Поделиться

Факторы биологической эволюции.
Факторы эволюции — движущие силы, вызывающие и закрепляющие изменения в генетической структуре популяций.
Популяция — группа особей одного вида, занимающих определенную территорию, свободно скрещивающихся между собой и относительно изолированных от других представителей данного вида. Популяция — элементарная единица эволюции.
Вид — совокупность особей, обладающих сходными морфологическими и физиологическими признаками, занимающих определенную территорию обитания (ареал) и одинаковую экологическую нишу, способных свободно скрещиваться между собой и давать плодовитое потомство.
Микроэволюция — процессы эволюции, идущие внутри вида (на уровне популяций). Их результатом может стать видообразование.
Основные движущие силы эволюции по Ч. Дарвину:
наследственная изменчивость; борьба за существование; естественный отбор.
Естественный отбор задает направление эволюционного процесса.
Согласно основному закону популяционной генетики — закону Харди-Вайнберга — в популяции при стабильных условиях окружающей среды частоты аллелей генов будут оставаться постоянными из поколения в поколение. Генетика и эволюционная теория не сразу нашли «общий язык». Так, одним из основных возражений теории Дарвина среди его современников был так называемый «кошмар Дженкина» (по имени автора возражения, английского инженера). Согласно ему, случайно появившийся у отдельной особи в результате мутации полезный признак в группе организмов (популяции) постепенно будет «размыт» скрещиванием с обычными особями. Это логическое затруднение преодолено с созданием популяционной генетики.
Ошибка Дженкина заключалась в том, что признаки, закрепляемые отбором, согласно законам генетики, не исчезают при скрещивании, а передаются в полном объеме («размывающий» эффект скрещивания просто не существует). Даже в случае полного доминирования рецессивный ген сохраняется и передается потомству. Но о законах генетики ни Дарвин, ни Дженкин еще ничего не знали.
Только в начале XX в. были сформулированы основные принципы генетики популяций. Главное ее положение — закон Харди–Вайнберга — гласит, что в идеальной бесконечно большой популяции, все скрещивания в которой случайны (принцип панмиксии) и в которой не действуют никакие факторы эволюции, частоты всех аллелей остаются постоянными в ряду поколений. Это положение легко проверить, пользуясь элементарной математикой. Доминантный аллель не вытесняет рецессивный, в противоположность убеждению многих не знакомых с этим принципом людей. Если в идеальной популяции существует ген альбинизма с частотой 0,01 %, то с той же самой частотой этот аллель будет появляться в каждом из последующих поколений.
Закон Харди–Вайнберга
Закон Харди–Вайнберга — основа математических построений генетики популяций и современной эволюционной теории. Сформулирован независимо друг от друга математиком Г. Харди (Англия) и врачом В. Вайнбергом (Германия) в 1908 г. Этот закон утверждает, что частоты аллелей и генотипов в данной популяции будут оставаться постоянными из поколения в поколение при выполнении следующих условий:
• численность особей популяции достаточно велика (в идеале бесконечно велика);
• спаривание происходит случайным образом (т.е. осуществляется панмиксия); мутационный процесс отсутствует;
• отсутствует обмен генами с другими популяциями;
• естественный отбор отсутствует, т. е. особи с разными генотипами одинаково плодовиты и жизнеспособны.
Иногда этот закон формулируют иначе: в идеальной популяции частоты аллелей и генотипов постоянны. (Поскольку описанные выше условия выполнения данного закона и есть свойства идеальной популяции.) Математическая модель закона отвечает формуле:
p2+2pq+q2=1.
Она выводится на основе следующих рассуждений. В качестве примера возьмем простейший случай — распределение двух аллелей одного гена. Пусть два организма являются основателями новой популяции. Один из них является доминантной гомозиготой (АА), а другой — рецессивной гомозиготой (аа). Естественно, что все их потомство в F1 будет единообразным и будет иметь генотип (Аа). Далее особи F1 будут скрещиваться между собой. Обозначим частоту встречаемости доминантного аллеля (А) буквой p, а рецессивного аллеля (а) — буквой q. Поскольку ген представлен всего двумя аллелями, то сумма их частот равна единице, т. е. р + q = 1. Рассмотрим все яйцеклетки в данной популяции. Доля яйцеклеток, несущих доминантный аллель (А), будет соответствовать частоте этого аллеля в популяции и, следовательно, будет составлять р. Доля яйцеклеток, несущих рецессивный аллель (а), будет соответствовать его частоте и составлять q. Проведя аналогичные рассуждения для всех сперматозоидов популяции, придем к заключению о том, что доля сперматозоидов, несущих аллель (А), будет составлять р, а несущих рецессивный аллель (а) — q. Теперь составим решетку Пеннета, при этом при написании типов гамет будем учитывать не только геномы этих гамет, но и частоты несомых ими аллелей. На пересечении строк и столбцов решетки мы получим генотипы потомков с коэффициентами, соответствующими частотам встречаемости этих генотипов.
