скрещивания
фенотипически различающихся форм огурца.
Полученные в результате формирования адекватной выборки гибридов в
расщепляющихся семьях F2, подлежащей генетическому анализу сведены в
таблице 9.
Таблица 9 Отношения расщепления по комплексу качеств признака
огурца, подлежащее генетическому анализу после корреляции данных
совокупности экспериментов
Признак
Тип цветка
Эмпирическое
отношение
расщепления
264: 74
Математическа
я модель
Теоретически
ожидаемое
χ 2, pH0
3:1
253,5:84,5
38
1,73
р>0,05269:69
Окраска
плода
Тип опушения 147:124
Поверхность
плода
Наличие
горечи
242:86
218:66
3:1
9:7
3:1
3:1
253,5:84,5
152,4:118,56
246:82
213:71
3,8
р>0,05
0,44
р>0,05
0,26
р>0,05
0,47
р>0,05
39Таблица 4 Результаты сравнения эмпирических отношений расщепления в
тринадцати вариантах эксперимента, подлежащих генетическому анализу
признака окраска плода огурца
Эмпирическое распределение в различных
семьях
Сумма
по
классам
Теоретически определяемые
∑
Поправочные коэффициенты
χ 2= < χ 2
05=
р>0,05
Н0 принимается
40