Поделиться

ФАУНАКАЗАХСТАНА
т.9
ПЕРЕПОНЧАТОКРЫЛЫЕ
(HYMENOPTERA)
В.Л.Казенас
РОЮЩИЕОСЫ (HYMENOPTERA, SPHECIDAE)
Выпуск 1
Алматы-1998
МИНИСТЕРСТВО НАУКИ-АКАДЕМИЯ НАУК
РЕСПУБЛИКИ КАЗАХСТАН
Институт зоологии и генофонда животных
т.9
ПЕРЕПОНЧАТОКРЫЛЫЕ
(HYMENOPTERA)
УДК 595.797 (574)
В.Л.Казенас
Выпуск 1
Общая характеристика семейства.
Алматы-1998
ОТ АВТОРА
Настоящая книга является первой для Казахстана фаунистической сводкой по роющим осам, одному из самых крупных семейств жалящих перепончатокрылых. Помимо определительных таблиц подсемейств, триб, родов и видов Sphecidae, в книге содержатся общие сведения по морфологии, систематике, экологии, зоогеографии и филогении роющих ос, а также характеристики всех таксонов. Определительные таблицы и описания таксонов сопровождаются рисунками таксономических признаков. Для каждого вида приводятся синонимы, морфологическое описание имаго, данные по распространению, экологии и биологии.
Книга предназначена для энтомологов различных направлений, преподавателей, аспирантов и студентов вузов биологического и сельскохозяйственного профиля.
1-й выпуск монографии посвящен подсемействам Ampulicinae и Sphecinae, но содержит также общую характеристику семейства в целом.
Кроме сведений из литературы, автор использовал результаты собственных исследований, а также коллекционные материалы из Зоологического института Российской академии наук, Зоомузея Московского государственного университета и некоторых других научных учреждений. За постоянную помощь в работе автор выражает сердечную благодарность проф. И.Д.Митяеву и проф. В.В.Пулавскому (W.J.Pulawski). За помощь в сборе и обработке материалов и ценные советы автор благодарен также В.И.Тобиасу, П.И.Мариковскому, С.Н.Мярцевой, В.Г.Маршакову, Э.Р.Будрису, К.А.Джанокмен, А.В.Антропову, В.Т.Якушкину, А.С.Лелею, Н.В.Курзенко, П.Г.Немкову, Ш.Д.Исламову,
Ш.Назаровой, Л.Д.Казенасу, Н.Г.Романенко и многим другим. Автор благодарен коллективу и руководству Института зоологии и генофонда животных Министерства науки – Академии наук Республики Казахстан за предоставленную возможность проводить в течение многих лет исследования по теме данной монографии, а также Международному фонду Дж. Сороса, предоставившему грант по проблеме «Биоразнообразие» (1992-1993) и Комитету по грантам Э.Майера Гарвардского университета (США, шт. Массачусетс), выделившему автору грант для работы в Лондонском музее естествознания (NHML) (1997).
1. ВВЕДЕНИЕ
Роющие осы, или сфециды, - крупное семейство (или надсемейство) жалящих перепончатокрылых, насчитывающее около 8000 видов на Земном шаре. Как энтомофаги роющие осы выполняют в природе важную роль, поскольку участвуют в регуляции численности многих видов насекомых (и пауков) и зачастую заметно снижают численность вредителей сельского и лесного хозяйства или насекомых, имеющих отрицательное ветеринарное или медицинское значение. Лишь немногие виды роющих ос наносят вред, уничтожая полезных насекомых и пауков: опылителей или энтомофагов. Несколько видов вредят пчеловодству.
Благодаря сложности и многообразию поведенческих реакций роющие осы с давних пор служат модельными объектами для зоопсихологических и этологических исследований, причем именно благодаря наблюдениям за сфецидами сделаны многие важные открытия в этих областях науки.
Немаловажна роль роющих ос в опылении многих цветковых растений. Яд некоторых видов роющих ос используется в физиологии и биохимии как инструмент тонких нейрофизиологических и биохимических исследований.
Благодаря своеобразному диплоидно-гаплоидному регулированию пола, роющие осы, как и другие перепончатокрылые, представляют собой интерес для исследований в области геномной структуры популяций. Разнообразием внутривидовых отношений сфециды могут помочь в решении вопросов происхождения и развития социальности у насекомых.
Наконец, изучение роющих ос как одной из крупных процветающих групп насекомых дает и может дать в будущем немало ценных сведений для разрешения общих проблем в зоогеографии, экологии и эволюционной теории.
Изучение роющих ос Казахстана имеет длительную, почти полуторавековую историю. Самой первой работой, касающейся фауны сфецид Казахстана, является статья Э.А.Эверсмана (Eversmann, 1849) о перепончатокрылых Волго-Уральского междуречья. Большинство видов указывается в ней для Поволжья, но более 10 видов – для Западного Казахстана («Киргизские степи»). Также для этого региона приведено 7 видов в работе А.Беккера (Becker, 1866), совершившего путешествие по Северному Прикаспию.
Начало изучения фауны и систематики роющих ос в Южном Казахстане и фаунистически тесно связанной с ним Средней Азии связано главным образом с именами Ф.Ф.Моравица и О.И.Радошковского. О.И.Радошковский, один из создателей и первых президентов Русского
энтомологического общества, опубликовал около 74 работ по систематике и биологии насекомых, в том числе перепончатокрылых, в частности роющих ос. Он одним из первых отечественных гименоптерологов обратил внимание на глубокое своеобразие фауны перепончатокрылых Казахстана и Средней Азии («Туркестана» и «Закаспия»). В 1876-м году им были описаны из Туркестана новый род и новый вид Pseudoscolia maculata (Radoszkowski, 1876). Большой интерес в научных кругах вызвала его работа о перепончатокрылых, собранных в Туркестане экспедицией А.П.Федченко в 1869-1871 гг. (Radoszkowski, 1877). Для территории северо-восточного Узбекистана (нынешних Самаркандской, Ташкентской, Ферганской и Андижанской областей) и прилегающих районов Таджикистана, Кыргызстана и Казахстана в ней приведено 144 вида сфецид, из которых 70 видов и 3 рода были описаны как новые. В последующем О.И.Радошковский неоднократно обращался к материалам по перепончатокрылым из Средней Азии и Южного Казахстана, причем особое внимание уделял закаспийским видам. Он опубликовал несколько работ о перепончатокрылых Закаспия, включающих около 110 видов сфецид (Radoszkowski, 1886, 1887, 1888, 1893a, 1893b). В этих работах приведены описания 32 новых видов. Наконец, еще по 1 новому виду было описано О.И.Радошковским (Radoszkowski, 1888) из Казахстана (Рын-пески) и Узбекистана (Ходжент и Ташкент).
Ф.Ф.Моравиц всю свою жизнь посвятил изучению фауны и систематики перепончатокрылых России. Он написал ряд работ по акулеатам Казахстана и Средней Азии. Материалом ему служили в основном сборы выдающихся отечественных путешественников: А.П.Федченко, Н.М.Пржевальского, Г.Н.Потанина, а также В.А.Варенцова, Д.К.Глазунова, Н.М.Мартьянова, В.А.Плющевского-Плющика и др. Ф.Ф.Моравиц описал 89 новых видов из Туркмении, Таджикистана, Узбекистана, Западного и Восточного Казахстана (Morawitz, 1888, 1890, 1891, 1892, 1893a, 1893b, 1894, 1896). Им было опубликовано несколько общих фаунистических списков акулеат: в 1891 г. - Нижнего Поволжья и Западного Казахстана (77 видов сфецид), в 1893 г. – Узбекистана и Таджикистана («Туркестана») (74 вида), в 1894 и 1896 гг. – Туркменистана (49 видов сфецид).
Фауна роющих ос Казахстана и Средней Азии привлекала внимание и некоторых зарубежных энтомологов конца XIX века, особенно австрийских. В своих монографиях о палеарктических видах рода Cerceris А.Шлеттерер (Schletterer, 1887, 1889a, 1889b) для Средней Азии и Казахстана указывает 18 видов и дает для них собственные описания или оригинальные первоначальные описания других авторов. Большинство видов приведены им на основе работ О.И.Радошковского, но несколько видов описаны впервые как новые для науки. А.Гандлирш (Handlirsch, 1887-1893) для этого региона указывает около 50 видов сфецид, но тоже в большинстве случаев ссылаясь на О.И.Радошковского. В 1889 г. совместно с Ф.Колем (Kohl) он опубликовал работу о закаспийских перепончатокрылых, в которой речь идет о 52 видах, в том числе 5 новых (3 из них описаны А.Гандлиршом, 2 – Ф.Колем (Kohl, Handlirsch, 1889).
Все виды, описанные О.И.Радошковским, Ф.Ф.Моравицом, А.Шлеттерером и А.Гандлиршом, были включены в каталог перепончатокрылых Далла Торре (Dalla Torre, 1897). В конце XIX-начале XX века ряд крупных монографий по отдельным родам роющих ос опубликовал Ф.Ф.Коль (Kohl, 1885, 1890, 1891, 1896, 1901a, 1906, 1915, 1918, 1923). В них были включены все известные к тому времени виды (около 100) из Казахстана и Средней Азии. Около 10 видов были описаны самим автором как новые. Следует также упомянуть еще 2 небольшие, но важные работы Ф.Ф.Коля, касающиеся фауны сфецид региона. Это статья о новых видах Tachysphex (Kohl, 1901b) и статья о видах Cerceris в коллекции О.И.Радошковского в г. Кракове (Kohl, 1916). В первой даются описания 3 новых видов Tachysphex из Туркменистана, во второй – описания и таксономические заметки для 20 видов Cerceris и 4 вида описываются как новые.
Фауну и систематику роющих ос (особенно рода Cerceris) Казахстана и Средней Азии изучал А.В.Шестаков. В работах 1912-1927 гг. (см. список литературы) им были описаны 23 вида. Специальная его статья (Шестаков, 1918) посвящена фауне ос рода Cerceris Южного Казахстана и Средней Азии («Туркестана»). В ней приведен список зарегистрированных в этом регионе 67 видов, 7 из которых описаны как новые. А.В.Шестаков (1923) описал новый подрод Apiraptrix рода Cerceris, соответствующий группе rybyensis современной классификации. Кроме того, он установил ряд синонимов и дал новые названия некоторым видам, описанным другими авторами.
Наибольший вклад в изучение фауны и систематики роющих ос Средней Азии и Казахстана внес В.В.Гуссаковский. Он выявил более 300 видов, свыше 100 описал как новые для науки. Помимо фаунистических работ, содержащих описания лишь отдельных новых видов, он опубликовал несколько ревизий по отдельным родам сфецид, в которые включил все известные ему среднеазиатские и казахстанские виды. Одной из первых была работа по роду Astata (Гуссаковский, 1927). В ней для Средней Азии и Казахстана указывается 11 видов, в том числе 2 новых для науки. В.В.Гуссаковский опубликовал ревизию палеарктических видов рода Solierella, а также поправки и дополнения к ней (Гуссаковский, 1928а, 1930г). В этих работах описано 10 новых видов для Средней Азии. В 1930-е годы он опубликовал свыше 10 статей, в том числе 6 ревизий, в которых для Казахстана и Средней Азии привел 6 видов Gastrosericus, в том числе 5 новых (Гуссаковский, 1930в), 6 видов Eremiasphecium Kohl = Shestakovia Guss. (все новые) (Гуссаковский, 1930д), 14 видов, в том числе 8 новых подсемейства Sphecinae (Гуссаковский, 1930а), 5 видов Ammoplanus, 3 вида Ammoplanops и 2 вида Spilomena (Гуссаковский, 1931), 12 видов Prosopigastra, в том числе 8 новых (Гуссаковский, 1933б), 8 видов Trypoxylon, в том числе 4 новых (Гуссаковский, 1936), 3 вида Didineis, 3 вида (2 из них новые) Pison, 15 видов Psenini (Гуссаковский, 1937). Наиболее крупная фаунистическая работа В.В.Гуссаковского о сфецидах вышла в 1952 году. В ней описывается 4 новых рода и 60 новых видов, даются таксономические заметки еще о 8 видах (Гуссаковский, 1952).
В своих работах В.В.Гуссаковский описал несколько новых родов, однако последующими авторами большинство из них были сведены в синонимы (Beaumont, 1955 и др.).
Для познания фауны роющих ос Казахстана и Средней Азии имеют большое значение фаунистические работы зарубежных авторов, в частности Я.Бомона по Афганистану, Ирану и другим соседним странам (Beaumont, 1957, 1960a, 1961, 1963, 1969, 1970 и др.) или К.Цунеки (Tsuneki, 1971a, 1971b, 1971c, 1972a, 1972b) по Монголии, в которых содержатся сведения не только о распространении видов, встречающихся в Казахстане и Средней Азии, но и даются их описания и таксономические заметки.
Сведения о среднеазиатских и казахстанских видах имеются также в некоторых монографических работах зарубежных авторов, содержащих ревизии отдельных палеарктических родов: J. de Beaumont (1953) – 2 вида Olgia из Узбекистана и Южного Казахстана; J. de Beaumont (1962) – 3 вида Chlorion из Туркменистана; P. Roth (1963) – 5 видов Palmodes из Средней Азии и Казахстана.
Наибольшую роль среди зарубежных исследователей в изучении сфецид Средней Азии и Казахстана сыграл В.В.Пулавский (Pulawski). В его работах в отношении сфецид исследуемого региона содержатся следующие сведения: описывается новый подвид Astata, даются таксономические заметки о 3 видах этого рода (Pulawski, 1957), указываются 22 вида Tachytes, из них 6 новых для науки и 5 новых синонимов (Pulawski, 1962); описывается новый вид Diploplectron (Pulawski, 1965b), приводится 8 новых синонимов, дается новый статус пяти таксонам (Pulawski, 1965а), указывается 67 видов Tachysphex (из них 15 новых) и 4 новых подвида, дается новый статус 4 таксонам, приводится несколько новых синонимов (Pulawski, 1971); указывается 5 видов Ammatomus, сводятся в синонимы 3 вида, восстанавливается валидность одного видового названия (Pulawski, 1973), приводится несколько новых синонимов среди Larrinae и Astatinae (Pulawski, 1974a), указывается 6 видов Parapiagetia (Pulawski, 1977), приводится 22 вида Prosopigastra, в том числе 5 новых (Pulawski, 1979b), сводятся в синонимы около десяти видовых названий разных Sphecidae (Pulawski, 1979a, 1981, 1983, 1984), указывается 6 видов Gastrosericus (Pulawski, 1995).
Много сделала для познания роющих ос Казахстана и Средней Азии С.Н.Мярцева. Ею опубликовано около 15 статей. Помимо выяснения видового состава сфецид региона, С.Н.Мярцева много внимания уделила изучению биологии и экологии отдельных видов и групп (Мярцева, 1963б – Podalonia ebenina; 1964a – Sphex; 1968a – Bembix; 1968b – Sceliphron; 1969 – Ammophilini; 1976 – Stizus, Bembecinus, Tachysphex, Larra, Palarus), их хищников и паразитов (Мярцева, 1964б, 1972б, Мярцева, Чаракулиев, 1964а, 1964б, 1964в), возможности практического использования ос (Мярцева, 1966), общих экологических и зоогеографических особенностей (Мярцева, 1968а, 1971, 1972а).
Несколько новых видов роющих ос из Казахстана и республик Средней Азии описал В.Г.Маршаков (1973а, 1973б, 1976а, 1976б, 1976г).
Он также опубликовал ряд ревизий в объеме СССР по отдельным родам Crabronini, Pemphredoninae и Philanthinae, в которые включил среднеазиатские и казахстанские виды (Маршаков, 1975а, 1975б, 1976а, 1976в, 1976г, 1979а, 1979б, 1980а, 1980б, 1980в).
В познание роющих ос Казахстана и Средней Азии внесли свой вклад в последние годы А.В.Антропов (Антропов, 1986, 1991, 1994, Казенас, Антропов, 1994а, 1994б) по Crabronini, Э.Р.Будрис (1985, 1988, 1990) по Pemphredoninae, П.Г.Немков (1990, 1992) по Nyssoninae и М.А.Нестеров (1993) по Goritini.
В Казахстане роющие осы до работ автора специально не изучались. Немногочисленные сведения о нахождении того или иного вида в республике содержатся в некоторых работах упомянутых выше авторов. Интересный случай подавления роющими осами вспышки численности атбасарской кобылки в юго-западном Казахстане описывается в работе М.П.Мальковского (1962). Ряд сведений по биологии сфецид содержится в работах П.И.Мариковского (1961, 1963, 1972 и др.).
Автором опубликовано более 70 работ о сфецидах Казахстана и прилегающих регионов, из них часть посвящена биологии отдельных видов (Казенас, 1964, 1965, 1968б, 1969а, 1970б, 1977в, 1985, 1994; Казенас, Джанокмен, 1970, 1974; Kazenas, Alexander, 1993) или отдельным сторонам их биологии (Казенас, 1970в, 1974а, 1974б, 1974и, 1976а, 1976г, 1984б, 1990, 1995; Казенас, Насырова, 1991; Казенас, Тобиас, 1992; Казенас, Есенбекова, 1995), часть – фаунистике и систематике (Казенас, 1968а, 1969б, 1970а, 1970г, 1971, 1972а, 1972б, 1972в, 1973, 1974а, 1974г, 1975а, 1975б, 1975в, 1976б, 1976в, 1977а, 1977в, 1978а, 1978б, 1978в, 1979а, 1979б, 1979в, 1979г, 1980а, 1980б, 1980в, 1980г, 1980д, 1980е, 1980ж, 1983, 1984а, 1984в, 1987, 1988, 1989а, 1989б, 1990, 1991а, 1991б, 1991в, 1992а, 1992б, 1993а, 1993б, 1993в, 1994а, 1994б, 1994в,
1995а, 1996; Исламов, Казенас, 1983; Будрис, Казенас, 1992; Казенас, Антропов, 1994а, 1994б), в которых для Средней Азии и Казахстана указывается более 400 видов, в том числе более 100 новых.
Отрывочные сведения по фауне и систематике сфецид района исследования имеются в работах ряда современных зарубежных сфецидологов (Merisuo, 1972 – 3 новых вида Pemphredon; Dollfuss, 1983 – 4 вида Spilomena; 1995 – 8 видов Pemphredon).
Таким образом, к настоящему времени имеется достаточно много литературы и накоплен довольно обширный коллекционный материал, свидетельствующие о своеобразии и богатстве фауны роющих ос в Казахстане и Средней Азии. В настоящей работе, состоящей из 4 выпусков 9-го тома 30-томной серии «Фауны Казахстана», обобщены и критически осмыслены доступные материалы по сфецидам Республики Казахстан.
Первый выпуск посвящен подсемействам Ampulicinae и Sphecinae, но содержит также общий очерк по морфологии, биологии, систематике, распространению и практическому значению семейства Sphecidae в целом и определительные таблицы подсемейств, триб и родов, составленные главным образом на основе определительных таблиц Р.М.Бохарта и А.С.Менке (Bohart and Menke, 1976).
При составлении определительных таблиц видов, написании родовых характеристик и повидовых очерков были использованы многочисленные литературные источники (Гуссаковский, 1932, 1935, 1948, 1952; Исламов, 1986; Казенас, 1971, 1972, 1974, 1978; Колесников, 1977; Малышев, 1959, 1966; Мярцева, 1963, 1972; Немков, 1986; Пулавский, 1978; Радошковский, 1877; Abrahamsen, 1972; Bajari, 1957; Balthasar, 1972; Beaumont, 1960, 1963, 1964, 1970; Bischoff, 1930; Bohart, Menke, 1976;
Ceballos, 1956; DallaTorre, 1897; Dollfuss, 1991; Friese, 1926; Iwata, 1976; Kohl, Handlirsch, 1889; Lomholdt, 1975-1976; Morawitz, 1890, 1893, 1894, 1896; Noskiewitz, Pulawski, 1960; Oehlke, 1970; Olberg, 1959; Pulawski, 1958, 1964; Schmiedeknecht, 1930; Zavadil, Snoflak, 1948 и многие другие). Перечисленные работы касаются семейства в целом. Использованная литература по отдельным родам и видам приведена в соответствующих породовых и повидовых очерках. Следует особо отметить, что в описаниях надвидовых таксонов использована обширная информация из монографии Р.М.Бохарта и А.С.Менке (1976), которая в нашей стране является большой библиографической редкостью.
В настоящей главе дается обобщенная морфологическая характеристика тела роющих ос и отдельных его частей с учетом варьирования их строения в отдельных таксонах. Описание тела и его частей основывается на литературных данных и результатах морфологического изучения автором таксонов, представленных в Казахстане. Основное внимание уделяется признакам, использующимся в таксономии роющих ос, что отвечает практическим потребностям построения определительных таблиц. Преимагинальные стадии почти не изучались, поэтому в главе дается только самая общая и очень краткая их морфологическая характеристика. Терминология принята главным образом по Р.М.Бохарту и А.С.Менке (Bohart and Menke, 1976) и В.И.Тобиасу (1978).
Общая форма, размеры, опушение, окраска и скульптура тела имаго. Тело роющих ос имеет хорошо развитую голову с большими глазами, 3 теменными глазками, усиками и ротовыми органами, сильно развитую грудь с 3 парами ног и 2 парами крыльев, состоящую из 3 грудных сегментов и слившегося с ними 1-го брюшного, так называемого промежуточного, сегмента, и удлиненного брюшка, подвижно сочлененного с грудью. Габитус некоторых представителей таксонов, имеющихся в фауне Казахстана, показан на рис. 1-36.
Длина тела у роющих ос Казахстана колеблется от 1,2 до 36 мм. Для большинства видов свойственны средние размеры (10-15 мм). Тело относительно длинное у большинства видов Sphecinae, некоторых видов Larra, Stizus, Sphecius, Bembix, Cerceris, Philanthus и Ectemnius и, повидимому, это связано со специализацией в охоте за крупной добычей, запасаемой для потомства в единственном числе или в небольшом количестве экземпляров.
Скульптура тела у роющих ос очень разнообразна. Она состоит из пунктировки, микропунктировки, нежной сетчатой микроячеистой морщинистости, более или менее грубых складок, морщин, гребней, выступов, бугров и т.д. Грубая скульптура, очевидно, связана с необходимостью усиления прочности покровов как каркаса (скелета) насекомого при интенсификации той или иной функции. Грубая скульптура тела присуща некоторым Cercerini, Oxybelini, Crabronini, Nyssonini, Pemphredonini, Palarini и др. Возможно, в ряде случаев она связана с необходимостью защиты от уколов жалом перепончатокрылых, являющихся их добычей (Philanthus, Cerceris, Palarus) или хозяевами (Nyssonini,
Stizoides).
Окраска тела у роющих ос весьма разнообразная: от однотонно черной до пестрой многоцветной. Темная «металлическая» или черная окраска присуща наиболее примитивным осам, в частности, многим Ampulicinae, наиболее примитивным представителям других подсемейств. Появление и усиление желтой и белой окраски отмечается у Crabronini, Nyssoninae, Astatinae и Larini. У Philanthinae светлый рисунок, как правило, хорошо развит. Посветление окраски, несомненно, связано с освоением аридных биотопов. Светлая окраска тела свойственна очень многим видам пустынных районов Казахстана и Средней Азии.
Интересно, что многим видам роющих ос, обитающим в пустынях, свойственна черная окраска тела (например, некоторым видам Sphex, Palmodes, Podalonia, Liris, Tachytes, Tachysphex, Parapiagetia, Prosopigastra и др.). В ряде случаев ее отрицательное значение как фактора перегрева тела снижается, по-видимому, за счет густого прилегающего опушения тела (например, у некоторых видов рода Prionyx) или зеркально блестящих, сглаженных наружных покровов. Однако в некоторых случаях виды имеют облик явных мезофилов (например, Podalonia ebenina, P. atrocyanea, некоторые виды рода Tachysphex). Скорее всего, в данном случае возможность обитания этих видов в пустынях обусловлена биологическими особенностями, заключающимися в сдвиге активности на ранние утренние часы или ранневесенний период года, что подтверждается наблюдениями автора за фенологией активности Podalonia atrocyanea, P. ebenina, Palmodes occitanicus и др.
Признаки, связанные с окраской тела, широко используются в систематике сфецид при дифференциации видов и подвидов. Видовые различия ярче всего проявляются в окраске брюшка.
Голова. Голова у роющих ос гипогнатическая (ортогнатическая), лишь у некоторых представителей подсемейства Pemphredoninae (например, Spilomena) она прогнатическая. В трибе Sceliphronini она удлиненная дорсо-вентрально и полупрогнатическая. Как правило, она имеет круглую или овальную форму, если смотреть спереди, в частности у представителей подсемейств Ampulicinae (рис. 37а), Sphecinae (рис. 37б-и), Astatinae (рис. 38а-г), Crabroninae (рис. 38д-н, 39а,б), Pemphredoninae (рис. 40а-м), Nyssoninae (рис. 41а-м), Philanthinae (рис. 42а-и). Иногда голова имеет почти квадратную или прямоугольную форму, например, у Crabronini (рис. 39в-и).
У большинства сфецид длина головы явно меньше ее ширины, но у Pemphredon и близких родов задняя часть головы – позади глаз – сильно увеличена, отчего гололва в целом становится почти кубообразной (рис. 43а,б). Сходную форму имеет голова у многих представителей трибы Crabronini (рис. 43в). У части видов Lestica голова сзади сильно сужена у самцов (рис. 43г).
Форма головы в некоторых родах значительно варьирует: от круглой до поперечно овальной (см. спереди), например, у Tachysphex и Eremiasphecium, от поперечно прямоугольной до квадратной у Passaloecus и Pemphredon.