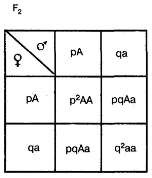
Из приведенной решетки видно, что в F2 частота доминантных гомозигот (АА) составляет р, частота гетерозигот (Аа) — 2pq, а рецессивных гомозигот (аа) — q. Поскольку приведенные генотипы представляют собой все возможные варианты генотипов для рассматриваемого нами случая, то сумма их частот должна равняться единице, т. е.
p2+2pq+q2=1.
Главное применение закона Харди–Вайнберга в генетике природных популяций — вычисление частот аллелей и генотипов. Рассмотрим пример использования этого закона в генетических расчетах. Известно, что один человек из 10 тыс. является альбиносом, при этом признак альбинизма у человека определяется одним рецессивным геном. Давайте вычислим, какова доля скрытых носителей этого признака в человеческой популяции. Если один человек из 10 тыс. является альбиносом, то это значит, что частота рецессивных гомозигот составляет 0,0001, т. е. q2 = 0,0001. Зная это, можно определить частоту аллеля альбинизма q, частоту доминантного аллеля нормальной пигментации р и частоту гетерозиготного генотипа (2pq). Люди с таким генотипом как раз и будут скрытыми носителями альбинизма, несмотря на то что фенотипически этот ген не будет у них проявляться и они будут иметь нормальную пигментацию кожи.
Из приведенных простых расчетов видно, что, хотя число альбиносов крайне невелико — всего лишь один человек на 10 тыс., ген альбинизма несет значительное количество людей — около 2 % . Иными словами, даже если признак фенотипически проявляется очень редко, то в популяции присутствует значительное количество носителей этого признака, т. е. особей, имеющих этот ген в гетерозиготе.
Благодаря открытию закона Харди–Вайнберга процесс микроэволюции стал доступен непосредственному изучению: о его ходе можно судить по изменениям из поколения в поколение частот генов (или генотипов). Таким образом, несмотря на то что этот закон действителен для идеальной популяции, которой нет и не может быть в природе, он имеет огромное практическое значение, так как дает возможность рассчитать частоты генов, изменяющиеся под влиянием различных факторов микроэволюции.
В реальных популяциях дело обстоит иначе. Они не бесконечны, скрещивания в них не являются полностью случайными, а также они почти всегда подвергаются действию факторов эволюции, изменяющих частоты аллелей.
Однако многие реальные популяции хорошо описываются законом Харди–Вайнберга. Например, пользуясь им, рассчитывают хорошо согласующиеся с реальными частоты аллелей групп крови, альбинизма и других признаков, по которым в большинстве случаев практически отсутствует отбор у человека. Человеческие популяции весьма велики и достаточно хорошо «перемешиваются». Изменение частот аллелей в популяции является элементарным событием микроэволюции.
К факторам эволюции относятся:
• наследственная изменчивость (мутационная и комбинативная);
• борьба за существование;
• естественный отбор;
• волны жизни;
• изоляция;
• миграция; дрейф генов.
Определение
Мутационная изменчивость — изменение генотипа под действием мутаций (случайных ненаправленных скачкообразных изменений генотипа).
Сами по себе мутации лишь создают материал для естественного отбора. Они не определяют направления эволюции, так как возникают случайно и не имеют приспособительного значения.
Мутации:
• положительные мутации — мутации, полезные в данных условиях;
• отрицательные мутации — мутации, вредные в данных условиях;
• нейтральные мутации — мутации, не влияющие на жизнедеятельность организма в данных условиях.
Но так как условия постоянно изменяются, мы не можем предсказать, какое значение будет иметь данная мутация завтра. Это говорит об относительности характера мутаций.
Комбинативная изменчивость — изменение генотипа в результате комбинации родительских генов в процессе оплодотворения.
Комбинативная изменчивость возникает благодаря гетерозиготности организмов в популяции и проявляется практически всегда. Мутационная является более редкой и часто приводит к появлению вредных признаков, но позволяет организмам приобретать принципиально новые свойства.
Борьба за существование Борьба за существование — это совокупность отношений, существующих между организмами и условиями среды.
Побеждают в борьбе за существование и продолжают род наиболее приспособленные особи, которые могут передать потомкам совокупность «выигрышных» признаков, что способствует сохранению популяции.
