Поделиться

Хромосомная теория наследственности. Сцепленное наследование генов
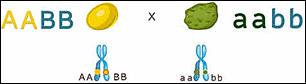 В начале 20-го века учёные генетики стали проводить
множество экспериментов по скрещиванию на самых различных объектах. В итоге
обнаружилось, что закономерности, установленные Менделем проявляются не всегда.
В начале 20-го века учёные генетики стали проводить
множество экспериментов по скрещиванию на самых различных объектах. В итоге
обнаружилось, что закономерности, установленные Менделем проявляются не всегда.
Мендель скрещивал дигетерозиготы – организмы которые отличались по двум признакам. Признаки, которые рассматривал Мендель были локализованы в разных гомологичных хромосомах.
Вспомним что третий закон Менделя формулируется так: каждая пара аллельных генов (и альтернативных признаков, контролируемых ими) наследуется независимо друг от друга.
Что значит независимо друг он друга?
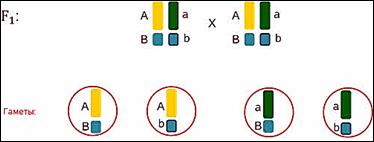 При скрещивании организмов 1-го поколения при мейозе
образуются 4 типа гамет.
При скрещивании организмов 1-го поколения при мейозе
образуются 4 типа гамет.
Где гены комбинируются в различных сочетаниях. Такие сочетания получились потому что гены находились в разных хромосомах.
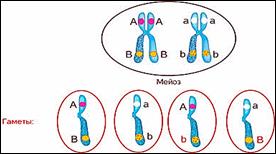 Но в 1906 году Уильям Бэтсон и Риджинальд
Пэннет, проводя скрещивание растений душистого горошка и анализируя
наследование нескольких признаков формы пыльцы и окраски цветков, обнаружили,
что эти признаки не дают независимого распределения в потомстве в соотношении
3:1, гибриды всегда повторяли признаки родительских форм.
Но в 1906 году Уильям Бэтсон и Риджинальд
Пэннет, проводя скрещивание растений душистого горошка и анализируя
наследование нескольких признаков формы пыльцы и окраски цветков, обнаружили,
что эти признаки не дают независимого распределения в потомстве в соотношении
3:1, гибриды всегда повторяли признаки родительских форм.
Стало ясно, что не для всех признаков характерно независимое распределение в потомстве и свободное комбинирование.
Дело в том, что ген формы пыльцы и ген окраски цветка лежат в одной хромосоме.
Признаков в организме значительно больше чем хромосом, в которых эти признаки локализованы. Следовательно, каждая хромосома несёт не один ген, а целую группу генов, отвечающих за развитие разных признаков.
Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался Томас Морган.
Он предложил закон сцепленного наследования (закон Моргана): гены, которые находиться в одной хромосоме, при мейозе попадают в одну гамету, то есть наследуются сцеплено.
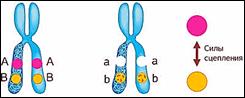 Что значит сцеплено?
Что значит сцеплено?
То есть между генами, которые находиться в одной хромосоме возникают силы сцепления, то есть силы взаимодействия. И чем ближе эти гены, тем сильнее взаимодействие.
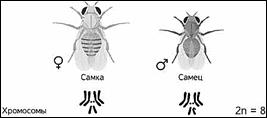
Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала фруктовая мушка дрозофила, которая имела диплоидный набор из 8 хромосом.
Небольшие размеры, короткий жизненный цикл и простота культивирования позволяет использовать ряд видов дрозофил как образцовые объекты генетических исследований.
Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее.
Дрозофила фруктовая — наиболее важный для научных исследований вид дрозофил. Главными её характеристиками как модельного объекта является малое число хромосом. Дрозофила каждые две недели при температуре 25 °С достаточно легко размножаются в пробирках и даёт многочисленное потомство.
 Рассмотрим один из первых экспериментов Томаса Моргана
по изучению сцепленного наследования.
Рассмотрим один из первых экспериментов Томаса Моргана
по изучению сцепленного наследования.
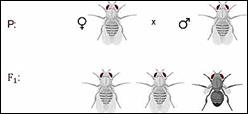 Скрещивая мушку дрозофилу с серым телом и нормальными
крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом
поколении Морган получал гибриды, которые имели серое тело и нормальные крылья.