Признаки общей формы головы и строения ее передней поверхности весьма изменчивы в родах Trypoxylon, Spilomena, Rhopalum, Crossocerus, Crabro, Ectemnius, Lestica, Astata, Dryudella, Mimesa, Ammoplanus, Ammoplanellus, Gorytes, Philanthus, Cerceris, Tachysphex и др. и используются для диагностики видов.
Темя. Темя – это верхняя поверхность головы от переднего глазка до вершины затылочного киля. Pemphredon – один из немногих родов Pemphredoninae, имеющих сильно развитое темя (рис. 43а,д), у большинства родов оно развито слабее, а у самцов Astata, Dryudella, некоторых видов Prosopigastra почти целиком редуцируется. У Belomicrus на нем иногда развиты бугры.
Глазки. У большинства роющих ос изучаемого региона глазки нормальные, т.е. округлые, но у некоторых представителей Crabroninae и Nyssoninae они более или менее деформированные (рис. 43е-м). У ос трибы Larrini средний глазок часто уменьшенный, а задние более или менее редуцированные и имеют вид эллипса, конуса, изогнутого или прямого узкого клина или узкого прямоугольника, при этом они непрозрачные. У Larra и Liris задние глазки очень маленькие и нечетко ограниченные, у Gastrosericus и Ancistromma задние глазки изогнуто клиновидные или каплевидные, у Tachytes – запятовидные, с длинным «хвостом», у Tachysphex – овальные или запятовидные, но с коротким «хвостом». У Parapiagetia они удлиненно-овальные или сигаровидные, у Prosopigastra – узко каплевидные. У Palarus задние глазки уплощены и овально удлинены, но имеют прозрачную линзу (рис. 43к).
Деформация глазков происходит также у некоторых представителей Nyssoninae. У большинства Nyssoninae глазки нормальные; у Kohlia они слегка уплощенные и вытянуто яйцевидные, а у Bembix средний (передний) глазок редуцирован до узкой дугообразной полоски и не имеет прозрачной линзы, задние глазки имеют вид коротких темных валиков (рис. 43м).
В систематике Crabronini имеет значение взаимное расположение глазков. У Entomognathus, Lindenius, Crabro, Ectemnius и Lestica глазковый треугольник широкий, т.е. его высота явно меньше ширины. У Rhopalum и Crossocerus глазковый треугольние равносторонний, т.е. его боковые стороны почти равны основанию.
Глаза. Глаза у роющих ос хорошо развиты, имеют овальную фоорму и занимают боковые части головы. В систематике роющих ос используются такие признаки, как общая форма глаз, особенно их внутреннего края, характер взаимного расположения внутренних краев глаз, различие размеров фасеток вверху и внизу, наличие или отсутствие волосков на поверхности глаз и некоторые другие.
У большинства сфецид глаза голые. У Nitela они покрыты нежным опушением. Густое опушение разито и у некоторых видов Pison. У Entomognathus глаза покрыты отстоящими волосками. У некоторых видов Lindenius заметны редкие, очень короткие волоски.
Наиболее обычным признаком у сфецид является однообразие глазных фасеток по размерам на всех участках глаз, но у многих представителей трибы Crabronini фасетки внизу, у наличника, явно крупнее, чем вверху, около темени, а у некоторых представителей трибы Larrini, наоборот, крупнее вверху.
Внутренние края глаз у сфецид прямые или дугообразно изогнутые, параллельные или сближающиеся книзу или кверху, иногда расходящиеся от середины книзу и кверху.
У представителей триб Dolichurini, Sphecini внутренние края глаз обычно прямые в нижней половине и изогнутые в верхней, или параллельные, или сближающиеся книзу, или расходящиеся книзу. У представителей трибы Ammophilini глаза обычно сближаются книзу, по крайней мере у самцов. У самок Podalonia, некоторых видов Eremochares и Ammophila они параллельные или почти параллельные.
У Pemphredoninae внутренние края глаз чаще всего параллельные и прямые или слегка сближающиеся кверху. У Ammoplanus и Ammoplanellus они обычно сближаются книзу.
У Astatinae глаза сближаются кверху, у самцов Astata и Dryudella соприкасаются вверху – голоптические (рис. 38в).
У большинства представителей Larrini внутренние края глаз сближаются кверху, изредка параллельные, например, у некоторых видов Miscophus, Parapiagetia и Gastrosericus. У самцов нескольких видов Prosopigastra глаза соприкасаются вверху, у Palarus почти соприкасаются. У Miscophus расположение внутренних краев глаз разное: они или сближаются кверху, или параллельные, или расходятся кверху, или расходятся от середины и кверху, и книзу. У Pison и Trypoxylon внутренние края глаз имеют глубокую вырезку, придающую глазам почковидную форму (рис. 38з).
У Oxybelini внутренние края глаз параллельные или расходящиеся от середины кверху и книзу (рис. 39а,б), у большинства Crabronini они сближаются книзу (рис. 39в-з). Также они сближаются книзу у Entomosericinae (рис. 39л). В подсемействе Nyssoninae взаимное расположение глаз очень разнообразное: они или параллельные, или сближающиеся внизу либо в середине (рис. 41а-м), иногда широко выемчатые. У Philanthinae внутренние края глаз параллельные или более или менее сближаются кверху (рис. 42а-и). У Philanthus на внутренних краях глаз имеется широкая выемка (рис. 42в,г).
Лоб. Лоб у роющих ос занимает большую часть передней поверхности головы, он ограничен наличником, глазами и теменем. Наличие тех или иных образований, как-то: килей, борозд, бугров – и другие модификации лба характерны для тех или иных представителей семейства сфецид и широко используются в систематике. К числу наиболее своеобразных из них относятся U-образный платформообразный выступ над усиковыми ямками у видов рода Dolichurus, срединная выступающая площадка около верхнего края усиковых ямок у самцов Chalybion, срединный лобный киль у Psen, продольный и поперечный лобные кили с расширением-площадкой между усиковыми ямками у Psenulus, неглубокое скапальное углубление у Stigmus, поперечный уплощенный выступ у самцов рода Dryudella, валикообразные вздутия у Larra, Liris и Ancistromma, большой срединный бугор у Prosopigastra, V-образное возвышение с килем у Solierella, кили, иногда образующую фигуру щита или сердца у Trypoxylon, скапальное углубление (иногда с окаймляющими килями и валиками) у Crabronini, орбитальные ямки у Lindenius, Crabro, Lestica и других Crabronini, налобник, или субантеннальный склерит, у Harpactus, Philanthus и Cerceris.
При диагностике видов и родов сфецид имеет значение наличие или отсутствие лобной линии или борозды. В систематике сфецид широко используются признаки опушения и скульптуры лба, имеющие значительные межвидовые различия.
Усики. Усики сфецид состоят, как у всех перепончатокрылых, из 3 отделов: скапуса, педицеллюса и жгутика, или флагеллума. Последний, в свою очередь, делится на членики (флагелломеры), которых насчитывается у самок 10, у самцов 11. У наиболее примитивных сфецид, в частности Dolichurus, Sceliphron, Chalybion и др., усики тонкие и длинные, членики жгутика цилиндрические, по длине значительно превосходящие свою ширину, покрытые равномерным коротким опушением и лишенные каких-либо образований в виде килей, выступов, выемок, площадок и т.п. У наиболее продвинутых представителей развиты различные модификации, что широко используется в таксономии. У Lestica, Ectemnius, некоторых видов Solierella и Trypoxylon число флагелломер у самцов сокращено до 10. Многие самцы Sceliphronini и Sphecini имеют плакоиды – особые маленькие сенсорные поля – на члениках жгутика.
Имеются плакоиды также у самцов Larra и Liris. У всех других сфецид плакоиды отсутствуют.
Строение усиков в целом и отдельных члеников изменчиво во меногих родах сфецид. Членики жгутиков самцов во многих родах имеют тилоиды - линейные складко- или килевидные образования. Они характерны для многих видов Podalonia, Ammophila, Mimumesa, Astata, Diploplectron, многих родов Crabronini, для Entomosericus, Mellinus, Gorytes и других родов.
Жгутик усиков обычно нитевидный, но у некоторых ос, например, родов Mimesa, Ammatomus и Philanthus он булавовидный (рис. 44а-в) или четковидный. Скапус у примитивных сфецид имеет небольшие размеры, сходные с величиной члеников жгутика. У более продвинутых представителей скапус относительно крупный, у Ammoplanops и Ammoplanellus, например, составляет четверть всего усика. Для ряда сфецид свойственна большая или меньшая деформация члеников усиков у самцов. Например, у самцов Dinetus членики жгутика уплощены, усики сильно изогнуты и закручены (рис. 44г). Отдельные деформированные членики характерны для жгутика самцов Prosopigastra (рис. 44д), Pison, многих Crabronini (рис. 44е-к), Bembecinus (рис. 44л), Bembix (рис. 44м), Pseudoscolia (рис. 44н), Psen (рис. 44о), Hoplisoides (рис. 44п), Harpactus (Рис. 44р) и других родов.
Часто особую форму приобретает только последний членик (иногда несколько вершинных члеников усиков). У Trypoxylon последний членик усиков самцов имеет большую изменчивость: как правило, сильно удлинен и расширен (рис. 45а,б). У Plenoculus и Solierella он или удлинен, или, наоборот, сильно укорочен (рис. 45в,г). У Alysson и Didineis он сильно изогнутый (рис. 45д). Особую форму, отличную от форму других члеников, к тому же весьма изменчивую, он имеет обычно также у Entomosericus (рис. 45е), Oryttus (рис.. 45ж), Psammaecius (рис. 45з), Sphecius (рис. 45и), Nysson (рис. 45к), Cerceris (рис. 45л), Pseudoscolia (рис. 45м) и некоторых представителей других родов.
У Argogorytes, Harpactus, Lestiphorus, Hoplisoides, Psammaecius и
Oryttus 4 вершинных членика усиков уплощенные или вдавленные снизу (рис. 45н). У Cerceris последний членик часто изогнут, иногда к вершине сильно сужен (рис. 45л). На жгутике самцов (например, в роде Cerceris) иногда развита особая бахрома отстоящих волосков.
Усики прикрепляются к голове, как правило, в нижней части лба выше наличника, но иногда – в середине лба или, наоборот, в непосредственном контакте с наличником. В одних подсемействах расположение усиковых ямок варьирует в широких пределах, в других более или менее стабильно.
Наличник. Обычно наличник у сфецид имеет вид поперечной. расширенной в середине пластинки. Его поверхность, как правило, более или менее равномерно выпуклая, а передний край дугообразно выступающий. Часто на его поверхности развиты различные морфологические образования: зубцы, кили, выступы, бугры, валики и т.п. Весьма изменчива форма переднего края, на котором развиваются зубцы, выступы, выемки, вырезки, лопасти и т.д. У некоторых представителей триб Larrini, Miscophini, Trypoxylonini, подсемейства Philanthinae наличник разделен на 3 части, отделенные друг от друга прогитбом и выемкой (вырезкой) на переднем крае. У Palarus эти части отделены друг от друга ясными швами-линиями.
Морфологические признаки наличника у роющих ос (форма переднего края, наличие выступов, зубцов, вырезок, лопастей) очень широко используются в систематике.
У самцов Philanthus, Pseudoscolia и Cerceris на переднем крае по бокам развита бахрома волосков, иногда имеющая вид кисточек или пучков (рис. 46а,б,в). Наличник самок Cerceris часто несет срединный выступ различной формы (рис. 46г,д).
Виски и щеки. Величина и форма висков (части головы между глазами и затылком) у сфецид сильно варьирует, иногда виски имеют выступы в виде зубцов (рис. 46е,ж). Промежуток между нижним краем глаз и мандибулярной ямкой (щека, или малярное поле) у большинства роющих ос очень маленький или отсутствует, но он хорошо развит у некоторых представителей подсемейств Astatinae, Pemphredoninae и Philanthinae.
У некоторых видов Rhopalum виски имеют вентральное вдавление. У некоторых видов рода Gastrosericus виски у самок имеют 1-2 вентральных бугра. У самцов некоторых видов рода Lestica виски сильно сжатые, иногда с килем. У некоторых видов Belomicrus на висках развит киль, у Entomognathus – ямчатая борозда, у Lindenius – киль, у Crabro – вдавление.
Мандибулярная впадина. У Dolichurus – одного из наиболее примитивных родов сфецид мандибулярная впадина соединена с ротовой впадиной, т.е. является открытой. Этот признак характерен также для большинства Larrini, для Psenini, Astatinae, Laphyragoginae, Oxybelini и Nyssoninae. У представителей подсемейства Sphecinae, у Pemphredonini, у видов рода Parapiagetia и большинства представителей Crabronini и
Philanthinae мандибулярная впадина закрытая, т.е. отделена от ротовой впадины парамандибулярным выступом гипостомы, который доходит до внутренней поверхности наличника.
Ротовой аппарат. У сфецид состоит из жвал, верхней губы и лабио-максиллярного комплекса. Максиллы роющих ос лишены лацинии, зато сильно развит кардо. В лабиальном отделе развиты лишь ментум, глосса и щупики. У большинства сфецид ротовые органы короткие, но у некоторых представителей они более или менее удлиненные, или же, наоборот, сильно укороченные. У большинства родов в подсемействе Sphecinae они короткие, но у Podalonia и Ammophila длинные. У некоторых видов Tachytes и Tachysphex (Larrini) также они длинные. Удлинение ротовых органов, очевидно, связано с питанием нектаром цветков, имеющих глубоко расположенные нектарники (например, у многих сложноцветных) в условиях нехватки источников питания, например, в аридных регионах. Укорочение ротовых органов происходит, повидимому, вследствие вторичного перехода к питанию падью или нектаром растений, цветки которых имеют открыто расположенные нектарники.
Верхняя губа. Верхняя губа у роющих ос обычно представляет собой короткую выпуклую или почти плоскую пластинку, скрытую под наличником или слегка выступающую из-под него. Строение верхней губы используется в систематике. Передний край ее у сфецид имеет очень изменчивую форму (зубцы, вырезки, ряды щетинок, лопасти). Обычно она имеет форму прямоугольной пластинки, скрытой или лишь слегка выступающей из-под наличника. У Sphecius и Kohlia она явно выступает и выступающая часть составляет от 1/2 до 2/3 ширины губы. У Stizini губа тоже выступает из-под наличника, но выступающая часть все же короче, чем ширина губы. Только у Bembix – одного из наиболее эволюционно продвинутых родов Nyssoninae – выступающая часть верхней губы значительно (в 2 раза и более) длиннее своей ширины у основания.
Функция верхней губы, очевидно, состоит в защите других ротовых органов от внешних механических повреждений и степень ее развития зависит от величины ротовых частей. В ряде случаев форма верхней губы, возможно, связана с особенностями транспортировки добычи.
Жвалы, или мандибулы. Верхние челюсти у сфецид сильно склеротизированы и разнообразны по форме. Признаки строения жвал в некоторых родах весьма изменчивы и широко используются в систематике. Вершинные, предвершинные и мощные срединные зубцы на внутренней поверхности жвал в связи с гнездованием в плотных почвах и в древесине развиваются у многих представителей Sphecinae, Pemphredoninae (рис. 46з), Crabroninae (рис. 46и,к), Philanthinae. В то же время зубцы редуцируются при сокращении строительной функции жвал. Они маленькие или отсутствуют у ос, использующих главным образом ноги при строительстве гнезд (многие Nyssoninae, Larrini, некоторые Sphecinae), например, у Bembix.
Одной из важнейших специализаций у сфецид является развитие вырезки на на нижнем наружном крае у основания. Вырезка отсутствует у Ampulicinae, Sphecinae и pemphredoninae. Она имеется у Dinetini, развита у многих Crabroninae, Entomosericinae. У Nyssoninae и Philanthinae жвалы без вырезки.
Пальпы (щупики). Пальпы у сфецид, как правило, состоят из симметричных цилиндрических члеников, но у ряда представителей появляются различные модификации. У некоторых видов Prionyx лабиальные щупики сильно укороченные и состоят из 1-3 члеников. У Astata 2-й членик лабиальных щупиков асимметричный (также у некоторых видов Dryudella). У Nitela лабиальные щупики очень короткие, причем 1-й членик значительно длиннее, чем остальные три. Число члеников максиллярных и лабиальных пальп у большинства сфецид Казахстана соответственно равно 6 и 4 (пальпальная формула – 6+4), только у некоторых видов Tachytes равно соответственно 5 и 3, Rhopalum – 5 и 3, Bembix – 4 и 2 и Eremiasphecium – 5 и 3.
Затылок. Важным таксономическим признаком является форма затылочного киля, ограничивающего затылок. У более примитивных сфецид затылочный киль обычно имеет вид замкнутого кольца, отделенного от гипостомального киля явным промежутком. У более продвинутых форм он на большем или меньшем протяжении сливается с гипостомальным килем или редуцируется в нижней части, приобретая Uобразную форму. На концах его в таких случаях иногда образуются острые зубцы или выступы. Иногда киль редуцируется целиком.
Грудь. Общая форма. Грудь у роющих ос, как у всех других высших перепончатокрылых (Apocrita), состоит из трех истинно грудных сегментов и слившегося с ними 1-го брюшного, так называемого промежуточного, сегмента. Она, как правило, хорошо развита, имеет 3 пары ног и 2 пары крыльев. По общей форме можно выделить 2 типа груди: 1) тонкая и длинная и 2) широкая компактная, но имеются и промежуточные формы. Первый тип характерен для Dolichurus (рис. 47а), Sceliphronini, Ammophilini, Passaloecus (рис. 47б), Parapiagetia, Trypoxylon (рис. 47в), Liris (рис. 47г), Crossocerus, Alyssonini (рис. 47д); второй тип – для Astata (рис. 48в), многих представителей Crabroninae (рис. 48г), Nyssoninae (рис. 48д,е), Entomosericinae и Philanthinae (рис. 48ж). Общая форма зависит от особенностей строения составляющих грудь отделов.
Переднеспинка. У большинства сфецид этот отдел развит относительно слабо, но у Dolichurus и большинства представителей подсемейства Sphecinae довольно велик, особенно у Dolichurus. Верхне-задняя часть ее у сфецид обычно расширена в виде валика, это так называемый воротничок. Строение переднеспинки очень сильно варьирует среди Crabronini, и это находит отражение в систематике на видовом уровне.
Характер сочленения переднеспинки со среднеспинкой имеет значение для дифференциации некоторых близких родов.
Наличие явного промежутка между плечевыми буграми (пронотальными лопастями) и тегулами является одним из важных признаков, характеризующих семейство Sphecidae. Однако у Dolichurus плечевые бугры и тегулы почти соприкасаются, разделены очень узким промежутком или соприкасаются у большинства представителей Pemphredonini. У Astatini тегулы и плечевые бугры тоже почти соприкасаются.
Среднеспинка, или скутум. У большинства сфецид среднеспинка имеет вид равномерно выпуклого или отчасти уплощенного склерита, снабженного бороздами. Среднеспинка Dolichurus характеризуется глубоко вдавленными нотаулями, доходящими до заднего края склерита. У остальных сфецид нотаули развиты только в передней части или отсутствуют. Обычно на среднеспинке имеются околосрединные линии. У Pemphredoninae нотаули тоже имеются и развиты обычно только впереди, иногда длинные и почти доходят до заднего края, иногда почти не заметны (у некоторых видов Stigmus и Spilomena). Околосрединные линии иногда слиты в одну.
Скутум Larrini имеет боковую кайму, которая налегает на тегулу.
У Entomognathus среднеспинка в середине иногда слегка вдавленная и имеет глубокие ямки на нотаулях и парапсидальных бороздах. В задней части скутума у многих Crabronini, Gorytini развита поперечная борозда, иногда имеющая вид ряда ямок, разделенных короткими килями или складками. У некоторых родов Nyssoninae развит косой скутальный киль по бокам среднеспинки. У Mellinus околосрединные линии ограничивают выпуклое продольное срединное поле.
В систематике многих родов используются признаки скульптуры среднеспинки.
Щитик, или скутеллум. Щитик у сфецид имеет форму узкой (короткой) поперечной выпуклой пластинки. В систематике сфецид имеют значение следующие признаки щитика: степень его выпуклости, наличие или отсутствие базальной или апикальной борозды, выступов, килей, ямок, зубцов и скульптура.
Бока среднегруди. Признаки строения среднегруди широко используются в систематике роющих ос. Бока среднегруди у сфецид обычно имеют сложный набор борозд, ямок, швов и килей. Важнейшие из них следующие: эпистернальная борозда – проходящая от субалярной ямки на вертральную часть мезоплеврона, в полном виде достигающая антеро-вентральный край склерита (часто сливается со скробальной бороздой), омаулюс – киль, проходящий от нижнего края плечевых бугров в нижне-вентральном направлении; ацетабулярный киль – поперечный киль в передней нижней части среднегруди, часто соединяется с омаулюсом; стернаулюс – горизонтальный вентрально-латеральный гребень мезоплеврона выше сигнума – короткой линии, расположенной в латерально-вентральной части мезоплеврона; скробальная борозда – горизонтальная борозда (ямка), проходящая через скробу – ямку или точку, расположенную выше и позади середины мезоплеврона; мезоплевральный шов – задний край мезоплеврона, идущий от среднетазиковой впадины до основания крыльев; гиперстернаулюс – обычно горизонтальная ямка, расположенная в нижней части эпистернальной борозды; вертикаулюс – киль, проходящий впереди средних тазиков косо или вертикально вверх.
Расположение, количество и форма борозд, килей, швов, ямок и полей у сфецид варьирует в широких пределах. Иногда в систематике используются признаки, характеризующие степень выпуклости мезоплевр.
Признаки строения и скульптуры боков среднегруди широко используются в систематике на родовом и видовом уровнях, например, в подсемействах Pemphredoninae, Crabroninae, Nyssoninae и других таксонах.
Метанотум (заднеспинка, или заднещитик). Он имеет довольно однообразную форму, более или менее выпуклый или плоский. У Belomicroides на нем развиты боковые гребни, иногда в виде пластинок. У Belomicrus oн имеет чешуевидные выросты. У Oxybelus по бокам метанотума развиты чешуевидные или листовидные выросты, в середине часто проходит продольный киль. Грубая складчатая скульптура характерна для некоторых Nyssonini. На границе щитика и заднещитика часто имеются короткие продольные кили или складки.
Метаплевры (бока заднегруди). Обычно метаплевры в значительной мере редуцированы. У Dolichurus (Ampulicinae) метаплеврон обычно состоит из верхнего и нижнего метаплеврального поля, обычно различающихся скульптурой.
Стернальная область груди развита у роющих ос оченьслабо. Стернит переднегруди представлен, как правило, маленькой четырехлучевой пластинкой у основания передних тазиков. Стернит среднегруди тесно сливается с мезоплеврами, так что не видно даже швов. В середине его обычно развит продольный шов, являющийся наружным проявлением мощного стернального гребня энтоскелета. Стернум заднегруди слабо развит или отсутствует. У Laphyragogus он, в отличие от других сфецид, лежит в иной плоскости, чем мезостернальное поле, сильно выпуклый.
Промежуточный сегмент у сфецид, как и у других жалящих перепончатокрылых, состоит обычно лишь из хорошо развитого тергита.
Форма искульптура сегментаизменчивы в значительных пределах.
Крылья. Особенности строения птеростигмы и жилкования крыльев имеют большое значение в систематике роющих ос и при установлении филогенетических отношений таксонов. Особенности жилкования у разных представителей показаны на рис. 49 и 50.
Что касается птеростигмы, то среди роющих ос проявляются как тенденция к ее уменьшению, так и к относительному увеличению. Она маленькая обычно на крыльях с растянутым (распределенным почти по всей поверхности крыла) жилкованием, например, у Nyssoninae.
Птеростигма крупная у сфецид, имеющих крылья с концентрированным в базальной части крыла жилкованием, обычно в значительной мере редуцированным. Наиболее крупная птеростигма у Stigmus, Protostigmus, Ammoplanus, Ammoplanellus и некоторых других представителей трибы Pemphredonini.
Маргинальная ячейка у сфецид варьирует от длинной и острой на конце до короткой, усеченной или закругленной на вершине.
У некоторых представителей Astatinae, Laphyragoginae, Larrinae, Nyssoninae и Pemphredoninae вершина маргинальной ячейки отодвинута от края крыла.
Короткая маргинальная ячейка характерна для пустынных видов Astatinae и Crabroninae. Возможно, это связано с общей редукцией жилкования у видов открытых мест обитания из-за перехода к более наземному образу жизни (возможно, из-за частых сильных ветров).
Переднее крыло сфецид имеет 1-3 субмаргинальных ячейки. Три характерны для Ampulicinae (рис. 49а), Sphecinae (рис. 49б), Astatini (рис. 49в), Psenini, Larrini, большинства Nyssoninae (рис. 50г-е) и Philanthinae (рис. 50ж-и). Для ряда сфецид характерна стебельчатая форма 2-й или 3-й субмаргинальной ячейки (рис. 49ж, 50г,д,ж,и). Форма субмаргинальных ячеек в некоторых родах имеет таксономическое значение, например, в роде Tachysphex. Только две субмаргинальные ячейки характерны для крыльев некоторых видов Pemphredonini (рис. 49д), Dinetini (рис. 49г), Gastrosericus (Larrini) и многих видов Miscophus (рис. 49ж).
Только одна субмаргинальная ячейка свойственна переднему крылу Ammoplanops, Ammoplanellus и Ammoplanus (Pemphredoninae) (рис. 49е), части видов Miscophus и Nitela (Miscophini) (рис. 49з), Trypoxylon (Larrini) (рис. 50б) и Crabronini (рис. 50а).