Формы борьбы за существование:
• борьба с неблагоприятными условиями среды; наиболее серьезное влияние оказывает в период новорожденности;
• межвидовая борьба за существование — борьба за природные ресурсы: территорию, убежища, пищу, воду, солнечный свет (у растений) и т. п.;
• внутривидовая борьба за существование — борьба за природные ресурсы и полового партнера. По мнению Ч. Дарвина, наиболее жесткая форма борьбы, так как происходит между животными, занимающими одну экологическую нишу, т. е. использующими одинаковые ресурсы.
Естественный отбор — это процесс отбора генотипов особей, наиболее приспособленных к данным условиям среды, и устранения генотипов особей, менее приспособленных к данным условиям.
Более приспособленные к данным условиям среды особи оставляют больше потомков, чем менее приспособленные. Естественный отбор является движущей силой эволюции.
Формы естественного отбора:
• Стабилизирующий отбор — форма естественного отбора, при котором действие направлено против особей, имеющих крайние отклонения от средней нормы, в пользу особей со средней выраженностью признака.
Ведет к сужению нормы реакции (пределов, в которых в зависимости от условий внешней среды может изменяться определенный признак) признака при постоянных условиях окружающей среды.
Понятие стабилизирующего отбора ввел в науку и проанализировал И. И. Шмальгаузен.
Пример:
• сохранение особей со средней плодовитостью. Чем больше птенцов или детенышей в гнезде, тем труднее их выкормить, тем каждый из них меньше и слабее. В результате наиболее приспособленными оказываются особи со средней плодовитостью;
• выживание детенышей со средним весом. У млекопитающих новорожденные с очень низким и очень высоким весом чаще погибают при рождении или в первые недели жизни, чем новорожденные со средним весом;
• сохранение реликтовых видов: латимерии, галапагосских черепах, гаттерии и др.
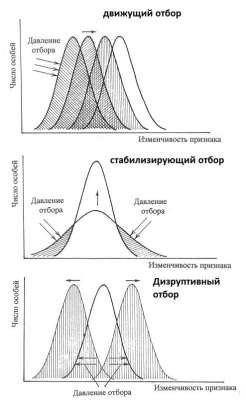
• Движущий отбор — форма естественного отбора, при котором происходит сдвиг нормы реакции признака.
В результате в популяции от поколения к поколению происходит сдвиг нормы реакции признака в определенном направлении. Движущий отбор осуществляется при изменении окружающей среды или приспособлении к новым условиям при расширении ареала. Он сохраняет наследственные изменения в определенном направлении, перемещая соответственно и норму реакции.
Один из немногих достоверных примеров движущего отбора продемонстрировал известный эколог, академик С. С. Шварц (1966 г.). Он показал, что у северного подвида узкочерепной полевки (Stenocranius gregalis), недавно освоившего Северный Урал, количественное распределение ряда признаков оказывается асимметричным. К таким признакам относятся относительная масса печени и сердца, а также абсолютный размер тела. У популяции, живущей в устойчивых внешних условиях, распределение по произвольному количественному признаку, скорее всего, будет симметричным. Если же условия меняются и признак оказывается под давлением отбора, то элиминация (устранение особей) по этому признаку с одной стороны усиливается, с другой — ослабевает. В результате меняется форма нормальной кривой, которая в этом случае с одной стороны круто обрывается, а с другой — дает длинный «хвост».
Примеров действия движущего отбора в природе до сих пор известно очень мало. Большинство из приведенных в учебниках примеров не соответствуют движущему отбору, т. к. в них не наблюдается изменение нормы реакции признака.
Индустриальный меланизм
«Индустриальный меланизм английских бабочек»: резкое повышение доли меланистических (имеющих темную окраску) особей в тех популяциях бабочек, которые обитают в промышленных районах. Из-за промышленного воздействия стволы деревьев значительно потемнели, а также погибли светлые лишайники, из-за чего светлые бабочки стали лучше видны для птиц. В XX в. в ряде районов доля темноокрашенных бабочек достигла 95 %, в то время как впервые темная бабочка была отловлена в 1848 г.

В природе процесс идет медленно: у позвоночных — около полумиллиона лет.
Искусственный отбор является, по сути, ускоренным движущим отбором. Чтобы вывести новую породу кур, требуется 3–6 лет, а чтобы получить расу (форму) насекомых, устойчивых к ядохимикатам, достаточно обработать поле и среди массы погибших насекомых найти единицы выживших. От них и возьмет свое начало новая форма.