Ген А-большое отвечает за серое тело, рецессивный ген а-малое за чёрную окраску
тела, доминантный ген В-большое за развитие длинных крыльев, а рецессивный ген
бэ-малое за не развитие крыльев, то есть крылья остаются в зачаточном
состоянии. Значит ген, определяющий серую окраску брюшка, доминирует над темной
окраской, а ген, обуславливающий развитие нормальных крыльев, — над геном
недоразвитых. Далее при скрещивании двух гетерозигот которые имеют серое тело и
длинные крылья в первом поколении получается два фенотипических класса
организмов. Которые имеют серое тело и длинные крылья и чёрное тело с
зачаточными крыльями.
Скрещивая мушку дрозофилу с серым телом и нормальными
крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом
поколении Морган получал гибриды, которые имели серое тело и нормальные крылья.
Ген А-большое отвечает за серое тело, рецессивный ген а-малое за чёрную окраску
тела, доминантный ген В-большое за развитие длинных крыльев, а рецессивный ген
бэ-малое за не развитие крыльев, то есть крылья остаются в зачаточном
состоянии. Значит ген, определяющий серую окраску брюшка, доминирует над темной
окраской, а ген, обуславливающий развитие нормальных крыльев, — над геном
недоразвитых. Далее при скрещивании двух гетерозигот которые имеют серое тело и
длинные крылья в первом поколении получается два фенотипических класса
организмов. Которые имеют серое тело и длинные крылья и чёрное тело с
зачаточными крыльями.
То есть здесь расщепление идёт именно на 2 класса, а не на четыре как при дигибридном скрещивании Менделя.
Почему два? Дело в том, что гены окраски тела и длинны крыльев сцеплены в хромосоме.
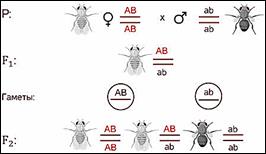 Символы АB АB ab аb располагаются не рядом как мы
записывали их ранее, а как бы друг под другом с двумя чёрточками. Чёрточками мы
условно обозначаем хромосомы.
Символы АB АB ab аb располагаются не рядом как мы
записывали их ранее, а как бы друг под другом с двумя чёрточками. Чёрточками мы
условно обозначаем хромосомы.
В первом поколении организм гетерозиготен по обоим генам, но при образовании гамет эта гетерозиготность не даёт всех возможных комбинаций. То есть родительские гены остаются связанные между собой и гаметы получаются 2х типов.
При комбинации такого рода гамет в потомстве возникают всего 3 генотипических класса потомков.
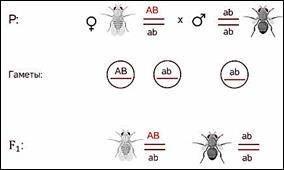 Морган исследуя наследование сцепленных генов
обнаружил что существует нарушение этого правила по дигибридному скрещиванию
Менделя.
Морган исследуя наследование сцепленных генов
обнаружил что существует нарушение этого правила по дигибридному скрещиванию
Менделя.
Он провёл анализирующее скрещивание.
Взял дигетерозиготную особь, которая получилась при скрещивании в первом поколении и скрестил её с чёрной мушкой с зачаточными крыльями, то есть оба рецессивных признака. У него получился необычный результат.
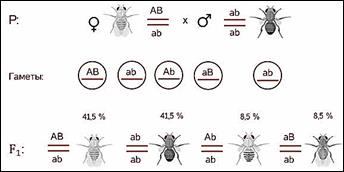 Морган рассуждал. Если
гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном
скрещивании должны были получиться две группы особей, повторяющие признаки
родительских форм, так как материнский организм должен образовывать гаметы
только двух типов — АB и аb, а отцовский — один тип — аb. Следовательно, в
потомстве должны образовываться две группы особей, имеющих генотип АB аb и аa
BB.
Морган рассуждал. Если
гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном
скрещивании должны были получиться две группы особей, повторяющие признаки
родительских форм, так как материнский организм должен образовывать гаметы
только двух типов — АB и аb, а отцовский — один тип — аb. Следовательно, в
потомстве должны образовываться две группы особей, имеющих генотип АB аb и аa
BB.
Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Аa bb и aa Bb.