У Oxybelini единственная субмаргинальная ячейка сливается с дискоидальными, вследствие чего образуется единая субмаргинальнодискоидальная ячейка.
У некоторых Pemphredonini и Miscophini нет ни одной замкнутой субмаргинальной ячейки (Bohart and Menke, 1976).
У Dolichurus, Astata, Dryudella и Laphyragogus внутри 1-й субмаргинальнойячейки имеется следили остаток редуцированной поперечной жилки, обычно в виде небольшого придатка дистальной жилки, которая в этом месте угловато изогнута (рис. 49в).
В трибах Pemphredonini, Miscophini, Oxybelini и Crabronini наблюдается большая или меньшая редукция дискоидальных ячеек. В подтрибах Ammoplanina и Stigmina (Pemphredonini) остается одна дискоидальная ячейка, т.к. исчезают 2-я возвратная жилка и задняя жилка 2-й дискоидальной ячейки (рис. 49е). 2-я дискоидальная ячейка исчезает также у некоторых видов Miscophus, Nitela (рис. 49з), Trypoxylon (рис. 50б), у всех Oxybelini и Crabronini (рис. 50а,в). У Oxybelini дискоидальная ячейка сливается с субмаргинальной (рис. 50в).
Субдискоидальная ячейка у части сфецид тоже испытывает редукцию. У Nitela (Miscophini) она открытая сзади, т.к. жилка A1 редуцировагна за жилкой cu-a (рис. 49з). У части видов Ammoplanus и Ammoplanellus (Pemphredonini) она тоже открытая, т.к. участок задней жилки редуцируется (рис. 49е).
Для сцепочного механизма у сфецид в качестве уникальной модификации можно указать разделение ряда крючков на 2 группы у представителей Trypoxylonini (кроме некоторых видов Pison).
Для системы жилок заднего крыла тоже свойственна тенденция к редукции. Она проявляется в основном в укорочении медиальной и субмедиальной ячеек, которое можно проследить почти во всех подсемействах сфецид. Редукция жилкования может выражаться в виде слабой склеротизации жилок, особенно в их дистальной части, или даже их полном исчезновении. Наиболее редуцированное жилкование можно обнаружить у некоторых родов Miscophini и Pemphredonini. У Ammoplanops и Ammoplanellus жилки часто очень слабо склеротизированы (рис. 49е). Среди Miscophini наиболее редуцированное жилкование у Nitela (рис. 49з). В этом роде часто даже костальная ячейка является незамкнутой, а жилка M+Cu представлена лишь коротким базальным остатком.
Одной из первых в заднем крыле у сфецид подверглась редукции субкостальная жилка. Она иногда имеется у Ampulicinae и Astatinae. У остальных роющих ос отсутствует.
Югальная лопасть у сфецид имеет разные размеры. У Ampulicinae она маленькая, у Sphecinae обычно большая, у Crabroninae она иногда бывает крупной, но обычно маленькая, у некоторых родов отсутствует совсем. У Nyssoninae обычно маленькая, иногда исчезает, у Philanthinae размер ее варьирует от большого до маленького. Очень крупная югальная лопасть у Astata (рис. 49в). У Pemphredoninae она обычно имеет средние, реже маленькие размеры.
Ноги. Признаки строения ног широко используются в систематике сфецид, особенно такие как наличие и форма выступов, зубцов, килей, шипов идругих подобных образований.
Внутри семейства сфецид проявляется тенденция к редукции наружной шпоры средних голеней, так что остается только одна внутренняя. Редукция наблюдается среди Ammophilini (у некоторых видов Podalonia, у Eremochares), у Pemphredoninae, Crabroninae (часто исчезает и внутренняя шпора), у некоторых Nyssoninae (Oryttus, некоторые виды Kohlia, Stizini, Bembicini), Philanthinae. У Alyssonini и Bembicini иногда редуцируются обе шпоры. Густота щетинок (или зубцов) на шпорах голеней у некоторых таксонов имеет диагностическое значение.
У многих роющих ос на лапках самок развивается особый ряд длинных щетинок, который используется при строительных работах (у ос-землекопов) в качестве граблей. Такое образование характерно для многих Sphecinae (рис. 51а,б), некоторых Pemphredoninae, Astatinae, Crabroninae (рис. 51в,г), для Entomosericinae, большинства Nyssoninae (рис. 51д) и Philanthinae (рис. 51в,ж). С ним часто связана асимметрия члеников передних лапок (рис. 51а,б,е,ж,з). Форма отдельных члеников лапок или лапок в целом весьма разнообразна и потому часто используется при дифференцировании родов и видов.
Для лапок многих относительно примитивных сфецид характерны плантулы – маленькие овальные подушечки на вершине члеников снизу
(рис. 52а,б).
Коготки вершинного членика лапок у многих сфецид имеют зубцы. Их число варьирует от 1 до 5 (рис. 52в-д). У некоторых видов Bembix коготки на одной лапке разные по величине (рис. 52е).
На конце лапок у роющих ос обычно развиты пульвиллы (рис. 52ж). Они могут быть крупными у некоторых Sphecinae, Larrini, Alyssonini. У самок некоторых Gorytini они значительно крупнее на передних ногах, чем на средних и задних.
На передних ногах и нижней части головы по бокам у некоторых Sphecinae, Crabroninae и Astatinae развит так называемый псаммофор – система крепких щетинок, образующих корзиночку для перетаскивания рыхлой земли.
Брюшко (метазома). У Sphecidae вершина брюшка может модифицироваться таким образом, что часть сегментов втягивается внутрь брюшка и становится скрытой. Особенно далеко этот процесс зашел у самцов Ampulicinae, у которых видны снаружи только 3 сегмента, хотя у самок остаются видны 6 сегментов, что является правилом для самок всех сфецид. У самцов большинства остальных роющих ос снаружи видны 7 сегментов, но у некоторых Sphecinae и Nyssoninae только 6. У некоторых Nyssoninae (например, Alyssonini) наружу торчат шипы 8-го стернума. У многих представителей Nyssoninae торчащий наружу конец 8-го стернума имеет вид загнутого шипа («ложное жало»). Среди Nyssoninae число видимых снаружи тергумов у самцов сокращено до 6 у некоторых видов Hoplisoides.
Брюшко по своей форме обычно овальное, иногда впереди расширенное до треугольного. По способу прикрепления брюшка к груди, т.е. по строению 1-го сегмента брюшка (истинного 2-го) оно бывает висячим или стебельчатым. Стебелек может состоять из 1-го тергума и 1-го стернума, или только из 1-го стернума. У Ammophila он состоит из 2 стернумов и 1 тергума (рис. 52з,и). Особенности строения стебелька имеют диагностическое значение.
К числу особых признаков тергумов у отдельных родов относятся: вершинные поперечные вдавления на тергумах (Crabroninae и др.), продольный киль на основании 2-го тергума (Larrini), поперечные базальные опушенные ямки на 2-4-м тергумах самца (Belomicroides), острый боковой изгиб 1-3-го (иногда до 5-го) тергумов (Belomicrus), большие латеротергумы на 1-м и 2-м тергумах (Entomosericus), субапикальные вдавления на 2-4-м (иногда и следующих) тергумах (Entomosericus), сокращенные до узкой полосы латеротергумы 1-го тергума (Mellinus), сильная изогнутость 1-го тергума у вершины (Lestiphorus), зубцы на вершине 7-го тергума (Nyssonini), большие дыхальцевые лопасти на 7-м тергуме самца (Stizini, большинство Bembicini), лопасти или выступы на вершине 7-го тергума самца (Bembicini).
Одной из важнейших особенностей вершины брюшка у многих роющих ос является наличие на 6-м тергуме самки или 7-м тергуме самца особого, так называемого пигидиального поля. Оно имеет разнообразную форму, скульптуру и опушение. Строение его связано со строительными работами ос. У большинства сфецид, строящих гнезда в земле, оно имеет вид плоского треугольника или четырехугольника, ограниченного по бокам килями (рис. 52е-м). У Ampulicinae и Sphecinae оно отсутствует. У Pemphredoninae хорошо развито, имеет или треугольную форму (рис. 53а,б,в), или форму трапеции либо прямоугольника (рис. 53г,д). У Pemphredon оно узкое и немного вдавленное, изредка редуцированное до срединного поля (рис. 53е).
У Astatinae пигидиальное поле имеется, иногда слабо развитое. У Crabroninae оно обычно имеется, у самок плоское и треугольное или желобообразное и вогнутое (в зависимости от характера гнездования) (рис. 53ж-и), а у самцов обычно трапециевидное, реже прямоугольное или квадратное, иногда отсутствует.
Пигидиальное поле хорошо развито у самок Entomosericus (Entomosericinae). Среди Nyssoninae оно имеется у самок Mellinus, явственно ограничено килями и покрыто щетинками у самок Alyssonini, хорошо выражено у Nyssonini и Gorytini. У Stizini и Bembicini оно отсутствует или только очень слабо выражено сзади, редко четко ограничено килями и занимает большую часть поверхности тергума.
Среди Philanthinae пигидиальным полем обладают многие Philanthini, Pseudoscoliini и Cercerini, и его форма, а также скульптура имеют большое таксономическое (диагностическое) значение.
В качестве специализированных признаков стернумов у сфецид можно отметить следующие: вельветообразный ворс на стернумах самцов; длинные отстоящие щетинки у самцов; киль или кровлеобразная выпуклость последнего стернума самки; апикальная бахрома волосков на 4-м (иногда и 3-м) стернуме самцов; пучки волосков на стернумах у самцов; U-образное вдавление на 2-м стернуме; уплощенные овальные площадки на основании 2-го стернума самки, разделенные продольным килем; срединный выступ на заднем крае 2-го стернума; вдавления на 37-м стернумах; продольный киль на 1-м стернуме; перевязь длинных волосков (бахрома) на дне неглубокого поперечного вдавления; косые углубления на 3-4-м стернумах; бахрома длинных волосков на вершине стернумов; широкое полукруглое апикальное вдавление с длинными прилегающими волосками на 2-3-м стернумах; поперечные валики или кили на 2-4-м стернумах самцов; поперечные кили, бугры, выступающие площадки на 2-м стернуме; бугорки или валики на стернумах; пластинчатый киль на 6-м стернуме самки; выступ разной формы на 2-м стернуме самцов ; продольная ямка и треугольный базальный выступ; апикальные щетки волосков на 3-м стернуме и изогнутый наружу двойной гребень на 6-м стернуме самок; двойной или одинарный киль на основании 1-го стернума; редукция (слабая склеротизация, укорочение) 7-го стернума; бугровидный или шиповидный выступ или угловатая выпуклость на 2-м стернуме; модифицированный 6-й стернум; апикальные пучки волосков на 2-5-м стернумах у самцов; короткая апикальная бахрома густых волосков на 3-4-м стернумах; горбообразный выступ на 2-м стернуме; базальные (обычно скрытые) пучки волосков на 5-м и 6-м стернумах самцов; выступ на 2-м стернуме самцов; выступ разной формы на 6-м стернуме самцов; срединное углубление и апикальная вырезка на 6-м стернуме самок; глубокая выемка на вершине 6-го стернума самок; выступающая площадка у основания 2-го стернума; углубление и вертикальная, часто вырезанная пластинка на 5-м стернуме самки; выступы, пучки и бахрома волосков на заднем крае вершинных стернумов; поперечная борозда и киль у основания или в середине 2-го стернума и др. Варьирование строения 6-го стернума самцов у Ammoplanus дает надежные признаки для различения видов в этом роде.
Большое значение в систематике сфецид имеет окраска брюшка, особенно четко различающаяся по видам в трибах Gorytini, Nyssonini, Bembicini, Cercerini, Philanthini, Pseudoscoliini и некоторых других, а также скульптура.
Различные морфологические модификации приобретает у сфецид 8-й стернум самцов. Обычно он имеет различные выступы, вырезки, лопасти и т.д. У многих Psenini он образует тонкий острый выступ, дугообразно загнутый вверх (так называемое «ложное жало» - pseudosting). 6-й стернум самки у Dolichurus (Ampulicinae) удлинен и образует почти замкнутую дорзально трубку, через которую жало торчит наружу.
Последним видимым снаружи сегментом у самки является истинный седьмой (считая первым промежуточный сегмент). Его форма обычно конусообразная. Сзади между его тергумом и стернумом имеется щель, ведущая в большой, ограниченный мембраной мешок, лежащий внутри сегмента. Внутри мешка помещается жало, анальное отверстие и все задние сегменты, начиная от восьмого.
Восьмой стернум сильно редуцирован, но на нем сохраняется дыхальце, вокруг которого расположено склеротизированное поле, носящее название стигмальной пластинки. В стернальной перепончатой части находится отверстие половой камеры, в которую открываются яйцевод спереди и семеприемник сверху. Яйцо выходит из полового отверстия, минуя жало. Бока 9-го тургума склеротизированы и образуют пару характерных квадратных пластинок, которые в покое частично прикрыты стигмальными пластинками. 9-1 стернум перепончатый и прилегает к основанию створок жала.
Строение самого жала у сфецид очень сильно напоминает таковое у медоносной пчелы. 1-я яйцекладная пластинка обычно невелика, имеет треугольную форму и называется треугольной пластинкой. Ее задний конец несет 2 сочленовные головки: верхняя сочленена с квадратной пластинкой (с 9-м тергумом), нижняя – со 2-й яйцекладной пластинкой. От переднего конца 1-й пластинки отходит эластичный стержень, который, описав большую дугу, переходит в 1-ю створку жала. 2-я яйцекладная пластинка довольно длинная и называется продолговатой пластинкой. От ее переднего конца также отходит эластичный стержень, переходящий в слитную вторую створку жала, от заднего конца отходит 3-я створка. 3-я створка желобообразна и служит футляром для жала.
Первые и вторые створки соединены скользящим швом – вдоль второй створки идут 2 рельсовидных валика, охваченные желобами первых створок. Между первыми и слитной створками проходит канал для яда. В основной части слитной створки он расширен, в соответствии с чем створка образует вздутие, или барабан. От первых створок на уровне барабана отходит вверх короткий поперечный отросток. Он помещается в полости барабана и при движении створок относительно друг друга передвигается внутри нее. Функция его состоит в нагнетании яда из полости барабана в более узкую дистальную часть канала. Кроме того, наличие поперечных отростков делает невозможным прохождение яйца по каналу, и, таким образом, жало утрачивает первичную и основную функцию яйцеклада (Шванвич, 1949). Резервуар ядовитой железы открывается в основание барабана. Признаки строения жалящего аппарата самок в систематике роющих ос почти не используются.
Брюшко самцов роющих ос оканчивается 7-м (настоящим 8-м) тергумом вверху и 7-м или 8-м стернумом внизу. 7-й тергум самцов часто имеет на конце зубцы, выступы, вырезки и т.п., что используется в таксономии на видовом уровне. Имеет значение в систематике и общая форма тергума. 7-й стернум самцов роющих ос не редуцированный. 8-й тергум более или менее редуцированный, в некоторых случаях представляет собой цельную пластинку, в других – разделен на 2 боковых склерита. 9-й сегмент представлен узким мембранозным полем. У некоторых представителей он несет пару тонких (слабо развитых) пигостилей, у отдельных видов Bembix – пару толстых волосистых лопастей. Анус на конце 9-го сегмента представляет собой наружное отверстие короткой полости, в которую открывается ректум (Snodgrass, 1941).
Гениталии самцов имеют очень разнообразную форму. Признаки их строения (форма вальв , вольселлы, эдеагуса и парамер) используются в систематике сфецид для дифференциации родов и видов.
Фаллус сфецид очень сильно варьирует по относительным размерам и форме его частей. Базальное кольцо располагается или антеровентрально, или прямо внизу, под основанием парамер и вольселл. Парамеры обычно слиты с парамеральными пластинками, но иногда частично разделены. Эдеагус имеет хорошо развитые боковые лопасти – вальвы пениса (пениальные створки). Вольселлы представлены обычно широкими склеритами, целиком отделившимися от парамеральных пластинок, но часть соединенными друг с другом склеротизированным мостом. Каждая вольселлярная пластинка несет куспис и дигитус. Дигитус особенно сильно варьирует по размерам и форме. Иногда куспис очень маленький или отсутствует.
В целом у сфецид проявляется тенденция к потере подвижности и редукции отдельных частей вольселлы и ее полному исчезновению.
Форма самого эдеагуса очень разнообразнпа, в целом удлиненноцилиндрическая, с расширением (головкой) на вершине. Часто он вооружен зубцами, щетинками, выступами и лопастями.
8-й тергум самцов некоторых примитивных древних групп роющих ос имеет церки (Dolichurini, большинство Sceliphronini), у остальных сфецид лишен их. Исключение представляют Eremiaspheciini (Philanthinae), хотя возможно, что их церки представляют собой образование, не гомологичное церкам примитивных сфецид.
Яйцо у роющих ос имеет овально-цилиндрическую форму. Оно белое или слегка желтоватое, как правило, слегка изогнутое в середине и имеет закругленные концы. Размеры яйца внутри семейства варьирует в значительных пределах: от 0,3 до 6 мм. Интересно, что у крупных видов Larra яйцо относительно маленькое, что, очевидно, связано с высокой активностью хозяина, на которое оно откладывается (маленькое яйцо меньше подвержено опасности повреждения, чем большое).
Многие авторы отмечают сильную изогнутость яиц у Crabronini. Это, вероятно, связано с тем, что яйцо лежит на шее жертвы, охватывая ее полукольцом.
Относительно маленькое яйцо у ос рода Nysson. Это, вероятно, связано с клептопаразитизмом этих ос.
Личинка. Взрослая личинка роющих ос имеет изогнутую форму, желтовато-белую окраску, состоит из головы и сегментированного тела. Голова часто имеет коричневые пятна. Глаза отсутствуют, усики редуцированы до простых ямок, содержащих сенсиллы, или представлены в виде более или менее развитых односегментных папилл. Мандибулы хорошо развиты, снабжены мощной мускулатурой, расположенной внутри головной капсулы. Максиллы и нижняя губа имеют форму мясистых лопастей, несущих конические пальпы, нижняя губа содержит прядильный орган, который имеет форму поперечной щели или пары небольших выступов, с которыми связаны выводные протоки лабиальных шелковых желез. Тело лишено ног, но снабжено 10 парами маленьких дыхалец.
Куколка у роющих ос, как и у всех перепончатокрылых, открытого типа. В ней видна голова с усиками, приложенными к вентральной стороне тела, вполне сформировавшиеся ноги, прижатые к телу, и маленькие крылья, расположенные вдоль боковых поверхностей груди.
К числу важнейших сторон биологии сфецид относятся тип гнездования (включающий выбор места для гнездования, характер субстрата, строение гнезда и способ его постройки), выбор добычи, тип охотничьего поведения (включающий поиски добычи и способы ее обработки), способ транспортировки добычи и втаскивания ее в гнездо, общий порядок актов поведения, связанных с заботой о потомстве, тип питания имаго, характер взаимоотношения полов, отношение гнездящихся самок друг к другу и фенологический цикл.
Гнездование. По способу гнездования среди роющих ос Казахстана выделяются 4 группы: 1) устраивающие гнезда в земле, 2) устраивающие их в древесине и стеблях растений, 3) изготавливающие свободные лепные гнезда из влажной земли и 4) клептопаразиты.
Для наиболее примитивных роющих ос характерно гнездование в готовых полостях (в земле, в камнях, скалах, стенах домов и других построек), в щелях между камнями и в других подобных местах. Такой выбор мест для гнезд обнаружен у видов Dolichurus и некоторых примитивных представителей подсемейства Sphecinae и трибы Larrini. Щтдельные виды Pemphredoninae и Crabroninae отыскивают для устройства гнезд норки насекомых и других мелких животных, причем возможно, что некоторые из них перешли к такому гнездованию вторично – от самостоятельного рытья или выгрызания норок в субстрате.
В земле гнездятся почти все виды Sphecinae, из Pemphredoninae – Mimesa, Diodontus, Ammoplanus, все Astatinae, Larrini, Palarini, Miscophini, Oxybelini, часть Crabronini (Entomognathus, Lindenius, Crabro, часть Lestica и Crossocerus, все Nyssoninae и Philanthinae.
Участки почвы, на которых устраиваются гнезда, могут различаться в значительных пределах по влажности, химическому и механическому сосьтаву, степени наклона и освещенности поверхности, температурному режиму и другим показателям.
В древесине, обычно более или менее сгнившей (трухлявой), и стеблях растений с мягкой сердцевиной или готовых полостях в древесине и стеблях растений гнездятся все виды родов Mimumesa, Psen, Psenulus, Pemphredon, Passaloecus, Stigmus, Spilomena, Trypoxylon, Nitela, Pison, Rhopalum, Ectemnius, часть видов Crossocerus, Lestica, возможно, также некоторые виды Solierella, причем большинство из них в той или иной степени используют уже готовые полости: ходы насекомыхксилофагов, заполненные буровой мукой, или пустые гнезда других перепончатокрылых. Некоторые виды Rhopalum, Crossocerus и ectemnius могут гнездится в сердцевине живых растений. Отдельные виды (в частности Ectemnius fossorius и Crossocerus annulipes) могут гнездится и в древесине, и в земле (на обрывах).
Многие роющие осы, которые гнездятся в готовых полостях в древесине и сердцевине растений, обычно могут также гнездиться в полых стеблях растений, а некоторые из них – также в пустых галлах насекомых на стеблях растений. Это говорит о близости свойств этих мест гнездования и о легкости перехода к гнездованию в субстратах, имеющих полости. Однако среди ос отмечены виды, гнездящиеся в строго определенном типе стеблей. Таковы некоторые виды Solierella, Nitela, Pison, Trypoxylon, Crossocerus и Ectemnius, которые гнездятся в тонких полых стеблях однолетних растений. Среди Sphecinae гнездование в полых стеблях отмечено в роде Chalybion.
Группа ос, строящих лепные гнезда из глины на скалах, камнях, стенах построек и на обрывах, самая малочисленная. Она включает только 5 видов рода Sceliphron, хотя виды родов Chalybion и Trypoxylon тоже используют влажную глину, но только для изготовления внутренних перегородок между ячейками и наружной затычки.
К группе ос, гнездящихся в земле, примыкает группа осклептопаразитов, которые сами не строят гнезд и не охотятся, а откладывают яйца в ячейки других ос, строящих гнезда в земле. К ним относятся виды родов Nysson, Synnevrus, Brachystegus и Stizoides. Сюда же можно отнести примитивных ос, использующих готовые полости в земле (Dolichurus), а также виды, которые используют норки своих жертв
(Larra).
По морфологическим особенностям гнезда роющих ос Казахстана делятся на 6 типов: 1) единственная ячейка в готовой полости в субстрате, 2) одноячейковые простые гнезда в полости, сделанной осой в субстрате, 3) многоячейковые разветвленные гнезда, 4) линейные многоячейковые гнезда, 5) разветвленно-линейные и 6) многоячейковые свободные гнезда из глины.
Готовые полости в земле для прятания добычи, на которую откладывается яйцо, используют виды рода Dolichurus. Виды родов Liris, Ancistromma и Chlorion часто тоже устраивают свои гнезда в готовых полостях в земле, хотя потом производят некоторую обработку их стенок.
Простые одноячейковые гнезда в субстрате (насколько известно, в земле) устраивают осы из родов Palmodes, Prionyx, Podalonia, Eremochares, Ammophila, Gastrosericus, вероятно, также Parapsammophila, иногда виды родов Dryudella, Dinetus, Liris, Tachytes, Tachysphex, Palarus, Plenoculus, Oxybelus, Argogorytes, Hoplisoides, Stizus, Bembix. Среди гнезд ос родов Prionyx и Ammophila, впрочем, известны и многоячейковые. Виды рода Larra откладывают яйца на медведок, которые после прекращения временного паралича от ужаления осой просто вползают в вою норку, и последняя становится убежищем для развивающейся личинки осы.
Многоячейковые разветвленные гнезда в земле сооружают виды родов Sphex из Sphecinae; Mimesa, Diodontus, Ammoplanus из Pemphredoninae; Diploplectron, Astata, Dryudella, Dinetus из Astatinae; Liris, Tachytes, Tachysphex, Prosopigastra, Palarus, Plenoculus, Miscophus, Belomicrus, Oxybelus, Entomognathus, Lindenius, Crabro, часть видов Crossocerus и Lestica из Crabroninae; все видв рода Mellinus, триб Alyssonini, Entomosericini, Gorytini, Stizini из Nyssoninae, подсемейства Philanthinae, часть видов рода Bembix. Некоторые осы среди перечисленных родов делают и одноячейковые гнезда, хотя, возможно, в ряде случаев, когда было зарегистрировано одноячейковое гнездо, оно не было достроено до конца. Только в отношении некоторых видов (например, Bembix rostrata) твердо установлен факт постройки одноячейкового гнезда, что позволяет рассматривать его как вторично упрощенное или модифицированное (гнездо усложнено изготовлением особого отнорка – «шпоры») (Evans, 1957b).
Линейные многоячейковые гнезда (насколько известно, в ходах ксилофагов в древесине, полых или имеющих мягкую сердцевину стеблях растений и в гнездах других насекомых) устраивают виды родов Chalybion, Psen, Psenulus, Stigmus, Spilomena, Nitela, Mimumesa, Pison, Trypoxylon, Rhopalum, Ectemnius и часть видов Lestica и Crossocerus, возможно, и некоторые виды Solierella. Большинство из перечисленных сфецид предпочитают использовать уже готовые трубчатые полости в древесине и стеблях растений (ходы, галлы и гнезда других насекомых, полые стебли и т.п.), перегораживая их на ячейки, но многие виды родов Pemphredon, Ectemnius, Crossocerus могут выгрызать трубчатые полости для гнезд самостоятельно, хотя выбирают не очень плотный субстрат – мягкую сердцевину стеблей растений или трухлявую древесину.