Провеивая семена культурных злаков, человек отделял семена сорняка погремка большого, у которых было крыло. В результате изменчивости на полях остались сорняки с бескрылыми семенами, т. к. их отвеять было невозможно, они оставались в семенном материале и размножались в посев.
Дизруптивный (разрывающий) отбор — форма естественного отбора, благоприятствующая двум или нескольким направлениям изменчивости, но не благоприятствующая промежуточному состоянию признака.
При действии дизруптивного отбора внутри популяции возникает полиморфизм — несколько отчетливо различающихся фенотипических форм. При снижении возможности скрещивания между такими популяциями в результате изоляции друг от друга происходит их дальнейшая дивергенция (расхождение признаков), вплоть до обособления в качестве новых видов.
Иногда дизруптивный отбор рассматривают как частный случай движущего отбора, поскольку обе эти формы отбора приводят к изменению фенотипического облика популяций в противоположность стабилизирующему отбору.
Примеры.
Мальки окуней питаются мальками других видов рыб. При отсутствии корма, необходимого для подрастающей молоди окуней, могут сохраниться только «карлики» (особи с резко замедленным ростом, которые длительное время могут питаться планктонными ракообразными) и «гиганты» (особи, способные уже к концу первого года жизни питаться мальками окуней своего же поколения). При такой ситуации в водоеме в течение ряда лет в результате дизруптивного отбора возможно формирование двух рас окуней: «гигантов» и «карликов».
Примером дизруптивного отбора является образование двух рас у погремка большого на сенокосных лугах. В нормальных условиях сроки цветения и созревания семян у этого растения покрывают все лето. Но на сенокосных лугах семена дают преимущественно те растения, которые успевают отцвести и созреть либо до периода покоса, либо цветут в конце лета, после покоса. В результате образуются две расы погремка — ранне- и позднецветущая.
Изоляция — исключение или затруднение свободного скрещивания между особями одного вида.
Изоляция является элементарным эволюционным фактором, действующим на микроэволюционном уровне, и приводит к видообразованию.
Географическая изоляция — пространственное разобщение популяций благодаря особенностям ландшафта в пределах ареала вида (водные преграды для наземных организмов, участки суши для обитателей водоемов, горы, крупные автомагистрали и т. п.).
Ей способствует малоподвижный или неподвижный (у растений) образ жизни.
Экологическая изоляция: разделение по экологическим нишам внутри популяции (особенности окраски покровов, изменение пищевого рациона, размножение в разные сезоны, использование в качестве хозяина организмов разных видов (у паразитов).
Этологическая (поведенческая) изоляция: появление различий в ритуале ухаживания, окраски, запахов, «пения» самцов из разных популяций.
Морфологическая изоляция: различия в структуре органов размножения, разница в размерах тела, препятствующие скрещиванию. У растений такая форма изоляции возникает при приспособлении цветка к определенному виду опылителей.
Описанные формы изоляции, особенно в начальный период их действия, снижают, но не исключают полностью межпопуляционные скрещивания.
Генетическая (репродуктивная) изоляция: несовместимость гамет, гибель зигот непосредственно после оплодотворения, стерильность или малая жизнеспособность гибридов.
Репродуктивная изоляция является непреодолимым барьером для скрещивания.
Волны жизни — колебания численности особей, характерные для любой популяции живых организмов.
Термин ввел С. С. Четвериков (1905 г.).
Их эволюционное значение сводится к случайным изменениям концентрации различных мутаций и генотипов, содержащихся в популяциях, а также к изменению направления и интенсивности отбора. Волны жизни могут быть опасны для выживания малочисленных популяций.
• Сезонные (периодические) волны жизни — генетически обусловленные волны жизни.
• Несезонные (апериодические) волны жизни — обусловленные непосредственным воздействием на популяцию различных абиотических и биотических факторов окружающей среды.
Длина волн жизни прямо пропорциональна продолжительности цикла развития организмов. Часто волны жизни сопровождаются пульсацией границ популяции.
Дрейф генов, или генетико-автоматические процессы — изменение частоты распределения генов из поколения в поколение в силу случайных причин.
Дрейф генов можно наблюдать на примере «эффекта бутылочного горлышка». Если по какой-либо причине численность популяции резко уменьшится (например, в случае необычной засухи или непродолжительного увеличения численности хищников), то результатом будет случайное устранение большого числа индивидуумов. К тому времени, когда популяция вновь будет переживать расцвет, в ней будут гены, характерные для случайно выживших индивидуумов, а вовсе не для исходной популяции.








Материалы на данной страницы взяты из открытых источников либо размещены пользователем в соответствии с договором-офертой сайта. Вы можете сообщить о нарушении.