В потомстве явно преобладали особи с признаками родительских форм (41,5% были серые длиннокрылые и 41,5% — черные с зачаточными крыльями), и лишь незначительная часть мушек имела иное, чем у родителей, сочетание признаков (8,5% были серые с зачаточными крыльями и 8,5% — черные длиннокрылые).
Такие результаты могли быть получены только в том случае, если гены, отвечающие за окраску тела и форму крыльев, находятся в одной хромосоме.
В профазе первого мейотического деления гомологичные хромосомы (то есть идентичные хромосомы одной пары) конъюгируют (сближаются), и могут разрываться в месте контакта в этот момент между ними может произойти обмен участками – кроссинговер.
Кроссинго́вер или перекрёст — это процесс обмена участками гомологичных хромосом во время конъюгации в профазе первого мейоза. В результате образуются крассоверные гаметы.
Организмы, которые возникают в результате слияния кроссоверных гамет называются рекомбинантными.
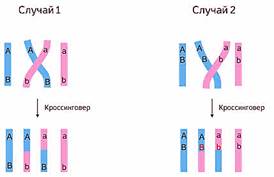 Так в результате, кроссинговера в некоторых
клетках происходит обмен участками хромосом между генами А и В, появляются
гаметы Аb и аB, и, как следствие, в потомстве образуются четыре группы
фенотипов, как при свободном комбинировании генов.
Так в результате, кроссинговера в некоторых
клетках происходит обмен участками хромосом между генами А и В, появляются
гаметы Аb и аB, и, как следствие, в потомстве образуются четыре группы
фенотипов, как при свободном комбинировании генов.
Однако кроссинговер происходит не после каждой конъюгации (сближения хромосом). И определить в каких участках хромосом он произойдет достаточно сложно.
В ходе эксперимента Томасу Моргану удалось доказать, что частота кроссинговера между генами прямо пропорциональна расстоянию между ними в хромосоме. То есть можно сказать что, чем дальше гены находятся друг от друга в хромосоме, тем чаще между ними происходит кроссинговер.
Если рассматривать 2 гена А и В мы можем увидеть 2 случая.
В первом случае гены А и В находиться по разные стороны перекрёста. Тогда после прохождения кроссинговера мы увидим новые сочетания аллелей этих двух генов. В данном случае Аb и аB.
Во 2м случае гены А и B находиться по одну сторону от перекрёста. Тогда после прохождения кроссинговера новых сочетаний аллелей данных двух генов мы не увидим.
Таким образом существуют понятия полного и неполного сцепления.
Неполное сцепление — это разновидность сцепленного наследования, при котором гены анализируемых признаков располагаются на некотором расстоянии друг от друга, что делает возможным кроссинговер между ними.
Полное сцепление — это разновидность сцепленного наследования, при котором гены анализируемых признаков располагаются так близко друг к другу, что кроссинговер между ними становится невозможным.
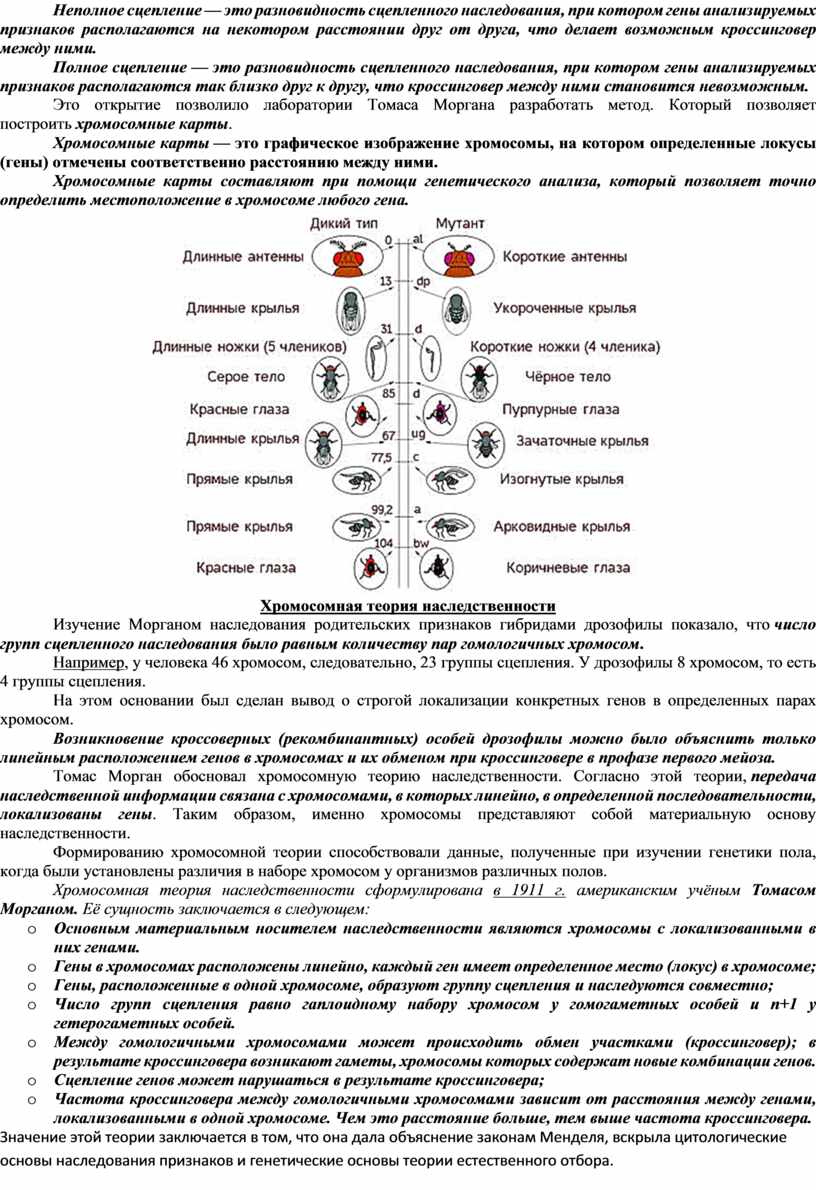