Линейно-разветвленные гнезда среди ос, гнездящихся в земле, отмечены у видов родов Astata, Entomognathus, Diodontus, а среди ос, гнездящихся в древесине, - у видов родов Pemphredon, Crossocerus, Ectemnius и Lestica. Возможно, их строят также некоторые виды родов Mimumesa, Psen и Mimesa. Такое строение гнезд, возможно, обусловлено плотностью субстрата и значительно уменьшает объем работы, т.к. делает ненужным изготовление боковых отнорков в каждой ячейке. В то же время, в ряде случаев оно определяется разветвленным строением системы уже готовых ходов других насекомых (например, жуковксилофагов) в субстрате.
Лепные многоячейковые гнезда из влажных комочков земли, собираемой на берегу водоемов, как уже упоминалось выше, делают осы рода Sceliphron.
У ос-клептопаразитов (Nyssonini, Stizoides) инстинкт постройки гнезд утрачен.
Способы постройки гнезд у роющих ос делятся на мандибулярный, псаммофорный, тарзальный, пигидиальный и лепной мандибулярный в зависимости от органов, использование которых преобладает.
Мандибулярный способ постройки характерен для видов родов Sphex, Palmodes, отчасти Prionyx, Ammophila. При этом способе частицы субстрата выносятся из норки с помощью жвал.
Псаммофорный способ отмечен в родах Eremochares, Ammophila,
Dinetus, Belomicrus. Рыхлый субстрат выносится в «корзинке», образованной щетинками нижней части головы, челюстями и передними ногами с их хорошо развитыми щетинками. Отдельные камешки и комочки земли выгрызаются и выносятся в челюстях, рыхлый субстрат может также отгребаться передними ногами.
Пигидиальный способ наиболее характерен для гнездящихся в знмле Pemphredoninae, многих Larrinae, Crabroninae, Philanthinae и некоторых Nyssoninae. Частицы субсьтрата выталкиваются в основном с помощью пигидия, хотя отчасти и с помощью передних ног и жвал.
Между строением пигидиального поля и характером субстрата существует тесная связь. Часто по строению пигидия можно судить о том, как и в каком субстрате гнездится та или иная оса. У ос, гнездящихся в земле, пигидиальное поле плоское, обычно треугольное, тогда как у ос, гнездящихся в древесине или стеблях растений, оно более или менее суженное и вогнутое в виде желобка.
Пигидии ос, гнездящихся в древесине и стеблях растений, более многообразны по форме, чем у ос, гнездящихся в земле. По строению пигидия А.К.Мерисуо (Merisuo, 1936) выделил среди сфецид несколько морфо-биологических типов: среди ос, гнездящихся в древесине и стеблях растений, - тип Solenius, тип Coelocrabro и тип Pemphredon; среди ос, гнездящихся в земле, - тип Dinetus, тип Cuphopterus и тип Ammophila.
Тарзальный способ характерен для большинства Crabroninae, Nyssoninae, части Sphecinae. Часто сочетается с пигидиальным и мандибулярным. Частицы субстратавыбрасываются из норки с помощью передних лапок, обычно имеющих гребень густых и длинных щетинок.
Наивысшим совершенством этого приема приема обладают представители рода Bembix. Быстрота работы их передних ног такова, что песок сыпется мз-под тела осы непрерывной струйкой. Кстати, у Bembix передние ноги работают синхронно (в такт), отчего происходят одновременные кивающие движения тела.
Лепной мандибулярный способ характерен для видов родов Sceliphron, Chalybion, Trypoxylon, которые приносят для постройки перегородок или целиком гнезд комочки влажной земли (грязи).
Наконец, некоторые виды не строят норки, а используют уже готовые полости в субстрате. Не строят норки и клептопаразиты. Виды, которые используют готовые полости в субстрате и у которых строительные инстинкты слабо развиты или редуцированы, принадлежат к роду Dolichurus, где такое гнездование носит первичный характер, роду Chalybion, где, по-видимому, оно тоже первично, а также родам Mimumesa, Psen, Psenulus, Pemphredon, Passaloecus, Stigmus, Spilomena, Solierella, Pison, Nitela, Trypoxylon, Crossocerus, Rhopalum, Ectemnius и Lestica, где этот способ гнездования носит вторичный характер.
Среди роющих ос Казахстана наиболее распространен тарзальный способ, хотя группа видов с пигидиальным способом тоже большая. Псаммофорный способ натболее характерен для облигатных обитателей песчаных пустынь.
Для многих ос свойственно применение особого инструмента, так называемого «вибратора», идентичного отбойному молдотку (Мариковский, 1961). Суть его действия заключается в том, что вибрация крыловых мышц, расположенных в груди, через систему трахей и их расширений передается жвалам. Вибрация помогает отрывать жвалами частицы субстрата.
Для некоторых ос отмечено использование задних ног для выталкивания частиц субстрата из норки (Alyssonini, Gorutini, Cercerini). У них на вершине задних голеней обычно имеется уплощенная лопастевидная площадка, скульптура которой сходна со скульптурой пигидиального поля (Bohart and Menke, 1976).
Некоторые осы могут выталкивать частицы субстрата из гнезда с помощью головы (Crabronini).
Многие роющие осы при изготовлении пробок и перегородок в гнезде используют различные инородные материалы.
Добыча. Добыча роющих ос Казахстана и Средней Азии относится, насколько известно, к 11 отрядам насекомых и одному отряду паукообразных. На тараканов (Blattoptera) охотятся виды рода Dolichurus и 1 вид рода Tachysphex, на пауков (Aranei) – виды Chalybion, Sceliphron, Miscophus, Pison и Trypoxylon, на прямокрылых (Orthoptera) – виды Sphex, Palmodes, Prionyx, Eremochares, Larra, Liris, Gastrosericus, Ancistromma, Tachytes, Tachysphex и Stizus. Некоторые виды (Tachysphex, Stizus и Eremochares dives) охотятся также на богомолов (Mantoptera). Среди охотников на прямокрылых одни специализируются в добывании сверчков – Gryllidae (Liris, Ancistromma), другие ловят кузнечиков – Tettigoniidae (Sphex), третьи – саранчовых – Acrididae (Prionyx, Eremochares, Tachytes, Tachysphex, Stizus). Виды рода Larra отыскивают и парализуют медведок – Gryllotalpidae, а виды рода Gastrosericus – триперстов – Tridactilidae.
На клопов охотятся виды Astata, Diploplectron, Dryudella, Dinetus, Prosopigastra, Plenoculus, некоторые виды Lindenius (например, L. albilabris), возможно, также некоторые виды Solierella и Belomicrus, на равнокрылых (Homoptera) – виды родов Mimesa, Mimumesa, Psen, Psenulus, Diodontus, Pemphredon, Passaloecus, Stigmus, некоторые виды Crossocerus (например, C. annulipes, C. capitosus, C. wesmaeli, виды триб Alyssonini, Gorytini, Entomosericini, рода Bembecinus, возможно, также некоторые виды Spilomena и Nitela. Перечисленные выше представители трибы Psenini и подсемейства Nyssoninae охотятся на цикадовых (Cicadinea), представители трибы Pemphredonini – на тлей (Aphididae).
На двукрылых (Diptera) охотятся виды Oxybelus, Rhopalum, Crabro, Ectemnius, Mellinus, Bembix, некоторые виды Lindenius (например, L. panzeri) и Crossocerus.
Сеноеды (Copeognatha) в качестве добычи известны у некоторых видов Solierella, Nitela, Rhopalum, Crossocerus, но достоверные сведения об охоте на сеноедов ос Средней Азии и Казахстана пока отсутствуют.
Трипсов (Thysanoptera) собирают виды Ammoplanus, Eremiasphecium, некоторые виды Spilomena, возможно, также виды родов Ammoplanops, Ammoplanellus и Protostigmus.
На жуков (Coleoptera) охотятся некоторые виды Belomicrus, виды Entomognathus и большинство видов Cerceris.
Перепончатокрылых (Hymenoptera) ловят Palarus, Philanthus, Pseudoscolia, часть видов Cerceris, некоторые виды Lindenius (например, L. pygmaeus). Личинки пилильщиков (Tenthredinidae) являются добычей некоторых видов рода Ammophila (в частности, A. campestris).
На взрослых бабочек охотятся виды рода Lestica, возможно, также некоторые виды Ectemnius; на гусениц бабочек (Lepidoptera) – большинство Ammophilini, а также некоторые виды Tachytes (например, T. bidens), возможно, виды Parapiagetia.
Добыча ос родов Parapsammophila, Protostigmus, Ammoplanops, Ammoplanellus, Belomicroides, Olgia, Kohlia, Philanthinus остается неизвестной. Виды Nysson, Synnevrus, Brachystegus и Stizoides не охотятся, а являются клептопаразитами.
Наиболее распространенной добычей роющих ос Казахстана являются прямокрылые, равнокрылые и двукрылые. Много видов охотятся на жуков, перепончатокрылых, клопов, пауков и бабочек (в основном гусениц). Трипсы, богомолы, сеноеды и тараканы в качестве добычи используются немногими осами.
Количество добычи, запасаемой в одной ячейке, колеблется в широких пределах – от 1 до 100 и более, что зависит, вероятно, от многих причин: вместимости ячеек, наследственных факторов, определяющих поведение ос, величины и веса каждого экземпляра добычи и пр. Использование только одного экземпляра характерно для видов родов Dolichurus, Larra, некоторых видов Sphex, Palmodes, Prionyx, Podalonia, Ammophila, Ancistromma, Tachytes и Tachysphex. У одних видов, а именно у тех, кто устройством гнезда занимается после охоты, запасание одного экземпляра является правилом (например, у видов родов Dolichurus и Podalonia), у других -–имеет характер исключения и происходит в случае поимки особенно крупной жертвы.
У большинства видов количество добычи в одной ячейке не выходит за пределы 20-30 экземпляров. До 50 экзнмпляров и даже больше приносят в каждую ячейку некоторые виды родов Chalybion, Psenulus, Pemphredon, Passaloecus, Spilomena, Trypoxylon, Crossocerus, Hoplisoides, Bembix, Rhopalum и Cerceris. Максимальное количество (110) отмечено у Rhopalum gracile, запасающего очень мелких двукрылых (мокрецов).
В общем, количество экземпляров добычи в одной ячейке не является строго постоянной величиной, не считая случаев, когда запасается только одна жертва (например, у Ampulicinae и многих Sphecinae).
Интересный факт снабжения ячеек разным количеством жертв в зависимости от пола будущего потомства (например, у Philanthus triangulum) подтверждает предположение о зависимости числа жертв от вместимости ячеек, поскольку ячейки осой строятся разных размеров сообразно полу будущего потомства, а пол определяется тем, оплодотворено яйцо или нет.
Примечательно, что размеры имаго Lestica очень сильно варьируют, что, вероятно, объясняется разным количеством провизии, съеденной личинкой за время развития. В этом случае пол будущего потомства не связан с размерами ячейки и величина взрослой личинки (и имаго) зависит от случайного совмещения пола яйца, размеров ячейки и количества провианта в ней.
У ос, снабжающих потомство провизией по прогрессивному методу (т.е. по мере роста личинки), например, у видов рода Bembix, количество приносимых жертв зависит главным образом от хода развития личинки. Состояние личинки определяет программу охотничьей деятельности осы на день. Иногда оса натаскивает столько запасов, что личинка к некоторым жертвам даже не прикасается. Вероятно, такая особенность является своеобразной страховкой, поскольку позволяет личинкам нормально питаться, когда оса из-за неблагоприятной погоды не охотится.
Охота. Большинство роющих ос Казахстана охотятся на насекомых, живущих на вегетативных частях растений, что и определяет места их охоты. Таковы все виды, охотящиеся на прямокрылых, клопов, равнокрылых, гусениц, бабочек и личинок пилильщиков, трипсов и жуков. Они в поисках добычи переходят или перелетают от одного растения к другому, при этом, по-видимому, главную роль играют зрительные стимулы. Охотники на тараканов и сверчков ловят своих жертв на поверхности земли и в различных укрытиях (трещинахб выемках, полуоткрытых полостях, норках, между камнями и т.п.). Осы рода Larra ищут медведок в норах, в которых те живут. Виды рода Podalonia, охотящиеся на гусениц подгрызающих совок, могут выкапывать своих жертв из-под земли (при этом отыскивают своих жертв, руководствуясь, вероятно, обонянием).
Охотники на двукрылых ловят своих жертв, как правило, на растениях, причем в основном на цветках, где мухи питаются нектаром, но могут ловить и на земле. Mellinus arvensis часто охотится на мух, сидящих на экскрементах домашних животных. Осы родов Bembix и Oxybelus часто хватают мух прямо с животных (млекопитающих), которых те питаются.
Осы, охотящиеся на перепончатокрылых, ищут или караулят их на цветущих растениях.
Охотники на пауков разыскивают своих жертв на растениях, причем часто снимают их с ловчей сети. Chalybion sp., охотящийся на каракуртов, забирается за своей жертвой прямо в укрытие, где прячется паук.
Перед помещением в ячейку осы, как правидло, парализуют своих жертв жалом. Уколы жалом большинства сфецид вызывают у жертвы необратимый паралич, только виды родов Dolichurus и Liris – временный паралич. Наибольшее число уколов наносят виды рода Ammophila, производящие уколы жалом почти во все сегменты тела жертвы. Виды ос рода Larra парализуют медведок временно. Через некоторое время оправившаяся от паралича медведка способна передвигаться (она вползает в свою норку, где личинка осы развивается как наружный паразит медведки). Некоторые виды родов Bembix и Mellinus парализуют своих жертв только одним уколом, приэтом жертва или сразу погибает, или впадает в более или менее полный паралич, но иногда может вполне оправиться и двигаться. Некоторые Pemphredoninae не жалят своих жертв, а только малаксируют, что тоже вызывает паралич. Судя по литературным данным, осы некоторых родов, известных в Казахстане, перед помещением добычи в норку ампутируют у нее выступающие части тела, в частности ноги, крылья, усики, но такие особенности поведения у ос региона пока не выявлены.
Малаксация (сдавливание тела с помощью жвал) известна для многих видов. Она выполняет ту же роль, что парализация жалом, т.е. приводит к инактивации жертвы. У многих ос она применяется наряду с парализацией жалом и только у Pemphredonini функционально заменяет ее.
Транспортировка добычи. По типам транспортировки добычи роющих ос можно разделить на 5 групп, имеющих соответственно 1-й, 2-й и 3-й мандибулярный, педальный и абдоминальный способы (по терминологии Г.Эванса (Evans, 1962)). Первый мандибулярный способ, заключающийся в транспортировке жертвы пешком и пятясь, свойственен только одному роду Dolichurus. Второй мандибулярный способ, при котором добыча тоже держится жвалами за усики или тело жертвы, но движение совершается головой вперед (хотя тоже пешком), характерен для видов рода Podalonia, некоторых видов Sphex, Palmodes, Prionyx, Parapsammophila, Ammophila, Ancistromma, Tachysphex. Большинство сфецид транспортируют добычу или по 3-му мандибулярному способу, при котором оса передвигается на лету и держит жертву с помощью жвал (или жвал и одной-двух пар ног), или по педальному способу, при котором добыча удерживается ногами, без участия жала, и движение совершается по воздуху (на лету). 3-й мандибулярный способ характерен для видов родов Chalybion, Sceliphron, многих видов триб Sphecini, Ammophilini, Psenini, Pemphredonini, подсемейств Astatinae и Larrinae, триб Mellinini и Alyssonini.
Педальный способ присущ большинству видов триб Crabronini, Gorytini, Stizini, Bembicini, подсемейства Philanthinae. Абдоминальный способ, или транспортировка на жале, характерен некоторым видам рода Oxybelus и рода Crossocerus.
По способу втаскивания добычи в гнездо всех ос можно разделить на 2 группы. Для первой характерно втаскивание добычи изнутри гнезда (при этом оса, оставив добычу у входа, сама входит в норку и там разворачивается), для второй – вхождение в гнездо вместе с добычей сразу по возвращению с охоты. 1-й способ считается более примитивным, поскольку добыча, оставшаяся без присмотра, может подвергнуться нападению врагов. Этот способ отмечен у Sphecinae, некоторых представителей Pemphredoninae, у Astatinae, Larrini, некоторых Nyssoninae. 2-й способ характерен для Oxybelini, большинства Crabronini, Nyssoninae и Philanthinae.
Принесенную добычу осы помещают в ячейку (иногда после предварительного накапливания в главном ходе). Добыча в ячейке располагается одним из следующих способов: 1) головой внутрь и брюшной поверхностью вниз, 2) головой внутрь, брюшной поверхностью вверх, 3) головой внутрь, на боку, 4) беспорядочно, 5) на спине, головой наружу, 6) на брюшной стороне, головой кнаружи, 7) на боку, головой кнаружи, 8) брюшной стороной к продольной оси ячейки, головой внутрь.
Для большинства сфецид характерно помещение добычи в ячейку в таком положении, в каком она была при транспортировке к гнезду. Наиболее примитивное положение – головой вперед и спиной вверх. Так переносят и помещают в гнездо свою добычу некоторые Sphecinae. Громадное большинство сфецид помещают своих жертв в ячейки головой вперед и спиной вниз. В таком положении они и транспортируют их. У некоторых ос, вероятно, в связи со значительным уменьшением размеров добычи, когда ориентировка ее уже не имеет значения при транспортировке (например, у охотников на тлей), добыча располагается в ячейке беспорядочно.
Однако у немногих роющих ос положение добычи в ячейке существенно меняется и подчиняется строгому контролю со стороны осы (например, у видов рода Lestica и некоторых видов рода Ectemnius). У них добыча упакована в ячейку в виде компактной сосиски, у которой оболочку составляют крылья, а начинку – тела бабочек.
Откладка яйца. Большинство роющих ос откладывают яйцо на тело своих жертв (как правило, головным полюсом) к такому месту, где кожные покровы очень тонкие, так что вылупившаяся личинка сразу может начать питание. Так посупают все осы, у которых охота предшествует яйцекладке. Лишь только те осы, которые охотятся после яйцекладки (Bembecinus, некоторые виды Bembix), кладут яйцо на дно ячейки на специальный постамент в виде камешка или комочка земли.
По К.Ивате (Iwata, 1976), все многообразие положения яйца на теле жертвы у роющих ос сводится к нескольким типам (они названы им по родовым названиям типичных представителей). Эти типы характеризуются следующими признаками: тип Ammophila – яйцо расположено на боку гусеницы в ее средней части, поперек продольной оси тела, прикрепляется передним концом; тип Trypoxylon – яйцо на базальной части брюшка жертвы, сбоку от средней линии, торчит назад, прикрепляется передним концом; тип Larra – маленькое яйцо, прикрепляется позади переднего или среднего тазика сверчка или медведки передним концом; тип Dolichurus – яйцо спереди среднего тазика таракана, прикрепляется передним концом; тип Prionyx – яйцо на сочленовной перепонке позади заднего тазика саранчового (у Stizus прикрепляется задним концом); тип Sphex – яйцо прикрепляется передним концом между передними и средними ногами добычи поперек ее тела (добыча – Orthoptera, Psocoptera, Hemiptera, Diptera, Hymenoptera); тип Crabro – яйцо прикрепляется передним концом к вентральной стороне шеи жертвы (мухи, бабочки и др.) поперек нее подобно полукольцу; тип Gorytes – яйцо располагается на вентральной поверхности груди вдоль тела жертвы (цикадовые) передним концом вперед; тип Cerceris – яйцо на вентральной стороне жертвы (Hymenoptera, Coleoptera) вдоль оси тела или диагонально, обычно передним концом обращено к голове жертвы; тип Pemphredon - яйцо на вентральной или дорзальной поверхности тела тли или листоблошки вдоль ее тела передним концом вперед или назад; тип Bembix – яйцо на боку мухи между грудью и брюшком; тип Bembecinus – яйцо помещается в пустую ячейку вертикально или наклонно обычно на комочек склеенных песчинок, прикрепляется задним концом, так что передний конец свободный; тип Nysson – яйцо прячется под крыло добычи хозяина.
Роющие осы, которые запасают только один экземпляр добычи, откладывают яйцо сразу после помещения его в ячейку. Некоторые осы прикрепляют его даже еще раньше, когда находятся на поверхности земли, перед втаскиванием добычи в норку. Большинство сфецид, запасающих несколько экземпляров добычи, откладывают яйцо или на 1-й экземпляр, или на один из следующих экземпляров до предпоследнего, или на последний экземпляр, или на один из экземпляров внутри массы добычи, но после завершения провиантирования. Наконец, как уже говорилось выше, некоторые роющие осы откладывают яйцо до начала провиантирования – до первой охоты, в пустую ячейку.
Ampulicinae откладывают яйцо на единственный экземпляр добычи до или после втаскивания его в гнездовую полость. У Sceliphronini откладка яйца или завершает все провиантирование, или наступает после притаскивания 1-го экземпляра добычи. Sphecinae откладывают яйцо обычно на 1-й экземпляр (иногда после завершения провиантирования), изредка – на последний. Виды рода Palmodes иногда откладывают в одну ячейку 2 и более яиц. У Pemphredoninae, по предположению К.Иваты (Iwata, 1976), момент откладки яйца наступает после того, как будет принесено не меньше половины нужного запаса провианта, чаще – на последний экземпляр добычи.
Для Astatinae момент откладки яйца точно не установлен, хотя известно, что яйцо прикрепляется к экземпляру добычи, принесенному первым, во всяком случае, лежащим в ячейке дальше всех от входа
(Evans, 1957).
Среди Crabroninae есть такие осы, которые откладывают яйцо на 1-й экземпляр добычи, и такие, которые приступают к откладке яйца после завершения провиантирования или внесения в ячейку нескольких экземпляров.
У Nyssoninae наблюдается варьирование момента откладки яйца по отношению к моменту начала или окончания запасания добычи в ячейку. У Stizini и Bembicini яйцо откладывается на 1-й экземпляр добычи или даже еще в пустую ячейку. У Philanthinae преобладает откладка яйца после окончания заполнения ячейки добычей.
Закрывание гнезда и ячейки. Одна из наиболее характерных черт гнездового поведения ролющих ос, присущих только этому семейству, является временное закрывание входа в гнездо или в ячейку на период охоты. Обычно гнездо закрывается осами, когда в ячейке скопится больше двух экземпляров добычи, но у аммофил ячейка временно закрывается сразу после постройки гнезда, т.е. даже в том случае, когда запасается только одна жертва. Способы временного закрывания у роющих ос довольно разнообразны. Можно выделить несколько типов: 1) закрывание гнезда около входа, 2) закрывание гнезда в глубине главного хода, 3) закрывание только входа в ячейку. Наконец, многие осы не делают временных пробок, оставляя гнездо открытым.
Разнообразны материалы, с помощью которых происходит закрывание, хотя у большинства видов используется материал субстрата. Некоторые осы при закрывании гнезда берут материал из одного определенного места («карьера»), при этом образуется так называемая «фальшивая» норка (например, у Eremochares dives), которая имеет, вероятно, также значение для отвлекания врагов от настоящего гнезда. При закрывании гнезда некоторые осы (например, Podalonia affinis, P. horsuta) использует камешек для утрамбовывания материала.
У Dolichurus (Ampulicinae) о временном закрывании гнезда не может быть и речи, поскольку у них охота предшествует поискам места для устройства ячейки. Временное закрывание появляется и почти всегда имеет место у Sphecinae, для которых характерны устройство гнезда до охоты и заготовка нескольких особей добычи. Среди Pemphredoninae ни одна оса не закрывает гнездо на время охоты, хотя имеются случаи закрывания гнезда изнутри после втаскивания добычи.
Astatinae на время отлучек обычно закрывают вход в гнездо временной пробкой, хотя некоторые виды оставляют его открытым. Не исключено, что у последних временная пробка располагается в глубине норки, и ее не видно с поверхности.
Среди Crabroninae одни осы закрывают вход в гнездо на время охоты, другие не закрывают. У Nyssoninae временное закрывание гнезда тоже имеет место не у всех представителей. У Philanthinae гнездо при отлучках или закрывается снаружи, или остается открытым, или закрывается внутренней пробкой. У некоторых представителей, например, у рода Philanthus это поведение неустойчивое, иногда даже внутри одного вида.
Как показали исследования многих авторов, закрывание гнезда на время охоты предохраняет яйцо, личинку и провиант от врагов и воздействия опасных абиотических факторов (большой сухости воздуха, высокой температуры).
После снабжения ячейки провиантом и откладки яйца оса приступает к окончательному закрыванию ячейки или гнезда в целом (в последнем случае если была запровиантирована единственная или последняя ячейка).
Ampulicinae используют для закрывания гнезда мелкие камешки, частицы земли, кусочки растений и другие частицы минерального и растительного происхождения, которые собирают поблизости от взода.
Среди Sphecinae окончательное закрывание гнезда обычно производится способом, аналогичным тому, каким проводилось временное закрывание, но только делается это более тщательно. Обычно оса сначала втаскивает в гнездо крупные камешки, затем засыпает ход мелкими частицами земли, которые трамбует головой или особым камешком, взятым в челюсти. В завершение работы оса тщательно разравнивает поверхность земли над гнездом и около него и маскирует вход частицами растений, камешками и другим подобным материалом. Многие Sceliphronini используют для закрывания гнезда грязь, смолу и другие пластичные материалы. Для некоторых видов Sphecinae отмечено использование в качестве «орудия труда» камешка в сочетании с вибратором для трамбования земли. Такая черта поведения, как применение «орудия труда», возникла, по Г.Бишофу (Bischoff, 1927), вследствие притаскивания крупных камешков и комочков земли при заполнении норки и трамбования их с помощью головы.
Окончательное закрывание гнезда у Pemphredoninae обычно производится с помощью материалов, приносимых извне, хотя перегородки между ячейками могут изготовляться с применением материала субстрата, взятого со стенок гнезда. Виды рода Passaloecus используют клейкие пластичные вещества, например, смолу хвойных деревьев и влажную глину, иногда добавляя к ним древесные опилки, песчинки, камешки и пр.
Все Astatinae закрывают ячейки и гнездо пробкой из земли. Большинство Crabroninae тоже закрывают норки землей и мелкими камешками. После окончания работы разравнивают поверхность земли над гнездом и маскируют вход. Представители Trypoxylonini запечатывают ячейки и гнездо грязью.
У Crabronini, большинство из которых временно не закрывают гнезда, окончательное закрывание происходит не так тщательно, как у многих других сфецид. Многие используют зля закрывания материал субстрата, но некоторые приносят для этого инородные материалы (смолу, минеральные частицы и пр.).
Среди Nyssoninae наружное окончательное закрывание гнезда проводится очень многими видами. Для них характерно также окончательное разравнивание поверхности над гнездом и тщательная маскировка входа.
Среди Philanthinae окончательное закрывание гнезд отмечено у некоторых видов рода Philanthus. Виды рода Cerceris каждый боковой ход, ведущий к ячейке, заполняют (и плотно утрамбовывают) землей, причем для трамбования, по-видимому, используют пигидий, а главный ход заполняют лишь рыхлой землей, и то не до самого верха. Возможно, настоящего окончательного закрывания церцерисы не производят.
Общий порядок актов гнездового поведения. Забота о потомстве у роющих ос слагается из отдельных актов поведения, важнейшие из которых следующие: изготовление гнезда, охота, транспортировка добычи и втаскивание ее в гнездо, откладка яйца, закрывание ячейки и гнезда в целом. Общий порядок этих актов поведения у роющих ос весьма разнообразный.
В зависимости от этого порядка сфециды Казахстана распределяются на 8 групп. К первой группе относятся осы, чье гнездовое поведение состоит из 2 актов: охоты и откладки яйца. К ней относятся виды рода Larra. Несмотря на простоту, это поведение, по-видимому, является вторично упрощенным, специализированным, обусловленным способностью медведки – добычи осы – после временного паралича вползать в свою норку и тем самым создавать необходимое убежище для потомства осы, играющее роль гнездовой ячейки.
Ко второй группе относятся виды, которые сначала охотятся, потом прячут добычу в случайную естестьвенную полость в субстрате и там откладывают на жертву яйцо. Из видов изучаемого региона к этой группе относятся виды рода Dolichurus, возможно, и некоторые виды рода Liris.
К третьей группе относятся осы, чье гнездовое поведение слагается из охоты, транспортировки и временного прятания добычи на растениях (иногда добыча просто оставляется на земле), постройки гнезда, втаскивания добычи в гнездо, откладки яйца и закрывания гнезда. В этом случае охота предшествует постройке гнезда, гнездо имеет только одну ячейку и запасается только один экземпляр добычи. Такой порядок характерен для неколторых видов Sphex, Podalonia, Liris и Ancistromma.
К четвертой группе принадлежат виды, у которых цикл гнездового поведения начинается с постройки гнезда, а затем уже следуют охота, транспортировка, втаскивание добычи в гнездо, откладка яйца и закрывание гнезда. Здесь гнездо также содержит только одну ячейку, в которой запасается один экземпляр добычи. К этой группе относятся часть видов родов Sphex, Prionyx, Palmodes, Ammophila, Tachytes, Tachysphex, Palarus, Plenoculus и Miscophus.
Пятую группу составляют виды, чье гнездовое поведение состоит из постройки гнезда и ячейки, массового провиантирования, откладки яйца и закрывания ячейки. Момент откладки яйца обычно не строго фиксирован и может происходить на любой стадии провиантирования, но чаще всего – после ее окончания. Некоторые осы могут на время очередной охоты закрывать гнездо временной пробкой. Количество ячеек в гнезде варьирует от 1 до 10-15. К этой группе относится большинство видов Казахстана: некоторые виды Sphex, Palmodes, Prionyx, Ammophila, виды родов Chalybion, Sceliphron, подсемейств Pemphredoninae, Astatinae, Crabroninae, большинство видов подсемейств Nyssoninae и Philanthinae.
Для многих видов триб Sphecini и Ammophilini, принадлежащих к этой группе, свойственно одноячейковое гнездо, для видов подсемейств Pemphredoninae, Astatinae, Crabroninae, Nyssoninae, Philanthinae – многоячейковое.
В шестую группу входят виды, для которых характерно прогрессивное провиантирование и откладка яйца на 1-й экземпляр добычи.
Гнездо одноячейковое. На время охоты всегда закрывается. К этой группе относятся некоторые виды Ammophila (например, A. campestris), Eremochares (например, E. dives) и Bembix (например, B. oculata, B. rostrata).
К седьмой группе относятся виды, которые тоже производят провиантирование по прогрессивному типу, но яйцо откладывают еще в пустую ячейку. Это виды рода Bembecinus и, возможно, некоторые виды рода Bembix.
Восьмую группу составляют клептопаразиты. Это виды родов Nysson, Synnevrus, Brachystegus и Stizoides. Забота о потомстве складывается у них из отыскивания чужих гнезд и откладки в них яйца.
Суточная активность. Большинство роющих ос Казахстана относятся к теплолюбивым видам. Все они – дневные насекомые. В солнечную погоду лет их начинается в утренние часы при температуре воздуха не менее 15-18о С. Началу деятельности обычно предшествуют солнечные «ванны». У большинства видов численность постепенно возрастает до 11-13 часов, затем начинает падать и заканчивается перед заходом солнца. Однако сильный нагрев воздуха и почвы в середине дня в пустынях обусловил у некоторых видов ряд своеобразных изменений. Очень многие виды имеют двувершинную кривую суточной активности вследствие резкого снижения активности в промежутке от 12-13 до 1518 часов. Некоторые виды (например, Dinetus psammophilus – обитатель песков) после наступления жары (в 12-14 часов) зарываются в норки и возобновляют активность только утром следующего дня. Существенные изменения в суточную активность вносят облачность, осадки, сильный ветер,которые снижают, задерживают или прекращают активность ос.
Ночь роющие осы проводят в неактивном состоянии. Большинство на ночлег забираются в свои или чужие (пустые) гнезда или просто закапываются в грунт. Но многие виды подсемейств Sphecinae и Nyssoninae ночуют на растениях, по крайней мере самцы (например, виды родов Stizus, Stizoides, Sphex, Ammophila, Podalonia, Prionyx и др.). Для некоторых видов Sphecinae и Nyssoninae известна ночевка в скоплениях, насчитывающих десятки и даже сотни особей. Наиболее своеобразна ночевка аммофил. Эти осы обычно обхватывают жвалами тонкий стебель какого-нибудь растения (чаще всего из злаковых) и висят, поджав передние ноги и вытянув назад средние и задние.
Копуляция. Встреча особей разных полов происходит у роющих ос в разнообразных местах. Это – или участки гнездования самок (например, у некоторых видов Sphex, Eremochares, Ammophila, Dinetus, Liris, Ancistromma, Tachytes, Tachysphex, Prosopigastra, Plenoculus, Solierella, Miscophus, Oxybelus, Mellinus, Harpactus, Bembecinus, Bembix, Cerceris), или места их питания и охоты (например, у многих видов Sceliphron, Prionyx, Pemphredoninae, Chalybion, Palarus, Trypoxylon, Crabronini, Gorytes, Sphecius, Bembecinus, Bembix, Philanthus, Cerceris). Иногда это специальные участки в местах пролета самок. Самцы караулят самок, сидя на земле, на камешках, комках земли или на растениях (Astata, Dryudella, Tachysphex, Oxybelus, Philanthus) и бросаются на самок при их появлении. Они при этом показывают элементы территориального поведения. Самцы аммофил часто преследуют самок, летая вслед за ними (образуя так называемый «тандем»).
Копуляция, как правило, происходит по обычному для жалящих перепончатокрылых типу: самец садится на самку сверху, хватает ее передними ногами и жвалами и поколачивает усиками ее усики, после чего происходит сближение и сцепление гениталий. Копуляция происходит на земле или растениях и длится от нескольких секунд до нескольких минут. У Dinetus psammophilus самка может перелетать с места на место во время копуляции, при этом самец располагается позади нее («форетическая копуляция») и находится в каталептическом состоянии. Сходно происходит копуляция у Sphecius.
Питание имаго. Взрослые сфециды питаются нектаром цветков растений, выделениями насекомых отряда равнокрылых («медвяная роса», или падь), содержимым зобиков насекомых, являющихся добычей, и гемолимфой жертв. Иногда осы пьют воду.
Преобладающим типом питания у ос Казахстана является высасывание нектара из цветков растений, причем главным образом из цветков с коротким венчиком (у растений семейств Tamaricaceae, Euphorbiaceae, Apiaceae, Fabaceae, Asteraceae и некоторых других). Среди Pemphredoninae и мелких Crabroninae преобладает питание медвяной росой. У многих Sphecinae и Nyssoninae в связи с удлинением у них ротовых органов появляется возможность использовать нектар цветков с глубоким венчиком (например, бобовых), что в условиях нехватки цветущих растений в аридных областях имеет большое значение.
Посещая цветки растений, роющие осы, несомненно, участвуют в опылении их, но размеры этой деятельности и значение в оплодотворении завязей цветков совершенно не изучены.
Развитие. Самцы сфецид, подобно другим группам жалящих перепончатокрылых, развиваются из гаплоидных яиц, самки – из диплоидных. Период между откладкой яиц до выплаживания личинок довольно кратковременный, колеблется от 1 до 3-4 суток и имеет примерно одинаковую продолжительность во всех группах роющих ос. Только для видов Nysson известно, что личинка вылупляется значительно быстрее, чем личинка хозяина, что, очевидно, является приспособлением к инквилинному образу жизни. Молодая личинка Nysson разыскивает яйцо хозяина и уничтожает его, после чего начинает уже питаться запасами добычи в ячейке. Возможно, в данном случае яйцо в момент откладки самкой находится на более поздней стадии эмбрионального развития, чем яйцо хозяина, хотя не исключено, что эмбрион просто развивается быстрее.
Вышедшая из яйца личинка внедряется передним отделом в тело жертвы и начинает питаться ее внутренним содержимым. Личинка поедает сначала наименее важные органы добычи (Мариковский, 1963). Возможно, у личинки преоральное пищеварение, т.е. содержимое тела жертвы под действием пищеварительных ферментов, впрыснутых личинкой осы, разжижжается и затем всасывается личинкой (Olberg, 1959).
Во время питания личинка несколько раз линяет.
Период роста и питания личинки не превышает 2 недели. Закончившая питание личинка плетет кокон, в котором проводит определенный (в зависимости от фенологических особенностей вида) период в состоянии покоя. Эта стадия часто известна под названием предкуколки, или пронимфы. Ее продолжительность очень сильно колеблется, особенно у поливольтинных видов. Зимующая предкуколка может оставаться в коконе до 10-11 месяцев. Известны случаи, когда предкуколка остается зимовать и на 2-й год, что, вероятно, можно рассматривать как страховку для вида от неблагоприятных условий в 1-й год после зимовки (Мариковский, 1963).
Функция кокона, в котором личинка проводит стадию покоя, а затем превращается в куколку, а куколку – в имаго, несомненно, связана с защитой личинки (и куколки) от врагов и неблагоприятных механических и климатических воздействий. Форма и строение кокона у сфецид различаются в значительной степени.
У Dolichurus он удлиненно-овальной формы, прочный, состоит из слоя плотного вещества, выстлан изнутри шелком, снаружи покрыт мелкими частицами земли и разным мусором.
У Sphecinae кокон обычно прочный, имеет веретенообразную форму, как правило, покрыт сверху еще тонкой пергаментовидной оболочкой. У Sceliphron он закруглен на одном конце и усечен на другом, пергаментовидная оболочка отсутствует. В усеченном конце находится мекониум, из-за чего эта часть кокона имеет черную окраску.
У Pemphredoninae кокон обычно овальный. Его стенки состоят из двух слоев шелка. У Diodontus шелковый кокон включает в стенки примесь посторонних частиц: песчинок, различных частичек растительного и минерального происхождения. У видов рода Pemphredon кокон имеет или форму овального, более или менее плотного мешочка или состоит лишь из верхнего (наружного) шелкового плотного колпачка и шелковой перегородки, отделяющей экскременты, внизу. У видов рода Passaloecus кокон имеет вид одного-двух толстых дисков или шелковых перегородок, так как боковые стенки исчезают.
У Astatini кокон тоже более или менее редуцируется (до шелковой выстилки ячейки), так как эти осы гнездятся в очень плотной почве.
Crabroninae, гнездящиеся в земле, характеризуются прочным коконом, стенки которого включают в себя частицы земли. Иногда имеются дыхательные отверстия – поры. Коконы ос рода Trypoxylon имеют тонкие пергаментовидные стенки.
У видов родов Lindenius, Ectemnius и Crabro наблюдаются вкрапления остатков добычи в стенки кокона у его вершины. Для некоторых видов Crabronini характерны коконы, имеющие особые, так называемые «дыхательные» выросты на головном конце.
Личинки Mellinus делают светлый яйцевидный кокон, стенки его пергаментовидные, снаружи покрыты песком и остатками добычи. У Gorytes и других представителей Gorytini коконы тоже удлиненнояйцевидные, довольно твердые, снаружи покрыты остатками добычи; стенки их инкрустированы песчинками. Сходное строение имеют коконы у Stizini. У видов родов Stizus и Bembecinus в средней части кокона обычно имеется несколько респираторных пор. У Bembicini коконы также состоят из шелка и песка и обычно снабжены несколькими дыхательными порами.
Личинки Philanthinae делают кокон из шелка. Он имеет грушевидную или бутылковидную форму; суженным концом, содержащим экскременты, прикрепляется к стенке ячейки. У видов рода Cerceris он обычно покрыт остатками добычи.
Что касается строения кокона у разных представителей сфецид, оно в большой степени зависит от субстрата, в котором находится ячейка. Кокон прочный, многослойный, часто инкрустированный различными инородными частицами у видов, гнездящихся в рыхлой земле, в песке. Зато у видов, гнездящихся в древесине (особенно в готовых ходах ксилофагов или в плотной древесине, а также в полых стеблях растений), он непрочный, тонкостенный, и иногда даже редуцированный до простого колпачка или диска в передней (входной) части ячейки. С изменением субстрата гнездования изменяется и строение кокона. Следовательно, эти изменения происходят по мере освоения представителями семейства различных биотопов (и соответствующих субстратов гнездования).
Моновольтинные осы, за немногим исключением, зимуют в коконе на стадии пронимфы (предкуколки). Минимальная продолжительность этой стадии у поливольтинных видов меньше недели.
После стадии покоя личинка превращается в куколку. Эта стадия у роющих ос довольно непродолжительная: насколько известно, длится от 10 до 60 дней.
Превращение личинки в куколку и затем куколки в имаго происходит у роющих ос в ячейке внутри кокона. Взрослая оса прогрызает стенку кокона (или открывает его крышку) и затем выбирается наружу из гнезда. Коконы в ячейке ориентированы таким образом, что вышедшая оса направлена головой к выходу из ячейки. Выход имаго происходит или почти одновременно, или растянут на более или менее длительный срок. Самцы, как правило, выходят раньше самок.
Интересной особенностью некоторых ос, строящих линейные гнезда (например, Pemphredoninae, Trypoxylonini), является такое расположение ячеек, при котором самцы, выходящие раньше самок, располагаются ближе к выходу из гнезда, чем самки. Этим, вероятно, предотвращается повреждение коконов осами, выходящими из более глубоких ячеек.
Фенология. У подавляющего большинства сфецид Казахстана зимуют личинки (пронимфы), которые весной превращаются в куколок. Вылет имаго происходит во второй половине весны или в начале лета. Наиболее высокая численность роющих ос в южной половине Казахстана в целом наблюдается в конце мая – начале июняч, сдвигаясь к середине июня в северной половине. На стадии имаго зимуют, по всей вероятности, только некоторые виды Liris и Podalonia. Не исключено, однако, что у этих видов могут зимовать и личинки (или куколки). Число поколений в году колеблется от 1 до 2-3 и даже больше. Соответственно различаются периоды летной активности вида. В зависимости от него все виды можно разделить на несколько фенологических групп: 1) весенние и весенне-раннелетние моновольтинные виды, 2) летние моновольтинные, 3) осенние и летне-осенние моновольтинные, 4) весеннелетние, весенне-летне-осенние и летне-осенние поливольтинные или бивольтинные.
Большинство втдов, несомненно, должно быть отнесено к группе поливольтинных видов, т.к. они имеют весьма растянутый период лета имаго, но большая часть – и группе летних и весенне-раннелетних моновольтинных видов. Моновольтинность большинства аридных видов, по всей вероятности, обусловлена необходимостью длительной диапаузы из-за летней жары и засухи. Для некоторых поливольтинных видов (в частности Tachysphex pompiliformis) установлено, что 2-е (и следующее) поколение имаго значительно малочисленнее, чем 1-е. Очевидно, часть личинок не претерпевают дальнейших превращений, а остаются в диапаузе. Это явление, видимо, широко распространено у роющих ос Казахстана, поскольку численность многих поливольтинных видов после весенне-раннелетнего пика значительно снижается. Для некоторых видов рода Tachysphex (T. pompiliformis, T. incertus, T. stachi) установлено, что имаго из части коконов не выходят весной следующего года, а диапауза личинок продолжается еще один год. Причины и механизм такого явления остаются неясными. Можно предположить, что оно обусловлено генетической неоднородностью популяций и биологически выгодно виду как своеобразная страховка на случай особенно неблагоприятных условий следующего года.
Возможно, у некоторых роющих ос, живущих в условиях жарких пустынь, бывает летняя диапауза, обусловленная жарой и засухой, после которой осенью деятельность ос возобновляется, однако такие случаи пока неизвестны.
Продолжительность жизни имаго обычно не превышает 1-2 месяцев, только в случаях зимовки на этой стадии она растягивается до 5-6 и даже 8 месяцев в зависимости от длительности холодного периода года.
Взаимоотношения гнездящихся самок. Большинство роющих ос являются одиночными насекомыми, т.е. они делают гнездо, запасают в нем провизию для потомства в одиночку, независимо от других особей этого же вида. Внутривидовые отношения у них сводятся к копуляции самок и самцов. Иногда наблюдаются даже «драки» между особями, если насекомые сблизятся на критическое расстояние.
Однако среди сфецид можно найти виды с некоторыми элементами колониальной и общественной организаций отношений между особями. Примеры колониального (точнее, псевдоколониального) гнездования самок у роющих ос региона довольно многочисленны. Колонии отмечены для Eremochares dives, Bembix rostrata, Philanthus coronatus, Cerceris arenaria, Bembecinus tridens, Tachytes integer, Oxybelus latro и некоторых других видов. Самки в колониях гнездятся самостоятельно, но их гнезда образуют более или менее плотные скопления.
Общественный образ жизни имеет только у Cerceris rubida. Во всяком случае, он был описан у этого вида в Южной Европе (Grandi, 1961). В одном гнезде обитают несколько самок: основательница гнезда и несколько ее дочерних особей.
Роющие осы в настоящей работе рассматриваются как одно семейство Sphecidae. Такая точка зрения принята в монографии Р.М.Бохарта и А.С.Менке (Bohart and Menke, 1976) и является наиболее распространенной в таксономической литературе о роющих осах.
В состав семейства входят 10 подсемейств: Ampulicinae, Sphecinae, Pemphredoninae, Astatinae, Laphyragoginae, Crabroninae, Entomosericinae, Xenosphecinae, Nyssoninae и Philanthinae. В фауне Казахстана представлено 8 подсемейств (все, кроме Laphyragoginae и Xenosphecinae). Следует отметить, что в соответствии с современными представлениями (Lomholdt, 1985; Alexander, 1922 и др.), подсемейство Crabroninae понимается в широком смысле, т.е. с включением таксонов, входящих и в подсемейство Larrinae системы Р.М.Бохарта и А.С.Менке. Надвидовая таксономическая структура фауны роющих ос Казахстана представлена в таблице 1. В этой системе намеренно опущены подродовые таксоны в связи с тем, что в настоящее время намечается четкая тенденция к вытеснению подродовых названий названиями отдельных видов для обозначения соответствующих групп видов подродового ранга. Нами также не используется категория «подтрибы», поскольку подавляющее большинство триб содержит не более 2 подтриб и практического смысла введение этой категории не имеет. В отличие от системы Р.М.Бохарта и А.С.Менке род Nitela переведен из трибы Miscophini в трибу Trypoxylini в соответствии с аргументацией О.Ломхольдта (Lomholdt, 1985).
Таблица 1 Система надвидовых таксонов семейства роющих ос
(Sphecidae) фауны Казахстана
Подсем. Ampulicinae
Триба Dolichurini Род Dolichurus Latr.
Подсем. Sphecinae
Триба Sceliphronini
Роды Chalybion Dahlb., Sceliphron Klug
Триба Sphecini
Роды Sphex L., Palmodes Kohl, Prionyx Lind.
Триба Ammophilini
Роды Parapsammophila Tasch., Podalonia Fernald, Eremochares Grib., Ammophila Kirby
Подсем. Pemphredoninae
Триба Psenini
Роды Mimesa Shuck., Mimumesa Malloch, Psen Latr.,
Psenulus Kohl
Триба Pemphredonini
Роды Diodontus Curtis, Pemphredon Latr., Passaloecus Shuck., Stigmus Panz., Spilomena Shuck., Ammoplanops Guss., Ammoplanus Gir., Ammoplanellus Guss.
Подсем. Astatinae
Триба Astatini
Роды Diploplectron W.Fox, Astata Latr., Dryudella Spin.
Триба Dinetini Род Dinetus Panz.
Подсем. Crabroninae
Триба Larrini
Роды Larra F., Liris F., Gastrosericus Spin., Larropsis
Patton, Tachytes Panz., Tachysphex Kohl, Parapiagetia Kohl, Prosopigastra A.Costa
Триба Palarini Род Palarus Latr.
Триба Miscophini
Роды Plenoculus W.Fox., Solierella Spin., Miscophus Jur.
Триба Trypoxylini
Роды Pison Jur., Trypoxylon Latr., Nitela Latr.
Триба Oxybelini
Роды Belomicroides Kohl., Belomicrus A.Costa, Oxybelus Latr.
Триба Crabronini
Роды Entomognathus Dahlb., Lindenius Lep. et Br., Rhopalum Steph., Crossocerus Lep. et Br., Crabro F., Ectemnius Dahlb., Lestica Billb.
Подсем. Entomosericinae
Триба Entomosericini Род Entomosericus Dahlb.
Подсем. Nyssoninae
Триба Mellinini Род Mellinus F.
Триба Alyssonini
Роды Alysson Panz., Didineis Wesm.
Триба Nyssonini
Роды Nysson Latr., Synnevrus A.Costa, Brachystegus
A.Costa
Триба Gorytini
Роды Olgia Rad., Argogorytes Ashm., Harpactus Shuck.,
Gorytes Latr., Pseudoplisus Ashm., Lestiphorus Lep.,
Oryttus Spin., Sphecius Dahlb., Ammatomus A.Costa, Kohlia Handl., Psammaecius Lep., Hoplisoides Grib.
Триба Stizini
Роды Stizus Latr., Stizoides G.-Men., Bembecinus
A.Costa
Триба Bembicini
Род Bembix F.
Подсем. Philanthinae
Триба Eremiaspheciini
Род Eremiasphecium Kohl
Триба Philanthini
Род Philanthus F.
Триба Aphilanthopsini Род Philanthinus Beaum.
Триба Pseudoscoliini
Род Pseudoscolia Rad. Триба Cercerini Род Cerceris Latr.
Как известно, естественная система того или иного таксона представляет собой отражение филогенеза группы. Существует довольно обширная библиография по вопросам филогенеза сфецид. Р.М.Бохарт и А.С.Менке (Bohart and Menke, 1976) обобщают и критически анализируют филогенетические воззрения предшественников. Они приводят дендрограмму, показывающую предполагаемый филогенез семейства Sphecidae (рис. 54). Как видно из дендрограммы, Ampulicinae, Sphecinae, Pemphredoninae и Astatinae образуют изолированные ветви, тогда как остальные подсемейства группируются около 2 главных стволов: ниссонинного и крабронинно-филантинного. В целом придерживаясь этой схемы, все же следует иметь в виду, что она не может считаться лостаточно устоявшейся и предстоят большие дополнительные исследования, возможно, на новой методической основе в этом направлении. Это достаточно убедительно показано в работе В.А.Александера (Alexander, 1992). Согласно результатам кладистического анализа семейства роющих ос, осуществленного В.А.Александером, установлено, что монофилетическими являются группы : а) Ampulicini + Dolichurini, b) Sceliphronini + (Sphecini + Ammophilini), c) (Aphilanthopsini + Philanthini + Cercerini + Pseudoscoliini), d) Nyssonini + Gorytini + Stizini + Bembicini, возможно, также e) Crabronini + Oxybelini. Недостаточно ясны связи этих групп друг с другом и с остальными группами роющих ос (Entomosericini, Dinetini, Astatini, Eremiaspheciini, Alyssonini, Pemphredonini, Psenini, Palarini, Mellinini, Miscophini, Larrini, Trypoxylini). Кстати, определенно монофилетической группой оказались пчелиные, которые произошли от роющих ос. Однако неясно, к какому подсемейству ос наиболее близки предки пчелиных: к Nyssoninae, Philanthinae, Pemphredoninae или Laphyragoginae.