Это открытие позволило лаборатории Томаса Моргана разработать метод. Который позволяет построить хромосомные карты.
Хромосомные карты — это графическое изображение хромосомы, на котором определенные локусы (гены) отмечены соответственно расстоянию между ними.
Хромосомные карты составляют при помощи генетического анализа, который позволяет точно определить местоположение в хромосоме любого гена.
Хромосомная теория наследственности
Изучение Морганом наследования родительских признаков гибридами дрозофилы показало, что число групп сцепленного наследования было равным количеству пар гомологичных хромосом.
Например, у человека 46 хромосом, следовательно, 23 группы сцепления. У дрозофилы 8 хромосом, то есть 4 группы сцепления.
На этом основании был сделан вывод о строгой локализации конкретных генов в определенных парах хромосом.
Возникновение кроссоверных (рекомбинантных) особей дрозофилы можно было объяснить только линейным расположением генов в хромосомах и их обменом при кроссинговере в профазе первого мейоза.
Томас Морган обосновал хромосомную теорию наследственности. Согласно этой теории, передача наследственной информации связана с хромосомами, в которых линейно, в определенной последовательности, локализованы гены. Таким образом, именно хромосомы представляют собой материальную основу наследственности.
Формированию хромосомной теории способствовали данные, полученные при изучении генетики пола, когда были установлены различия в наборе хромосом у организмов различных полов.
Хромосомная теория наследственности сформулирована в 1911 г. американским учёным Томасом Морганом. Её сущность заключается в следующем:
o Основным материальным носителем наследственности являются хромосомы с локализованными в них генами.
o Гены в хромосомах расположены линейно, каждый ген имеет определенное место (локус) в хромосоме;
o Гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются совместно;
o Число групп сцепления равно гаплоидному набору хромосом у гомогаметных особей и n+1 у гетерогаметных особей.
o Между гомологичными хромосомами может происходить обмен участками (кроссинговер); в результате кроссинговера возникают гаметы, хромосомы которых содержат новые комбинации генов.
o Сцепление генов может нарушаться в результате кроссинговера;
o Частота кроссинговера между гомологичными хромосомами зависит от расстояния между генами, локализованными в одной хромосоме. Чем это расстояние больше, тем выше частота кроссинговера.
Значение этой теории заключается в том, что она дала объяснение законам Менделя, вскрыла цитологические основы наследования признаков и генетические основы теории естественного отбора.

Материалы на данной страницы взяты из открытых источников либо размещены пользователем в соответствии с договором-офертой сайта. Вы можете сообщить о нарушении.