Что касается происхождения семейства (или надсемейства) роющих ос, то достаточно убедительных доказательств их связи с определенными группами нет. Скорее всего, как считает А.П.Расницын (1980), на роль предков сфекоидов могут претендовать раннемеловые Baissodidae, обладающие обширным набором примитивных признаков, встречающихся у сфекоидов. Палеонтологическая история роющих ос начинается с раннего мела (Расницын, 1980).
Сфекоидные осы филогенетически и систематически наиболее близки к помпилоидным осам. Помпилоиды в целом гораздо примитивнее сфекоидов. Наиболее характерные отличия состоят в следующем: переднеспинка Sphecoidea почти всегда не достигает тегул; ее задний край почти прямой, только перед плечевыми буграми выемчатый; у Pompiloidea переднеспинка всегда достигает тегул, задний край дугообразно изогнутый, но перед плечевыми буграми невыемчатый. Средние тазики Pompiloidea сомкнуты, у Sphecoidea расставленные. Метапостнотум Pompiloidea короткий, у Sphecoidea посредине сильно удлинен.
Сфекоидные осы, как упоминалось, близки к пчелиным. Отношения этих групп, как пишет А.П.Расницын (1980), соответствует отношениям между предком и потомком. В литературе распространено разделение пчелиных и роющих ос на 2 надсемейства: Sphecoidea (с 1 семейством Sphecidae) и Apoidea с несколькими семействами. Различия между этими надсемействами незначительны: 1-й членик задних ног пчелиных уплощен, расширен и обладает щеточкой волосков, у роющих ос он цилиндрический и без щеточки. В отличие от густоволосистого тела большинства пчелиных тело роющих ос голое или слабоволосистое и волоски простые, а не перистые. В литературе имеется и другое мнение: об объединении пчелиных и роющих ос в надсемейство Sphecoidea (Расницынб 1980; Brothers, 1975; Lomholdt, 1982).
Использована терминология Р.М.Бохарта и А.С.Менке (Bohart and Menke, 1976). Номенклатура жилок и ячеек крыльев показана на рис. 55.
1 (2). Усиковые ямки прикрыты нависающим платформообразным выступом. Нотаули хорошо развитые, доходят до заднего края скутума. Коготки с зубцами на внутреннем крае или на конце раздвоенные. Средние голени с 2 шпорами. Югальная лопасть заднего крылы маленькая (меньше половины анальной области). Брюшко висячее. Жилка M переднего крыла отходит дистальнее жилки cua. Конец маргинальной ячейки лежит на крае крыла. (Триба Dolichurini)…………………………………….………Dolichurus Latr.
2 (1). Усиковые ямки не прикрыты выступом. Нотаули, если имеются, не доходят до заднего края скутума. Признаки: наличие зубцов на коготках, 2 шпоры на средних голенях и маленькая югальная лопасть – никогда не сочетаются (не бывают вместе). Брюшко висячее или стебельчатое. Жилка М переднего крыла отходит или дистальнее cu-a, или проксимальнее, или интерстициально. Конец маргинальной ячейки лежит на крае крыла или отодвинут от него.
3 (28). Задние крылья с большой югальной лопастью, занимающей по длине более половины анальной области. Брюшко с длинным цилиндрическим стебельком, состоящим из 1-го стернума (у Ammophila стебелек состоит из 2 сегментов: 1-го стернума и 1-го тергума, причлененных последовательно друг к другу). Коготки обычно с зубцами на внутреннем крае. (Подсем. Sphecinae)
4 (9). Лапки с плантулами. Коготки с 1 зубцом в середине внутреннего края (иногда у основания) или без зубца. Тело металлически синее или черное с желтым. (Триба Sceliphronini)
5 (8). Промежуточный сегмент с U-образным срединным полем, ограниченным, по крайней мере сзади, бороздкой или швом.
6 (7). Обе возвратные жилки впадают во 2-ю субмаргинальную ячейку. Дыхальцевая борозда отсутствует. Тело обычно с желтыми пятнами, без металлической окраски. Самки без гребня (ряда длинных щетинок) на передних лапках. ………………………Sceliphron Klug
7 (6). 2-я возвратная жилка впадает в 3-ю субмаргинальную ячейку или интерстициально. Дыхальцевая борозда имеется. Тело без желтых
пятен, часто металлически синее или зеленое. Самки с гребнем на передних ногах. ………………………………………...Chlorion Latr.
8 (5). Промежуточный сегмент без срединного поля, самое большее со срединной бороздой, оканчивающейся ямкой. Обе возвратные жилки впадают во 2-ю субмаргинальную ячейку. Самки без гребня щетинок на передних лапках. Тело металлически синее………….
………………………………………………………..Chalybion Dahlb.
9 (4). Лапки без плантул (если с плантулами, то тело черное с красным или целиком черное). Коготки простые или с 1-5 зубцами на внутреннем крае.
10 (19). Коготки с 2 и более зубцами, 2-я возвратная жилка обычно впадает в 3-ю субмаргинальную ячейку, если во 2-ю, то коготки, по крайней мере, с 3 зубцами или 5-й членик задней лапки с очень широкими щетинками на конце снизу. (Триба Sphecini)
11 (14). Базальная сторона 2-й субмаргинальной ячейки равна или короче, чем передняя сторона. Внутренние края глаз прямые внизу, слегка широко вогнутые вверху. Внутренняя шпора задней голени с нежными густыми щетинками по всей длине. Дыхальцевая борозда на промежуточном сегменте обычно имеется. (Подтриба Sphecina)
12 (13). Дыхальцевая борозда на промежуточном сегменте имеется, полная. Стебелек брюшка короче 2-4-го члеников задних лапок, взятых вместе. …………………………………………………..Sphex L.
13 (12). Дыхальцевая борозда более или менее редуцированная. Стебелек брюшка по длине равен или длиннее, чем 2-4-й членики задних лапок вместе. ………………………………Isodontia Patton
14 (11). Базальная сторона 2-й субмаргинальной ячейки длиннее, чем передняя. Внутренние края глаз прямые. Внутренняя шпора задней голени с редкими крепкими щетинками, по крайней мере, в середине. Дыхальцевая борозда отсутствует. (Подтриба
Prionyxina)
15 (16). Свободный край наличника самки цельный или со срединной вырезкой. Коготок задних лапок с 2-5 зубцами на внутреннем крае. Жгутик усиков самца часто с плакоидами на 3-4-м члениках.
…………………………………………………………….Prionyx Lind.
16 (15). Наличник самки с усеченной или слегка выемчатой срединной лопастью, ограниченной по бокам вырезкой или угловытым изгибом. Коготок задних лапок с 2 базальными зубцами на внутреннем крае. Жгутик самца без плакоидов.
17 (18). Эпистернальная борозда тянется вентрально почти до передневентрального края плеврона. Самка с хорошо развитым гребнем щетинок на передних лапках. ……………….……….Palmodes Kohl
18 (17). Эпистернальная борозда оканчивается на уровне скробальной ямки. Самка без гребня щетинок на передних лапках. ……………………………………………….……Chilosphex Menke
19 (10). Лщготки обычно простые или с 1 зубцом (если с 2 зубцами, то вершинные щетинки 5-го членика задних лапок узкие, разделенные промежутками, в 3 и более раз превышающими ширину щетинок). 2-я возвратная жилка впадает во 2-ю субмаргинальную ячейку. (Триба Ammophilini)
20 (23). Коготки простые; если с 1 зубцом, то ротовые части очень длинные: галеа в развернутом состоянии достигает основания стипеса.
21 (22). Вершина 1-го стернума брюшка достигает и часто покрывает основание 2-го. Дыхальце 1-го тергума расположено перед вершиной 1-го стернума. …………………………………Podalonia Fernald
22 (21). Вершина 1-го стернума брюшка не достигает основания 2-го, промежуточная часть обычно длинная и состоит из мембраны и связки (лигамента). Дыхальце 1-го тергума расположено на уровне вершины 1-го стернума или позади него. …………………………
……………………………………………….………Podalonia Fernald
23 (20). Коготки с 1 или 2 базальными зубцами на внутреннем крае. Галеа в развернутом состоянии никогда не тянется за середину стипеса и обычно короче.
24 (25). Стебельковая ямка почти полностью окружена стернумом промежуточного сегмента. Внутренние края глаз самки сильно сближаются книзу. ………………………………...Eremochares Grib.
25 (24). Стебельковая ямка окаймлена вентрально Т-образным стернумом промежуточного сегмента. Внутренние края глаз самки параллельные или слегка сближающиеся книзу или кверху.
26 (27). 1-й членик передней лапки самки сильно асимметричный, его наружная вершина удлинена, сильно выступающая. Передний край наличника самца закругленный или усеченный. Лапки обычно с плантулами. ……………..……………..Parapsammophila Tasch.
27 (26). 1-й членик передней лапки самки почти симметричный, его наружная вершина только слегка выступающая. Наличник самца впереди треугольный. Плантулы отсутствуют……………………..
………..…………………..…………………...Hoplammophila Beaum.
28 (3). Югальная лопасть заднего крыла короче половины длины анальной области; если длиннее, то брюшко висячее или со стебельком, состоящим из тергума и стернума. Коготки иногда с зубцами.
29 (54). Средние голени с 1 шпорой, брюшко со стебельком, состоящим только из стернума, если по-другому, то птеростигма, по крайней мере, равна дискоидальной ячейке.
30 (37). Переднее крыло с 3 субмаргинальными ячейками. Усиковые ямки расположены ясно выше лобно-наличникового шва. (триба Psenini)
31 (32). Жилка М заднего крыла отходит от поперечной жилки cu-a или дистальнее нее. Лобный киль между основаниями усиков с особым выступом и соединен внизу с поперечным килем, по крайней
мере, у самцов. Передняя лапка самки без гребня щетинок. (Подтриба Psenulina) …………………………………...Psenulus Kohl
32 (31). Жилка М заднего крыла отходит ясно перед жилкой cu-a. (Подтриба Psenina)
33 (34). Омаулюс изогнут полукругло вперед к переднегруди. Скробальная борозда неглубокая. Гипоэпимеральное поле слабо выраженное, обычно сильнее пунктированное или морщинистое, чем средняя часть мезоплеврона. Лобный киль обычно неполный. (Затылочный киль везде одинаковой высоты, достигает или почти достигает гипостомального. Пронотальный воротничок с поперечным килем по всей ширине. Пигидиальное поле самки ограничено
мощными килями по бокам)…………….…………….Mimesa Shuck
34 (33). Омаулюс, сливаясь с ацетабулярным килем, тянется до средневентральной линии или оканчивается, как только переходит на вентральную поверхность груди или поворачивает назад. Скробальная борозда глубокая. Гипоэпимеральное поле обычно гладкое и сильно выпуклое. Лобный киль обычно тянется от среднего глазка до межусикового промежутка.
35 (36). Омаулюс продолжен ацетабулярным килем до середины вентральной поверхности груди. Стебелек брюшка сверху с продольным килем или сзади с бороздой (вершина наличника тонкая, без вздутия)…………………………………………..…Mimumesa Shuck.
36 (35). Омаулюс не продолжен ацетабулярным килем до середины груди. Стебелек без продольного киля, по крайней мере, у самцов (самки с наличником, направленным вниз и прикрывающим частично верхнюю губу, без выступа на щеках. 4-й и 3-й стернумы самца часто с апикальной бахромой волосков. …………...Psen Latr. 37 (30). Передние крылья не более, чем с 2 субмаргинальными ячейками. Усиковые ямки обычно расположены сразу выше лобноналичникового шва. (Триба Pemphredonini)
38 (43). Передние крылья с 2 возвратными жилками и 3 дискоидальными ячейками. Птеростигма маленькая или умеренных размеров. (Подтриба Pemphredonina)
39 (40). Эпистернальная борозда хорошо развита, тянется от субалярной ямки до гиперстернаулюса и ниже. Гиперстернаулюс горизонтальный. Верхняя губа с цельной вершиной, обычно дугообразно выступающая. Мандибулы с 2-3 зубцами. Самка без пигидиального поля. Задняя голень без ряда шипов вдоль заднего края. (Внутренние края глаз почти параллельные. Щеки без длинных отстоящих щетинок снизу. Омаулюс обычно отсутствует. Средние членики жгутика длиннее своей ширины). ………………..Passaloecus Shuck.
40 (39). Эпистернальная борозда неполная, нечеткая. Гиперстернаулюс наклонный. Верхняя губа выемчатая или цельная. Мандибулы с 26 зубцами. Самка с пигидиальным полем. Задняя голень часто с рядом шипов вдоль заднего края.
41 (42). Стебелек брюшка длиннее своей ширины. Верхняя губа с цельной вершиной (иногда слегка выемчатой). ……….Pemphredon Latr.
42 (41). Cтебелек шире своей длины (см. сверху). Верхняя губа выемчатая. ………………………………………………..……Diodontus Curt. 43 (38). Передние крылья с 1 возвратной жилкой и 2 дискоидальными ячейками, редко с одной. Птеростигма большая.
44 (47). Передние крылья с удлиненной маргинальной ячейкой, которая больше, чем птеростигма, и закрытая апикально. Воротничок переднеспинки с полным поперечным килем. Омаулюс имеется или отсутствует. (Подтриба Stigmina)
45 (46). Cтебелек брюшка значительно длиннее своей ширины. (Ацетабулярный киль развит и соединен с омаулюсом. Субомаулюс имеется. Мандибулы самки с 3 зубцами. Наличник самца с густым серебристым прилегающим опушением)………………..Stigmus Panz.
46 (45). Cтебелек брюшка не выраженный или не длиннее своей ширины. (Переднее крыло с 2 субмаргинальными ячейками. Затылочный киль отсутствует. Жилка М заднего крыла отходит от передней вершины субмедиальной ячейки)………….…Spilomena Shuck.
47 (44). Переднее крыло с короткой маргинальной ячейкой, которая по величине не превышает птеростигму и на вершине часто открытая. Воротничок переднеспинки без полного поперечного киля. Отмаулюс отсутствует. (Подтриба Ammoplanina)
48 (49). Маргинальная ячейка переднего крыла полная, но не доходит до переднего края крыла. Субмаргинальных ячеек две, вторая из них иногда стебельчатая. ……………………………...Protostigmus Turn.
49 (48). Маргинальная ячейка переднего крыла неполная или полная и доходит до переднего края крыла.
50 (51). Птеростигма широкая, заостренная на дистальной вершине. Маргинальная ячейка переднего крыла замкнутая. Дистальные стороны маргинальной, субмаргинальной и 1-й дискоидальной ячеек образуют почти прямую линию. ………...Ammoplanops Guss.
51 (50). Птеростигма овальная, округленная дистально. Маргинальная ячейка переднего крыла замкнутая или открытая. Дистальные сто-
роны маргинальной, субмаргинальной и 1-й дискоидальной ячеек не образуют прямую линию.
52 (53). Маргинальная ячейка переднего крыла замкнутая………………..
...…………………………………………………..Ammoplanus Giraud
53 (52). Маргинальная ячейка переднего крыла незамкнутая: отсутствует жилка, расположенная дистальнее птеростигмы…………………… …………………………………………………...Ammoplanellus Guss.
54 (29). Средние голени с 2 шпорами; если с одной, то брюшко без стернального стебелька и птеростигма меньше, чем 1-я дискоидальная ячейка.
55 (62). Предптеростигмальный отдел 1-й субмаргинальной ячейки переднего крыла значительно больше, чем половина всей длины ячейки.
56 (61). Глазки нормальные. (Триба Stizini)
57 (58). Промежуточный сегмент сзади по бокам с килями, обычно выемчатыми. Эпистернально-скробальная борозда отсутствует. Жилка М заднего крыла отходит вблизи жилки cu-a. 9-й членик жгутика самцов с шиповидным апикальным выступом…………………..
……..………….…………………………………..Bembecinus A.Costa
58 (57). Промежуточный сегмент сзади по бокам без килей. Эпистернально-скробальная борозда имеется. Жилка М заднего крыла отходит далеко впереди жилки cu-a. 9-й членик жгутика самцов без шиповидного апикального выступа.
59 (60). Глаза с почти параллельными или слегка сближающимися книзу внутренними краями. Мандибулы с субапикальным зубцом. Лоб с небольшим низким выпуклым двойным возвышением ниже среднего глазка. .………………………………………………..Stizus Latr.
60 (59). Глаза сильно сближаются книзу. Мандибулы простые. Лоб с бугровидным выступом ниже среднего глазка………………….. ……………………………………………………Stizoides Guer.-Men.
61 (56). Глазки более или менее деформированные и редуцированные. (Триба Bembicini). Число члеников в пальпах 4 и 2 или 3 и 1. Средний глазок обычно редуцирован до узкого поперечного рубца. …………………………………………………………....Bembix F.
62 (55). Длина предптеростигмального отрезка 1-й субмаргинальной ячейки переднего крыла немного больше, чем половина всей длины ячейки.
63 (92). Средняя голень с 2 апикальными шпорами. Скутум с косым килем в заднебоковых углах. Воротничок переднеспинки короткий и поперечный. Югальная лопасть заднего крыла отсутствует или короче половины длины анальной области. Мандибулы без выемки на наружно-вентральном крае.
64 (69). 1-й стернум с парой килей у основания, но без срединного киля. Скутум со срединной бороздой и косым скутальным килем в задних углах. Передние крылья с 3 субмаргинальными ячейками, из которых 2-я стебельчатая. (Триба Nyssonini)
65 (66). Задняя голень неправильно или пильчато-зубчатая сзади. Самцы с выступающими пучками волосков на 2-5-м стернумах………….
…..……………………………………………….Brachystegus A.Costa
66 (65). Задняя голень только с волосками и щетинками. Самцы без пучков волосков на стернумах.
67 (68). Задние края тергумов не утолщенные и не двойные …………….
….………………………………………………………….Nysson Latr.
68 (67). Задние края тергумов утолщенные и двойные…………………
………...…………………………………………….Synnevrus A.Costa
69 (64). 1-й стергум без пары килей у основания или со срединным килем. Скутум с околосрединными линиями, но без срединной борозды.
70 (91). 1-й стернум со срединным килем и иногда с другими килями. Косой скутальный киль часто имеется. (Триба Gorytini)
71 (76). Жилка М заднего крыла отходит дистальнее жилки cu-a не ближе, чем на диаметр среднего глазка.
72 (73). Отогнутый боковой край скутума прерван напротив задней части тегулы косым килем, который ограничивает маленькую, часто наклонную область. Задняя сторона 2-й субмаргинальной ячейки (между возвратными жилками) не длиннее четверти задней стороны 1-й субмаргинальной ячейки. Внутренние края глаз почти параллельные. ……………………………………….Harpactus Shuck.
73 (72). Боковой край скутума отогнут вверх и не прерван напротив тегулы. Задняя сторона 2-й субмаргинальной ячейки длиннее четверти задней стороны 1-й субмаргинальной ячейки. Внутренние края глаз параллельные или сближающиеся книзу.
74 (75). Лоб на уровне среднего глазка уже, чем недалеко ниже него. Омаулюс продолжается вентрально в виде ацетабулярного киля. 8й стернум самцов мечеобразный, часто торчит наружу. Жилка М заднего крыла отходит от жилки M+Cu под тупым углом………………………………………………….Argogorytes Ashm.
75 (74). Лоб на уровне среднего глазка шире, чем ниже него. Омаулюс продолжается вентрально только как тонкий шов и оканчивается перед срединной линией. 8-й стернум самцов скрытый, на вершине широко закругленный, не мечеобразный. 3-5-й стернумы самца с бахромой волосков. ………………………………………Olgia Rad.
76 (71). Жилка М заднего крыла отходит перед (базальнее) жилкой cu-a или интерстициально, если дистальнее, то не дальше, чем на диаметр переднего глазка.
77 (78). Самка с 2 щетинками на 1-м членики передних лапок перед вершиной. Последние 4 членика усика самца без особых модификаций. Дыхальцевая борозда имеется. Срединное поле промежуточного сегмента со складчатой скульптурой, по крайней мере, впереди. …………………………………………………..Gorytes Latr.
78 (77). Сочетание признаков иное.
79 (84). Мезоплеврон без стернаулюса.
80 (81). Омаулюс отсутствует. Внутренние края глаз сильно сближаются книзу. Стернумы самца без бахромы волосков. 1-й сегмент брюшка иногда в виде стебелька. ………………...….Ammatomus A.Costa 81 (80). Омаулюс имеется.
82 (83). Глазки отчасти редуцированные (уплощенные).Эпистернальная борозда продолжается вниз почти вертикально до омаулюса…….
………………………………………………………..…..Kohlia Handl.
83 (82). Глазки нормальные. Эпистернальная борозда изгибается назад и сливается со скробальной бороздой. (Наличник сбоку в виде прямого или тупого угла. Мандибулы двухзубцовые)………………….
………………………………………………………….Sphecius Dahlb.
84 (79). Мезоплеврон со стернаулюсом.
85 (86). 1-й сегмент брюшка у основания в виде стебелька, его тергум сильно расширенный у вершины (вздутый). (Жилка cu-a заднего крыла слегка изогнута. Мезоплеврон с редкой мелкой пунктировкой. 8-й стернум самца на вершине в виде 2 шипов)…………… …………………………………………………...…….Lestiphorus Lep.
86 (85). 1-й сегмент иногда суженный, но тергум без сильного расширения у вершины (не вздутый).
87 (90). Ацетабулярный киль имеется, четкий и полный, если отсутствует, субомаулюс имеется и тянется на вентральную сторону; 8-й стернум самца мечеобразный. Стернумы самца без апикальной бахромы волосков. (Промежуточный сегмент и скутум обычно пунктированные. 5-й и 6-й стернумы самца со скрытыми пучками волосков).
88 (89). Ацетабулярный киль имеется. Субомаулюс не продолжается на вентральную сторону груди. ……………………….Hoplisoides Grib.
89 (88). Ацетабулярный киль отсутствует. Субомаулюс продолжается на вентральную сторону груди. ………………...……Psammaecius Lep.
90 (87). Ацетабулярный киль отсутствует или неполный или нечеткий. Субомаулюс короткий. 8-й стернум самца на вершине двухзубцовый. (Дыхальцевая борозда имеется. Срединное поле промежуточ-
ного сегмента продольно морщинистое. Эпистернальная и скробальная борозды в виде единой дугообразной или угловато изогнутой линии. Аролиум на передних ногах самки значительно больше, чем на остальных ногах. Жилка М заднего крыла отходит от жилки cu-a или вблизи нее. 3-5-й стернумы самца без апикальной бахромы.)…………………………………………….Oryttus Spin.
91 (70). 1-й стернум у основания без килей. Косой скутальный киль отсутствует. (Брюшко стебельчатое. Жилка М заднего крыла отходит перед жилкой cu-a.) (Триба Mellinini) ………………Mellinus F.
92 (63). Средняя голень с 1 апикальной шпорой или без шпор. Косой скутальный киль отсутствует. Если средняя голень с 2 шпорами, то или воротничок длинный и закругленный, или югальная лопасть длиннее половины анальной области, или мандибулы с вырезкой на наружно-вентральном крае.
93 (110). Задние глазки деформированные: овальные, эллиптические или запятовидные. Наличник полностью не разделен продольными линиями на 3 части. (Триба Larrini)
94 (95). Переднее крыло с 2 субмаргинальными ячейками……………. ...…………………………………………………….Gastrosericus Spin.
95 (94). Переднее крыло с 3 субмаргинальными ячейками.
96 (97). Стебельковая ямка изолирована от метакоксального углубления парой темных склеритов промежуточного сегмента. Внутренний коготок лапок самца часто сильно редуцированный. Мандибулярные ямки закрытые…………….. ……………..…..Parapiagetia Kohl
97 (96). Стебельково-метакоксальное углубление полностью мембранное. Внутренний коготок лапки самца не редуцированный. Мандибулярные ямки открытые.
98 (101). Глазки очень маленькие, узкие, эллиптические, их длинные оси лежат на прямой линии, проведенной от одного глаза к другому. Лоб сразу ниже среднего глазка с поперечным вздутием, идущим от одного глаза к другому и прерванным срединной лобной линией, и линейными вздутиями вдоль внутренних краев глаз. Мандибулы простые.
99 (100). Последний членик лапок самки равномерно дугообразно изогнутый (см. сбоку), с расходящимися почти до вершины боковыми сторонами (см. сверху), без густого опушения на вентральной поверхности. Коготки не хватательные и без зубца. Бока промежуточного сегмента густо пунктированные, блестящие и без килей. Воротничок переднеспинки плоский или дугообразно выпуклый (см. спереди). Пигидиальное поле гладкое и блестящее, вершина без поперечного ряда щетинок. Бока промежуточного сегмента
самца густо пунктированные, блестящие и с несколькими килями. Мандибулы без внутренних зубцов. …………….…Larra F.
100 (99). Последний членик лапок самки угловато изогнутый (см. сбоку), с параллельными в апикальной половине боковыми краями, с пятном густых волосков на вентральной поверхности. Коготки хватательные, иногда с зубцом. Бока промежуточного сегмента обычно матовые, иногда блестящие, но не пунктированные, иногда с диагональными складками. Воротничок переднеспинки клинообразный (см. спереди). Пигидиальное поле обычно покрытое щетинками и часто с поперечным рядом крепких щетинок на вершине. Бока промежуточного сегмента самца матовые и непунктированные, если блестящие, то с редкими мелкими точками. Мандибулы часто с зубцами. ……………………………………………….Liris F.
101 (98). Глазки овальные, запятовидные или удлиненно-клиновидные, их длинные оси ориентированы косо (не перпендикулярно продольной срединной линии головы). Лоб без вздутий, описанных выше.
102 (103). Глазки очень длинные, запятовидные, их длинные оси образуют угол 700 или меньше. Промежуток между средним глазком и концом «хвоста» одного из передних глазков меньше длины переднего глазка. Оба пола с пигидиальным полем, обычно покрытым густыми золотистыми или серебристыми щетинками.
…………………………………………………………..Tachytes Panz.
103 (102). Глазки овальные или продолговатые, если запятовидные или удлиненно-клиновидные, то их длинные оси образуют угол 800 и более. Промежуток между средним глазком и нижним концом переднего глазка равен или больше, чем длина переднего глазка. Пигидиальное поле обычно имеется у самки, но голое или в редких щетинках. Самцы обычно без пигидиального поля.
104 (107). 2-й тергум без бокового киля.
105 (106). 1-2-й членики передних лапок самки с многочисленными густыми длинными нежными щетинками. Вершина 8-го стернума самца выемчатая. Лоб сразу выше усиковых ямок с парой бугров, обычно гладких и блестящих. Глазки овальные или продолговатые, редко удлиненно-клиновидные. …………Tachysphex Kohl
106 (105). 1-й членик передних лапок самки с 4-5 крепкими шипообразными щетинками, разделенными широкими промежутками. 2-й членик только с 2 шипами на вершине снаружи. Вершина 8-го стернума самца закругленная или усеченная. Лоб без пары бугров над усиковыми ямками. Глазки запятовидные. Воротничок переднеспинки с острым краем, тесно прилегающим к скутуму. 3-я субмаргинальная ячейка удлиненная. Скапус короткий, широкий. Затылочный киль соединяется с гипостомальным………………….. ...…………………………………………………..Ancistromma W.Fox
107 (104). 2-й тергум с боковыми килями, по крайней мере, в базальной половине.
108 (109). Гребень на передних лапках самки состоит из длинных тонких щетинок. Передние бедра самцов простые. Оба пола с выступающим лобным бугром, обычно гладким и блестящим. Глаза у самцов иногда соприкасаются вверху. …….....Prosopigastra A.Costa
109 (108). Самки без гребня щетинок на передних ногах. Передние бедра самцов с вырезкой сзади у основания. Лоб без выступающего бугра. Глаза самцов не соприкасаются вверху………………………… ………...………………………………………Holotachysphex Beaum. 110 (93). Задние глазки нормальные или наличник разделен продольными линиями на 3 части.
111 (114). Омаулюс имеется. Жилка М заднего крыла отходит перед жилкой cu-a. Эпистернальная борозда отсутствует. (Триба Alyssonini)
112 (113). Жилка М переднего крыла отходит позади жилки cu-a, если перед ней, то очень близко. Последний членик усиков самца изогнутый, но вершина его не загнута навстречу выступу предпоследнего членика. Метаплеврон по длине вдвое короче своей высоты. 2-й тергум с парой бледных пятен. …………….Alysson Panz.
113 (112). Жилка М переднего крыла отходит перед жилкой cu-a не ближе, чем на длину последней. Последний членик усиков сильно изогнутый и вершина его загнута навстречу выступу 12-го членика. Метаплеврон значительно уже своей высоты. 2-й тергум без бледных пятен. ……………………………………..…Didineis Wesm.
114 (111). Омаулюс отсутствует. Жилка М заднего крыла отходит дистальнее жилки cu-a (или интерстициально). Эпистернальная борозда имеется.
115 (128). Средние голени с 2 апикальными шпорами, если без шпор, то югальная лопасть заднего крыла маленькая и мандибулы с вырезкой на наружно-вентральном крае. (Подсем. Astatinae)
116 (127). Передние крылья с 3 субмаргинальными ячейками. Югальная лопасть заднего крылы длиннее половины длины анальной области. Жилка A2 имеется. Мандибулы без вырезки на наружновентральном крае. Средняя голень с 2 апикальными шпорами обоих полах. (Триба Astatini)
117 (122). Самцы. Усики 13-члениковые. Брюшко с 7 видимыми снаружи сегментами.
118 (119). Глаза вверху не соприкасаются. Только 2-я возвратная жилка впадает во 2-ю субмаргинальную ячейку. Югальная вырезка глубокая. …………………………………………….Diploplectron W.Fox
119 (118). Глаза вверху соприкасаются.
120 (121). Малярный промежуток (щека) длиннее, чем диаметр среднего глазка. 2-я субмаргинальная ячейка короче, чем 1-я на медиальной жилке. 1-й тергум уже, чем 2-й. ……………………..Dryudella Spin.
121 (120). Малярный промежуток короткий, основание мандибулы почти касается нижнего края глаза. 2-я субмаргинальная ячейка по медиальной жилке длиннее, чем 1-я. 1-й тергум по ширине равен
2-му. ………………………………………………………..Astata Latr.
122 (117). Самки. Усики 12-члениковые. Брюшко с 6 видимыми снаружи тергумами.
123 (124). Пигидиальное поле окаймлено крепкими изогнутыми краевыми щетинками. 2-я субмаргинальная ячейка равна 1-й или длиннее. ……………………………………………………..…..Astata Latr.
124 (123). Пигидиальное поле не окаймлено крепкими щетинками. 2-я субмаргинальная ячейка короткая, не больше 0,75 длины 1-й.
125 (126). 1-я возвратная жилка впадает в 1-ю субмаргинальную ячейку или интерстициальная. Срединная лопасть наличника усеченная и срединное поле промежуточного сегмента с грубой скульптурой или наличник треугольный и срединное поле промежуточного сегмента с зернистой скульптурой. ………..…..Diploplectron W.Fox 126 (125). Сочетание указанных выше признаков иное. …………………..
…………………………………………………………..Dryudella Spin.
127 (116). Передние крылья с 2 субмаргинальными ячейками. Югальная лопасть заднего крыла меньше половины длины анальной области. 2-я анальная жилка отсутствует. Мандибулы обычно с вырезкой на наружно-вентральном крае. Средняя голень у самок с 2 апикальными шпорами, у самцов без шпор. (Триба Dinetini) ………………
. ……………………………………………………………Dinetus Panz.
128 (115). Средние голени с 1 апикальной шпорой; если без шпор, то югальная лопасть заднего крыла длиннее анальной области или мандибулы без выемки на наружно-апикальном крае.
129 (130). Жилка М заднего крыла отходит перед жилкой cu-a. Мандибулы с вырезкой или уступом на наружно-вентральном крае. Задние бедра дистально не усеченные. Средние голени с 1 апикальной шпорой. Югальная лопасть заднего крыла длиннее половины длины анальной области. (Триба Laphyragogini) ………………………..
……..……………………………………………….Laphyragogus Kohl
130 (129). Сочетание признаков иное.
131 (134). Жилка М заднего крыла отходит перед жилкой cu-a. Заднее бедро усеченное на дистальном конце или с ложкообразным выступом.
132 (133). Внутренние края глаз сближаются книзу. (Триба Entomosericini) ……………………………………...…..Entomosericus Dahlb.
133 (132). Внутренние края глаз сближаются кверху. (Триба Pseudoscoliini) ……………………………………………....….Pseudoscolia Rad.
134 (131). Жилка М заднего крыла отходит дистальнее жилки cu-a, если иначе, то заднее бедро простое на верщине, без выступа и не усеченное.
135 (140). Внутренние края глаз с глубокой вырезкой или угловато изогнутые.
136 (139). Югальная лопасть заднего крыла значительно короче половины длины анальной области. (Триба Trypoxylonini)
137 (138). Переднее крыло с 1 субмаргинальной ячейкой. Усиковые ямки не соприкасаются с лобно-наличниковым швом. 1-й сегмент брюшка обычно тонкий и длинный, по крайней мере, в 2 раза длиннее своей ширины. ……………………….……Trypoxylon Latr.
138 (137). Переднее крыло с 2 или 3 субмаргинальными ячейками. Усиковые ямки соприкасаются с лобно-наличниковым швом. 1-й сегмент брюшка широкий, короче своей ширины. ……….….Pison Jur.
139 (136). Югальная лопасть заднего крыла большая, ее длина, по крайней мере, равна половине длины анальной области. Последний членик усиков закругленный на вершине и с овальным гладким пятном. 1-й сегмент брюшка обычно шире своей длины. (Триба Philanthini) ……………………………………………..…Philanthus F.
140 (135). Внутренние края глаз без вырезки, цельные.
141 (142). Заднее бедро утолщенное к вершине, на апикальном конце тупое или усеченное. Эпистернальная борозда отсутствует. (Триба
Cercerini) …………………………………………………Cerceris Latr.
142 (141). Заднее бедро не утолщенное к вершине, на апикальном конце не усеченное и не тупое.
143 (164). Переднее крыло с 1 субмаргинальной ячейкой или 1-я субмаргинальная ячейка слита с 1-й дискоидальной ячейкой. Скапус не короче 3 базальных члеников жгутика, взятых вместе.
144 (149). Субмаргинальная ячейка слита с 1-й дискоидальной. (Триба Oxybelini)
145 (146). 1-3-й (иногда 4-й и 5-й) тергумы с боковыми килями ………… ...……………………………………………………Belomictus A.Costa
146 (145). Только 1-й и 2-й тергумы с боковыми килями.
147 (148). Чешуи заднещитика маленькие или отсутсьтвуют. Шип промежуточного сегмента отсутствует. 6-й стернум самки с высоким
килем. ……………………………………………..Belomicroides Kohl
148 (147). Чешуи заднещитика хорошо развитые. Шип промежуточного сегмента развит. 6-й стернум самки без высокого киля. ……………
……………………………………………………..…….Oxybelus Latr.
149 (144). Субмаргинальная ячейка не слита с 1-й дискоидальной ячейкой. (Триба Crabronini)
150 (151). Мандибулы с выемкой на наружно-вентральном крае, на вершине простые. Глаза опушенные. …………...Entomognathus Dahlb.
151 (150). Мандибулы без выемки на наружно-вентральном крае, часто на веоршине с зубцами. Глаза не опушенные.
152 (153). Щутиковая формула: 6+4. Передний вертлуг очень тонкий и длинный. Передний тазик с раздвоенным выступом впереди. …….. ……………………………………………………...Odontocrabro Tsun.
153 (152). Щупиковая формула: 6+4 или 5+3. Передний вертлуг не так сильно удлиненный. Передний тазик без раздвоенного выступа.
154 (155). Щупиковая формула : 5+3. Брюшко стебельчатое, тонкое, длинное. 1-й тергум на конце вздутый. …………....Rhopalum Steph.
155 (154). Щупиковая формула: 6+4. Брюшко висячее, иногда 1-й сегмент более или менее суженный и удлиненный.
156 (157). Глазки расположены в виде равностороннего треугольника. Промежуточный сегмент гладкий или с нежной скульптурой. Вертикаулюс отсутствует. …..…………………Crossocerus Lep. et Serv.
157 (156). Глазки расположены в виде низкого равнобедренного треугольника (редко в виде равностороннего треугольника у некоторых видов Lestica, но тогда голова сильно суженная позади глаз, и у некоторых видов Ectemnius, но тогда имеется вертикаулюс).
158 (161). Вертикаулюс отсутствует. Пигидиальное поле плоское.
159 (160). Югальная лопасть длиннее, чем субмедиальная ячейка заднего крыла. Мандибулы простые на вершине. Самцы с пигидиальным полем, но без щита на голенях.
160 (159). Югальная лопасть короче, чем субмедиальная ячейка заднего крыла. Мандибулы обычно на вершине двухзубцовые. Самцы без пигидиального поля, но со щитом на передних голенях… Crabro F.
161 (158). Вертикаулюс имеется. Пигидиальное поле плоское или чаще в виде желобка.
162 (163). Орбитальные ямки отсутствуют или нечетко выраженные, если ограничены тонким килем изнутри, то лоб вверху без густой умеренной или грубой пунктировки. Усики самцов 12- или 13члениковые. ………………………………...……….Ectemnius Dahlb.
163 (162). Орбитальные ямки четко выраженные. Лоб вверху с очень грубой или умеренно грубой густой пунктировкой. Усики самцов 12-члениковые. …………………………………….…….Lestica Billb.
164 (143). Передние крылья с 2-3 субмаргинальными ячейками, если с одной, то она слита с 1-й дискоидальной ячейкой и скапус короче трех базальных члеников жгутика.
165 (166). Наличник разделен продольными линиями на 3 части. Задние глазки деформированные. (Триба Palarini) ……….……Palarus Latr. 166 (165). Наличник не разделен продольными линиями на 3 части или разделен не полностью. Глазки нормальные.
167 (168). Югальная лопасть заднего крыла значительно длиннее половины длины анальной области. Усиковые ямки расположены ясно выше края наличника. Жилка М заднего крыла отходит проксимальнее жилки cu-a. Воротничок переднеспинки ниже уровня скутума. (Триба Aphilanthopsini) ……..…….………Philanthinus Beaum.
168 (167). Югальная лопасть заднего крыла немного длиннее половины длины анальной области. Усиковые ямки обычно соприкасаются с краем наличника.
169 (178). Жилка М заднего крыла (если имеется) отходит дистальнее жилки cu-a. Щупиковая формула: 6+4. Мандибулярные ямки открытые. (Триба Miscophini)
170 (173). Передние крылья с 3 субмаргинальными ячейками. 2-я из них стебельчатая.
171 (172). Наружно-вентральный край мандибул обычно простой, без выемки. Лоб с V-образным вздутием, иногда имеющим Vобразный киль. Пигидиальное поле отсутствует. Передний вертлуг самцов вогнутый сзади у основания. Передний тазик часто с выступом сзади. ………………………….………Solierella Spin. (часть)
172 (171). Наружно-вентральный край мандибул выемчатый или угловато изогнутый. Лоб без V-образного вздутия. Пигидиальное поле имеется (иногда отсутствует у самцов). Передние вертлуг и тазик самцов не модифицированные.
173 (170). Передние крылья с 1 или 2 субмаргинальными ячейками.
174 (175). Наружно-вентральный край мандибул цельный, без выемки. Передние крылья с 1 субмаргинальной и 1 дискоидальной ячейками. Мезоплеврон с гиперстернаулюсом. Задние крылья без з амкнутых ячеек. Мандибулярные ямки открытые. …………Nitela Latr.
175 (174). Наружно-вентральный край мандибул с выемкой или угловато изогнутый. Передние крылья с 2 субмаргинальными и 2 дискоидальными ячейками. Мезоплеврон без гиперстернаулюса.
176 (177). 2-я субмаргинальная ячейка четырехугольная, не стебельчатая. ……..………………………………………Solierella Spin. (часть)
177 (176). 2-я субмаргинальная ячейка треугольная и стебельчатая...…………………….……………………………….Miscophus Jur.
178 (169). Жилка М заднего крыла отходит от жилки cu-a. Щупиковая формула: 5+3. Мандибулярные ямки закрытые. Передние крылья с 3 субмаргинальными ячейками. (Триба Eremiaspheciini) …………..
…….……………………………………………..Eremiasphecium Kohl
Диагностические признаки: Внутренние края глаз без вырезки, параллельные или слегка сближающиеся кверху или книзу. Глазки круглые. Усиковые ямки соприкасаются или почти соприкасаются с наличником. Усики ♀ 12-члениковые, ♂ - 13-члениковые. Мандибулы без вырезки на наружно-вентральном крае. Пальповая формула: 6+4. Ротовые органы короткие. Мандибулярные ямки открытые (за исключением Ampulex). Гипостома отделена от затылка широкой гуларной областью. Переднеспинка с высоким удлиненным воротничком. Плечевые бугры соприкасаются или почти соприкасаются с тегулами. Нотаули обычно имеются, глубоко вдавленные и тянутся через весь скутум. Косой скутальный гребень отсутствует. Скутеллум с ямчатой поперечной базальной бороздой. Эпистернальная борозда обычно отстутствует. Омаулюс обычно имеется. Средняя голень с 2 апикальными шпорами. Среднетазиковые ямки широко раздвинутые, но средние тазики сближенные, стернальная область между ними узкая, часто с острыми краями; средние тазики без дорсо-латерального киля. Прекоксальные лопасти отсутствуют. Задние бедра простые на вершине. Коготки с единственным зубцом на внутреннем крае или раздвоенные на конце. Передние лапки без гребня щетинок. Аролиумы маленькие. Промежуточный сегмент более или менее удлиненный, с U- образным или почти треугольным дорсальным полем сверху, со стернумом. Переднее крыло с 2-3 субмаргинальными ячейками и 2 возвратными жилками, впадающими соответственно в 1-ю или 2-ю, 2-ю или 3-ю субмаргинальные ячейки. Маргинальная ячейка на вершине заостренная. Югальная лопасть отсутствует или маленькая. Медиальная жилка заднего крыла отходит или перед жилкой cu-a, или от нее, или за ней. Заднее крыло иногда с субкостальной жилкой. Анальная область без анальной жилки. Брюшко сидячее или со стебельком, состоящим из стернума и тергума. 1-й тергум с латеральной линией или килем. Брюшко самца обычно только с 3 видимыми снаружи сегментами. Пигидиальное поле отсутствует. 6-1 стернум самки удлиненный, образует трубку, через которую проходит жало. Вольселла с подвижно соединенными дигитусом и кусписом. Головка эдеагуса с зубцами на вентральном крае. 8-й стернум самца со срединным выступом. Церки имеются (кроме Ampulex).
Осы охотятся на тараканов (в том числе и на синантропных), которых парализуют и затем оттаскивают в подхлдящую полость в субстрате, передвигаясь задом и держа мандибулами жертву за основание усиков. В каждой ячейке запасается 1 таракан, на средний тазик которого откладывается яйцо. Гнездо закрывается кусочками листьев, травинок и прочим подобным материалом.
Подсемейство состоит из 2 триб (Ampulicinae и Dolichurini). В фауне Казахстана отмечены представители только трибы Dolichurini.
Диагностические признаки: Мандибулярные ямки открытые, соприкасающиеся с ротовой ямкой. Наличник широко выпуклый, не кровлеобразный, иногда со срединной лопастью. Метастернум сзади не расщепленный и не Y-образный. Задние тазики почти соприкасающиеся друг с другом. Предпоследний членик лапок сходен по форме с 3-м. Маргинальная ячейка переднего крыла в вершинной части не отходит от края крыла. Брюшко прикреплено позади и выше задних тазиков. Вершина брюшка у самца коническая. Церки имеются.
В трибе 5 родов, из которых в фауне Казахстана представлен только 1 род: Dolichurus.
Типовой вид: Pompilus corniculus Spinola, 1808
Синоним рода: Thyreosphex Ashmead, 1904
Диагностические признаки: Средние и мелкие осы: длина тела от 5 до 13 мм. В основном черные, часто с красными вершинными сегментами брюшка, иногда с белыми пятнами на мандибулах, наличнике, лобном выступе, переднеспинке и тазиках. Внутренний край мандибул самки с 2-3 зубцами, у самца с 1 субапикальным зубцом. Максиллярные пальпы длинные и тонкие, лабиальные короткие. Наличник самца короткий, поперечный, обычно с зубцами на переднем крае. Наличник самки с закругленным или почти прямо усеченным передним краем, часто со срединным продольным килем, с поперечным рядом из 4-8 крепких щетинок. Лабрум скрытый, с закругленной вершиной у самци. Усиковые ямки почти соприкасаются, прикрыты U-образным пластинчатым выступом лба. Лобный киль (или борозда) слабо выражен. Темя выступает над уровнем глаз. Постоцеллярное поле длиннее, чем глазковый треугольник (см. сверху). Затылочный киль U-образный, оканчивается на уровне оснований мандибул. Воротничок переднеспинки шире его длины, короче, чем скутум. Нотаули отчетливые и почти полные. Скутум часто с короткой срединной бороздой впереди. Скутеллум почти плоский. Промежуточный сегмент с широким U-образным морщинистым дорсальным полем, ограниченным сзади поперечным килем. Мезоплевры с омаулюсом, в который впадает передне-вентральный остаток эпистернального шва. Омаулюс часто соединяется с ацетабулярным килем. Стернаулюс более или мене редуцированный. Скробальная борозда имеется, часто образует широкую ямку. Гипоэпимеральное поле сильно выпуклое и часто гладкое. Интеркоксальный киль имеется. Плантулы на лапках имеются, расположены между уплощенными щетинками. Переднее крыло с медиальной жилкой (media), отходящей за cu-a, с 3 субмаргинальными ячейками, из которых 1-я менее чем в 2 раза длиннее 2-й. 1-й интерсубмаргинальный отрезок угловато изогнут и часто с придатком. 3-я субмаргинальная ячейка трапециевидная, иногда почти треугольная. 1-я возвратная жилка впадает во 2-ю субмаргинальную ячейку, 2-я – в 3-ю. Югальная лопасть заднего крыла имеется, но маленькая. Медиальная жилка отходит перед cu-a. Брюшко висячее. 2-й стернум обычно с мощным килем и бороздой или тольтко с бороздой вдоль основания вздутой части.
Биология: Самки используют для гнезд готовые полости в субстрате (земле, древесине, коре деревьев, лесной подстилке) или роют неглубокие (до 8 см) норки в земле. В каждом гнезде устраивают только одну ячейку. Добычей являются тараканы (сем. Blatellidae и Blattidae). В каждое гнездо помещается единственный экземпляр добычи, на который и откладывается яйцо.
Распространение: Род распространен всесветно. Известно 34 вида, в Палеарктике – 4, в Казахстане – 2.
Литература: Гуссаковский, 1952; Казенас, 1976.
Определительная таблица видов рода Dolichurus Latr.
1 (2). Наличник у основания сильно выпуклый, кровлеобразный. Лоб в верхней половине морщинистый и пунктированный. Брюшко с очень редкой неравномерной пунктировкой. Крылья с ясным буроватым затемнением. Последний членик усиков ♀ постепенно сужающийся к вершине, на конце слегка закругленный. Три вершинных сегмента брюшка ♀ буровато-рыжие. ♀ 9 мм. ♂ неизвестен. …………………………………………….…….D. major Kazenas
2 (1). Наличник у основания слегка выпуклый, но не кровлеобразный. Лоб в верхней половине гладкий и блестящий, с редкой мелкой пунктировкой. Брюшко гладкое, без пунктировки. Крылья прозрачные, светлые, с очень слабым буроватым затемнением. Последний членик усиков ♀ почти не сужающийся к вершине, на конце косо усеченный. Лишь последний (иногда и предпоследний) сегмент брюшка ♀ буровато-рыжий. ♀ 5-6 мм, ♂ 3,5-5 мм………
……………………………………………………….D. turanicus Guss.
Описание: Самка. Наличник на переднем крае с широким, прямо усеченным впереди выпуклым выступом (рис. 56а), у основания кровлеобразный, но без четкого срединного киля. Пунктировка наличника нежная и редкая, промежутки между точками гладкие и блестящие. Лобная пластинка (рис. 56б) с неправильной косой морщинистой скульптурой. Лоб выше основания усиков почти до уровня переднего глазка с грубой морщинистой скульптурой и пунктировкой; нижнебоковые части лба с довольно густой, но нежной, постепенно укрупняющейся кверху пунктировкой, без морщин. Темя с редкой пунктировкой. Точки в 3-4,5 раза уже переднего глазка. Промежуток между одним из задних глазков и ближайшим глазом приблизительно в 1,2 раза уже промежутка между задними глазками. Усики длинные и тонкие (рис. 58в). Переднеспинка с сильно выпуклыми и блестящими заднебоковыми углами, посредине и впереди с нежными поперечными морщинами, с неглубокой продольной срединной бороздой. Среднеспинка и щитик гладкие и блестящие, лишь с очень редкими мелкими точками. Заднещитик матовый, с нечеткой микроскульптурой, продольными морщинами и короткими волосками (они приблизительно в 2 раза короче диаметра переднего глазка). Промежуточный сегмент сверху с грубой неправильной ячеисто-морщинистой скульптурой (рис. 56г), на границе с задней поверхностью с поперечной, направленной косо вверх и назад складкой, покрытой короткими нежными волосками. Задняя поверхность промежуточного сегмента неправильно ячеисто-морщинистая, ограниченная по бокам продольной складкой, образующей острый зубец ниже середины. Брюшко гладкое и блестящее, с очень редкими мелкими точками. Тело почти целиком черное. Мандибулы, тегулы, жгутик усиков снизу, голени и лапки буровато-рыжие. 4-6-й сегменты брюшка более или менее буровато-рыжие. Крылья (особенно передние) с равномерным буроватым затемнением. Жилки бурые. Длина тела 9 мм. Самец неизвестен.
Отличия от D. turanicus Guss. приведены в определительной таблице видов рода Dolichurus.
Ареал и распространение в Казахстане: Вид известен (Казенас, 1976) по одному экземпляру, пойманному в северных отрогах Джунгарского Алатау (хр. Малайсары, пер. Архарлы, 140 км сев. Алматы).
Биология и экология: Биология не изучена. Вероятно, самки гнездятся в земле, используя готовые полости, и охотятся на мелких тараканов. Единственная самка этого вида поймана в мелкосопочнике на дне отщелка среди кустов таволги (сухая кустарниково-разнотравная степь) в середине осени (4 октября).
Описание (по Гуссаковскому, 1952): Самка. Наличник посредине переднего края с четырехугольным, на вершине прямо усеченным выступом, как у D. corniculus (Spin.), но основание наличника менее выпуклое и без ясного срединного киля. Лобная пластинка, как у D. corniculus, но скульптура головы другая: лоб лишь в нижней части чуть более чем до половины пунктирован и слабо морщинистый. Верх лба и темя гладкие, лишь с очень немногими разбросанными точками. Строение груди и промежуточного сегмента, как у D. corniculus.
Валик переднеспинки с окаймленными по бокам передними и слегка выдающимися назад задними углами, гладкий и блестящий, лишь с отдельными, очень маленькими и редкими точками. Нотаули среднеспинки и базальная борозда щитика очень глубоко вдавленные. Мезоплевры довольно густо и более грубо, чем среднеспинка, пунктированные. Промежуточный сегмент нежно неправильно морщинистый. Продольные кили на его верхней стороне (особенно боковые) гораздо слабее, чем у D. corniculus. Задняя сторона сегмента матовая, густо морщинистая, сверху ограниченная очень резкой поперечной складкой, а на боковом крае (немного ниже середины) с осьтрым зубчиком. Бока промежуточного сегмента густо, довольно нежно, не особенно правильно, косо морщинистые.
Брюшко сверху гладкое, сильно блестящее, без пунктировки. Тело черное. Мандибулы, кроме вершины, тегулы, жгутик усиков снизу, передние голени (особенно на внутренней стороне) и все лапки буроваторыжие. Вершины средних и задних бедер бурые, последний сегмент брюшка буроватый. Крылья прозрачные, со светло-бурыми жилками. Длина тела 5-6 мм.
Самец. Наличник посредине переднего края 3-хзубцовый (рис. 56з). Лоб густо морщинисто-пунктированный (с неправильными морщинами между точками). Лобный выступ, как на рис. 56е. Темя и виски лишь с редкой (более крупной, чем у самки) пунктировкой, между точками гладкие и блестящие. Грудь с редкой некрупной (но крупнее, чем у самки) пунктировкой, мезоплевры с густой и сравнительно крупной пунктировкой, без морщин в промежутках между точками. Боковой край промежуточного сегмента сзади не с острым зубцом, а лишь с небольшим угловатым выступом ниже середины. Брюшко с более крупной и гораздо более густой пунктировкой, особенно 2-й и 3-й тергумы. Окраска тела почти как у самки, несколько изменчива. Лобная пластинка с белой каймой, иногда переднеспинка с 2 маленькими белыми пятнами у задних углов. Наличник обычно с 2 маленькими белыми пятнами у основания. Длина тела 3,5-5 мм.
Отличия от D. major приведены в определительной таблице. От D. corniculus, по В.В.Гуссаковскому (1952), отличается более мелкими размерами, рыжеватыми мандибулами, передними голенями, лапками и нижней стороной жгутика, менее резкой скульптурой промежуточного сегмента и более грубой пунктировкой брюшка. Не исключено, что D. turanicus является лишь вариацией D. corniculus.
Ареал и распространение в Казахстане: Вид, по литературным данным, известен из Таджикистана (Гиссарский хр., ущ. Кондара), Узбекистана (окр. Самарканда) и Казахстана (Заилийский Алатау, окр. АлмаАты) (Гуссаковский, 1940, 1952; Казенас, 1971, 1972). Один самец найден автором также в Центральном Казахстане (26 км сев.-вост. Целинограда, ныне Астаны), 2 самца, возможно, этого же вида собраны в тугае р. Сыр-Дарьи (3 км южн. Яны-Кургана).
Биология и экология: Самки, вероятно, гнездятся, используя готовые полости в субстрате. Одна самка в окрестностях Алматы поймана с неполовозрелым мелким тараканом рода Ectobius. Обитают в местах с древесной растительностью.
Диагностические признаки: Внутренние края глаз без вырезки, сближающиеся вниз или вверх или параллельные; глазки нормальные. Усики обычно прикреплены к середине лица. Усиковые ямки обычно отделены от лобно-наличникового шва промежутком. Самцы с 13-, самки с 12-члениковыми усиками. Мандибулы без вырезки снизу снаружи. Переднеспинка с высоким воротничком; пронотальная лопасть отделена от тегулы широким промежутком. Нотаули, если имеются, короткие, слабо развитые. Косой скутальный киль не развит. Эпистернальная борозда обычно тянется до передне-вентральной части мезоплевр. Омаулюс отсутствует. Средняя голень обычно с 2 апикальными шпорами. Коготки с 1-5 зубцами или без них. Передние лапки с гребнем щетинок или без него. Аролиум обычно крупный. Промежуточный сегмент с Uобразным срединным полем или без него. Стернум сегмента имеется.
Передние крылья почти всегда с 3 субмаргинальными ячейками и 2 возвратными жилками. Югальная лопасть заднего крыла крупная, с анальной жилкой, занимает большую часть анальной области. Брюшко с цилиндрическитм стебельком, состоящим только из 1-го стернума (у некоторых Ammophilini – из 1-го стернума и 1-го тергума). Пигидиальное поле отсутствует. Церки обычно имеются. 1-й тергум без боковых линий или килей.
Подсемейство включает в себя 3 трибы: Sceliphronini, Sphecini и Ammophilini.
Триба Sceliphronini
Диагностические признаки: Усики самцов с плакоидами (кроме Sceliphron). Усиковые ямки расположены обычно ниже середины лица. Скапус относительно короткий. Наличник обычно шире своей длины (иногда такой же у некоторых видов Chalybion). Верхняя губа шире длины. Мандибулы простые или с 1-2 зубцами на внутреннем крае. Мандибулярная ямка замкнута парамандибулярным выступом. 3-й членик максиллярного щупика обычно расширенный с одной стороны. Гипостомальный киль часто неполный у самки. Срединное поле промежуточного сегмента иногда имеется. Нижнее метаплевральное поле обычно не выражено. Переднее крыло с 3 субмаргинальными ячейками. Обе возвратные жилки обычно впадают во 2-ю субмаргинальную ячейку, редко иначе. Внутренний край коготков с 1 зубцом в середине, редко без зубца. Лапки обычно с плантулами (кроме большинства видов Chalybion). Стебелек брюшка состоит только из 1 стернума. 8-й стернум с церками (кроме большинства видов Sceliphron). Вольселла с подвижно сочлененным дигитусом.
Триба включает 8 родов: Stangeella, Chlorion, Podium, Trigonopsis, Dynatus, Penepodium, Sceliphron и Chalybion. В фауне Казахстана имеется только два: Chalybion и Sceliphron. Не исключено нахождение 3-го рода – Chlorion – в Юго-Западном и Южном Казахстане.
Типовой вид – Chalybion caeruleus (L.), 1763
Диагностические признаки: Внутренние края глаз обычно вверху изугнуты в сторону срединной линии. Голова с многочисленными отстоящими волосками. Лоб выше усиковых ямок в середине выступает в виде плоской или слегка вогнутой площадки, нижние углы которой нависают над ямками. 1-й членик жгутика по длине примерно равен 2-му. Членики жгутика самца с плакоидами. Наличник длинный, обычно длиннее своей ширины, более или менее выпуклый, с 3 или 5 зубцами или лопастями на переднем крае (у подрода Hemichalybion с широкой срединной лопастью, иногда выемчатой в середине). Мандибулы простые, иногда с субапикальным зубцом у самок. Ротовые части короткие. 3-й членик максиллярных щупиков более или менее расширенный с одной стороны. Затылочный киль обычно в виде полного кольца. Воротничок переднеспинки обычно со срединной продольной бороздой. Самка без гребня щетинок. Лапки обычно без плантул. Промежуточный сегмент длинный, без срединного поля, без дыхальцевой борозды по бокам. Скробальная борозда обычно имеется. Маргинальная ячейка обычно острая на вершине. 2-я субмаргинальная ячейка обычно сильно суженная кпереди, редко стебельчатая, в нее обычно впадают обе возвратные жилки. 3-я анальная жилка заднего крыла отделена широким промежутком от края крыла, не короче 1/3 промежутка от основания крыла до югальной вырезки. Эпистернальная борозда длинная. Тело обычно металлически синее.
Биология. Самки устраивают свои гнезда в готовых трубчатых полостях в древесине и стеблях растений, в полостях в стенах и в покинутых глиняных гнездах других ос. Делают перегородки между ячейками и закрывают гнездо влажной глиной (землей), причем не собирают ее во влажных местах, а смешивают глину с водой вблизи гнезда, принося только воду. Интересно, что наружную перегородку осы иногда делают с использованием извести (со стен построек человека) или мочевой кислоты из фекалий крупных позвоночных животных. Добыча – пауки. Оса откладывает яйцо на брюшко одного из пауков.
Род распространен почти по всему миру. Отсутствует только в Южной Америке. Известно более 30 видов. В Казахстане 2 вида.
Литература: Гуссаколвский, 1935; Казенас, 1971, 1972; Bitsch, 1988; Hensen, 1988; Kohl, 1918.
Определительная таблица видов рода Chalybion
1 (2). Задние бедра черные, с металлически синим блеском. Жилки крыльев бурые. 4-1 стернум брюшка без вогнутой площадки. Передний край наличника ♀ с 5, ♂ с 3 зубчиками. Внутренний край мандибул ♀ с субапикальным зубцом. Подрод Chalybion Dahlb. Минимальное расстояние между глазами больше, чем длина 2-го и 3-го члеников усика вместе взятых. Наличник равномерно выпуклый, лишь у ♀ со слабым продольным гребнем. Боковые поверхности промежуточного сегмента морщинистые и пунктированные.
♀ 17-19 мм, ♂ 15-16 мм. Иран, Средняя Азия, Казахстан. …………
………………………………………………Ch. (Ch.) turanicum Guss. 2 (1). ). Задние бедра в дистальной половине красные. Жилки крыльев отчасти рыжие. 4-й стернум брюшка с широкой вогнутой площадкой. Передний край наличника с широкой срединной лопастью, выемчатой в середине у ♂. Внутренний край мандибул ♀ без субапикального зубца. Подрод Hemichalybion Kohl. ♀ 15-20 мм, ♂ 13-
20 мм. ……………………………………..….Ch. (H.) femoratum (F.)
Синоним: Сhalybion chalybeum (Vander Linden), 1827 (Pelopaeus)
Описание: Самка. Мандибулы без зубца перед вершиной. Наличник выпуклый, с продольным килем в середине, с почти прямым или слегка дугообразно изогнутым передним краем, без зубцов (рис. 57а). Внутренние края глаз выемчатые (рис. 57б). Ширина лица внизу (на наличнике) приблизительно в 1, 3 раза меньше максимальной ширины на лбу и почти равна ширине на темени. Промежуток между одним из задних глазков и ближайшим глазом примерно в 1,5 раза шире промежутка между глазками. Базальные членики усиков как на рис. 57в. Переднеспинка почти плоская спереди, круто обрывающаяся впереди, сильно выпуклая, сверху в виде 2 бугров, разделенных глубоким продольным вдавлением (рис. 57г), впереди с нежными поперечными морщинами. Среднеспинка с густой, довольно крупной пунктировкой. Промежутки между точками гладкие и блестящие, с микропунктировкой, с тенденцией к образованию поперечных складок. Щитик с более редкой, чем среднеспинка, пунктировкой, слегка продольно вдавленный в середине. Бока среднегруди густо пунктированные, без складок. Бока заднегруди внизу (позади стигм) с нежными поперечными складками. Бока промежуточного сегмента пунктированные и морщинистые. Верх промежуточного сегмента с нежными, неправильно изогнутыми поперечными складками и густой пунктировкой между складками. Стебелек брюшка приблизительно в 1,7 раза короче 1-го членика задних лапок. 4-й стернум с широким вдавлением. Тело темно-металлически-синее. Мандибулы бурые, отчасти рыжевато-красные. Ноги бурые, задние бедра на большей части красно-рыжие. Тазики впереди бурые, сзади светлорыжие. Крылья рыжевато-желтые, жилки рыжие и коричневые. 15-20 мм.
Самец. Передний край наличника в середине слегка выемчатый (рис. 57д). Ширина лица на наличнике почти равна ширине лица на темени (слегка меньше) и почти в 1,4 раза меньше максимальной ширины на лбу (рис. 57е). Промежуток между задними глазками приблизительно в 1,7 раза уже промежутка между одним из них и ближайшим глазом.
Базальные членики усиков, как на рис. 57ж. Скульптура тела, как у самки. 4-6-й стернумы брюшка слегка уплощенно-вдавленные в середине, 7-й кровлеобразный, с продольным килем, сзади слегка выемчатый в середине. Окраска и опушение тела, как у самки. Задние бедра на большей части красно-рыжие, у основания бурые. Стебелек брюшка примерно в 1,25 раза короче 1-го членика задней лапки, который вдвое длиннее 2-го и втрое длиннее 3-го членика. Крылья слабее затемненные, чем у самки. Длина тела 13-20 мм.
Ареал: Южная и Средняя Европа, Юго-Западная Азия, Казахстан и Средняя Азия.
Распространение в Казахстане: Обнаружен в низкогорьях Западного Тянь-Шаня (р. Бадам, 30-40 км южнее Ленгера). Указывается для Казахстана впервые.
Биология и экология: Обитает в долинах рек, оазисах и низкогорьях. Самки для гнезд используют пустые глиняные гнезда других ос и пчел, возможно, также полости в стеблях растений и древесине. Охотятся на пауков (Aranei).
Описание: Самка. Передний край наличника с 5 широкими закругленными зубцами (рис. 57з), латеральные слабо развитые. Наличник сильно выпуклый, в середине с продольным килем. Ширина лица на наличнике слегка уже, чем на темени (рис. 57и), приблизительно в 1,5 раза уже максимальной ширины на лбу. Промежуток между одним из задних глазков и ближайшим глазом примерно в 1,5 раза шире промежутка между задними глазками. Лоб с четким срединным килем. Базальные членики усика, как на рис. 57к. Воротничок переднеспинки продольно вдавленный в середине задней выпуклой части и поперечно выпуклый в передней половине, с довольно грубыми поперечными складками (рис. 57л), с некрупной пунктировкой. Среднеспинка с густой, довольно крупной пунктировкой. Промежутки между точками имеют слабую тенденцию к образованию поперечных складок (морщин). Щитик равномерно выпуклый, с довольно редкой мелкой пунктировкой. Бока среднегруди с густой пунктировкой, без четких складок. Бока промежуточного сегмента примерно с такой же скульптурой, с едва намеченной тенденцией к образованию косых складок. Дорсальная поверхность промежуточного сегмента кровлеобразно выпуклая, с густой пунктировкой. Промежутки между точками образуют узкие поперечные складки. Стебелек брюшка примерно равен по длине 1-му членику задних лапок.
Голова и грудь с длинными отстоящими светлыми (серыми) волосками. Лицо, переднеспинка и среднеспинка по бокам со слабо развитым прилегающим серым опушением.
Тело фиолетово-синее, крылья с бурым затемнением, особенно в апикальной части и вдоль базальной поперечной жилки переднего крыла. Жилки бурые. Жгутик усиков черно-бурый. Ноги темно-фиолетовосиние, отчасти буроватые. Длина тела 17-19 мм.
Самец. Передний край наличника 3-хзубцовый (рис. 57м). Наличник кровлеобразно-выпуклый, со срединным продольным килем (рис. 57н). Минимальное расстояние между глазами на наличнике больше длины 2-го и 3-го члеников усика вместе взятых примерно в 1,3 раза, немного меньше расстояния на темени (в 1,1-1,2 раза). Промежуток между задними глазками примерно в 1,9 раза уже промежутка между одним из них и ближайшим глазом. Базальные членики усиков, как на рис. 57о. Переднеспинка (рис. 57л) выпуклая в виде 2 бугров, с глубокой продольной срединной выемкой. Среднеспинка с густой и грубой пунктировкой. Бока промежуточного сегмента пунктированные, почти без складок. Тело темное, преимущественно с синим блеском. Стебелек брюшка немного длиннее 1-го членика задних лапок.
Длина тела 15-16 мм.
Ареал: Средняя Азия, северный Иран, Казахстан.
Распространение в Казахстане: Заилийский Алатау и его предгорья, предгорья Джунгарского Алатау, хр. Кетмень, хр. Каратау, Киргизский хребет и его предгорья, хр. Каржантау, хр. Тарбагатай, долины рек Семиречья. Для Казахстана указывают В.В.Гуссаковский (1935 – АлмаАта) и В.Л.Казенас (1971, 1972, 1975).
Биология и экология: Вид встречается в основном в предгорьях и низкогорьях, в оазисах и по долинам рек. Самки для своих гнезд используют готовые полости в древесине, возможно, также пустые глиняные гнезда Sceliphron spp. Несколько гнезд было обнаружено автором в трубчатых полостях в древесине (в чурбаках, напиленных из тонких стволов осины и черешни). Добыча – пауки-тенетники, живущие в кронах деревьев. Во всяком случае, наблюдалось, как самки облетают кроны деревьев, осматривая тенета пауков. П.И.Мариковский (1972) описал охоту синего сцелифрона (вероятно, это Ch. turanicum) на каракуртов (Latrodectus), близ г. Капчагая. Оса влетает в логово паука, парализует уколом жала, затем, отлетев с ним на несколько метров, мнет челюстями ему голову. В каждой ячейке, вероятно, запасается несколько экземпляров пауков.
Типовой вид – Sceliphron spirifex (L.), 1758
Синонимы: Pelopoeus Latreille, 1802-1803; Pelopaeus Latreille, 1804; Sceliphrum Schulz, 1906.
Диагностические признаки: Внутренние края глаз сближаются кверху или параллельные у самки, их взаимное расположение варьирует у самца. Голова с многочисленными отстоящими волосками. 1-й членик жгутика обычно длиннее 2-го. Жгутик самца без плакоид. Усиковые ямки отделены от лобно-наличникового шва ясным промежутком, равным по крайней мере половине диаметра ямки. Наличнике в 1,5-2 раза шире своей длины. Передний край наличника самки с 2 широкими плоскими лопастями, у самца обычно тоже с 2 лопастями. Мандибулы простые, редко с внутренним субапикальным зубцом у некоторых самок; ротовые части короткие. 3-й членик максиллярных пальп расширен на одной стороне. Воротничок переднеспинки короткий. Промежуточный сегмент умеренной длины, с U-образным срединным полем, ограниченным, по крайней мере, на вершине широкой бороздой. Бока сегмента без дыхальцевых борозд. Эпистернальная борозда тянется до передневентрального края мезоплеврона. Маргинальная ячейка закругленная или заостренная на вершине. 2-я субмаргинальная ячейка шире своей длины, в нее впадают обе возвратные жилки. 3-я анальная жилка заднего крыла ясно отделена от края крыла. Самка без гребня щетинок на передних лапках. Коготки обычно с зубцом. Стебелек брюшка обычно длинный. 8-й тергум самца иногда с церками. 9-й стернум самца широко треугольный.
Биология и экология: Самки строят свободные надземные гнезда из влажной глины (грязи) в различных, обычно защищенных от дождя, местах: в углублениях скал, дуплах деревьев и других местах, часто связанных с обитанием человека (под крышами, на стенах домов и других построек, под мостами, в тоннелях и пр.). Гнездо обычно состоит из нескольких ячеек, в которых запасаются пауки. Самка откладывает яйцо на паука, принесенного в ячейку первым (у некоторых американских видов – на последнего паука). Все гнездо обычно покрывается общим слоем глины. В гнездах сцелифронов часто паразитируют осы-блестянки
(Chrysis, Trichrysis, Ceratochrysis и др.), наездники (Acroricnus, Osprynchotus), хальциды (Melittobia), немки (Dolichomutilla), ьмухи-жужжала (Antrax, Hyperalonia) и саркофагиды (Amobia).
Род имеет всесветное распространение. Известно более 30 видов; большинство – обитатели Старого Света.
Род делится на 2 подрода: Sceliphron s. s. и Prosceliphron van der Vecht.
В Казахстане 4 вида. Не исключено обнаружение на юге пятого вида – S. spirifex L.
Литература: Гуссаковский, 1928, 1930, 1934, 1935, 1938; Исламов, 1971; Казенас, 1971, 1972, 1975; Мярцева, 1968; Hensen, 1987; Kohl, 1918; Van der Vecht, 1961; Van der Vecht, Van Breugel, 1968; White, 1962.
Определительная таблица подродов и видов рода Sceliphron Latr.
1 (6). Вершинный стернум ♀ слегка выпуклый, без киля. Внутренние края глаз в нижней половине ясно сближаются книзу. Вершинный тергум ♂ без пигостилей. Грудь более или менее сглаженная, блестящая. ……………………………………….Подрод Sceliphron Latr.
2 (3). Задние тазики нормальные, закругленные на наружном крае. Внутренний край мандибул ♀ с зубцом. Тегулы с желтым пятном, бока среднегруди с желтым пятном ниже тегул. Промежуточный сегмент с желтыми пятнами. ………………..S. (S.) madraspatanum (F.)
3 (2). Задние тазики вздутые на наружной стороне, ясно угловато изогнутые снаружи (если смотреть сверху или снизу). Мандибулы простые. Тегулы желтые (S. destillatorium) или черные (S. spirifex); бока среднегруди без желтого пятна ниже тегул. Промежуточный сегмент без желтых пятен.
4 (5). Тегулы и обычно заднещитик с желтыми пятнами. Стебелек брюшка относительно короткий, не длиннее 5/6 длины задней голени. Задние голени и задние бедра желтые……………………
.………………………………………………S. (S.) destillatorium (Ill.)
5 (4). Тегулы и заднещитик черные. Стебелек брюшка по длине примерно равен задней голени. Апикальная часть задних голеней и бедер черная. Южная Европа, Юго-Западная Азия, Африка. В Казахстане пока не обнаружен. ………………………...…..S. (S.) spirifex (L.)
6 (1). Вершинный стернум брюшка ♀ с довольно острым килем. Внутренние края глаз в нижней половине почти параллельные. Вершинный тергум ♂ с пигостилями. Грудь матовая, с нежной скульптурой. ……….……………….Подрод Prosceliphron v.d.Vecht
7 (8). Тело и ноги с менее обильным желтым рисунком (промежуточный сегмент только на вершине с желтым пятном, стебелек черный, перевязи тергумов узкие). Стебелек брюшка длиннее заднего бедра. Лоб выше основания усиков с резко выраженными бугорками. Вершинные лопасти наличника сильно выступающие. Нижняя часть метаплевр матовая. ………………………..S. (P.) deforme Sm.
8 (7). Тело и ноги с более обильным желтым рисунком (промежуточный сегмент с 5 желтыми пятнами, стебелек брюшка более или менее желтый, перевязи тергумов широкие). Стебелек брюшка короче заднего бедра. Лоб выше основания усиков лишь с неясными бугорками. Вершинные лопасти наличника слабо выступающие. Нижняя часть метаплевр гладкая и блестящая…………………….
………………………...………………………..S. (P.) shestakovi Guss.
Синонимы: flavipes (Christ), 1791 (Sphex); pensite (Illiger), 1807 (Sphex); sardonium (Lepeletier), 1845 (Pelopaeus); sardous (Carrucio), 1872 (Pelopoeus); trinacriense (De Stefani), 1889 (Pelopoeus)
Описание: Самка. Передний край наличника самки с 2 лопастевидными выступами, отделенными с обеих сторон остроугольными вырезками (рис. 58а). Наличник с черно-бурыми длинными отстоящими волосками и темно-серым прилегающим опушением. Минимальная ширина лица на темени приблизительно в 1,4 раза меньше максимальной в середине лба (рис. 58б). Промежуток между задними глазками равен по ширине промежутку между одним из них и ближайшим глазом. Переднеспинка сверху с продольным вдавлением, воротничок спереди не очень крутой, с четкой мелкой, довольно редкой пунктировкой, с очень нежными поперечными морщинами. Среднеспинка с густой нежной поперечно-морщинистой скульптурой, с нечетко ограниченными некрупными редкими точками, сзади с продольными и косыми морщинами. Щитик с уплощенной верхней поверхностью, с продольными нежными морщинами. Бока среднегруди с густой мелкой пунктировкой, только внизу со слабо заметными морщинами. Задние тазики снаружи у основания угловато расширенные. Срединное поле промежуточного сегмента с густой поперечно-косой морщинистой скульптурой и нечетко выраженной пунктировкой, широко продольно вдавленное, у основания с коротким килем, на всей поверхности с одинаковыми нежными, довольно густыми светлыми отстоящими волосками, загнутыми на вершине. Бока промежуточного сегмента с продольными, слегка наклонными нежными довольно густыми морщинами и пунктировкой. Голова и грудь с длинными отстоящими черно-бурыми волосками. Прилегающее опушение слабо развитое. Стебелек брюшка по длине равен 1-му и 2-му членикам задних лапок вместе. Мандибулы на большей части буроватокрасные. Передний край наличника красно-бурый. Голова и грудь черные. Скапус желтый, с маленьким бурым пятном сбоку. Жгутик усиков черный, педицеллюс отчасти бурый. Заднещитик желтый. Тегулы с большим желтым пятном. Стебелек брюшка желтый. Брюшко черное. Вершинная часть передних и средних бедер, задние вертлуги, базальная часть задних бедер, передние и средние голени, задние голени (кроме вершины) желтые. Лапки рыжие. 1-й членик задних лапок на большей части желтый. Длина тела 22-29 мм.
Самец. Срединная лопасть наличника равномерно выпуклая, с почти прямым или слегка выемчатым передним кораем (рис. 58в), с густым светло-серым прилегающим опушением и длинными отстоящими светлыми желтовато- или коричневато-серыми волосками. Лицо относительно широкое (рис. 58г), его максимальная ширина примерно в 1,2 раза больше максимальной. Промежуток между задними глазками слегка уже, чем между одним из них и ближайшим глазом. Верх лба, темя, задняя и нижняя поверхности головы с густыми черными и бурыми длинными волосками. Базальные членики усиков, как на рис. 58д. Переднеспинка с широким продольным вдавлением вверху. Среднеспинка с густыми, нечетко ограниченными точками и поперечными нежными густыми морщинами, с длинными густыми –бурыми волосками. Бока среднегруди с густой некрупной пунктировкой. Щитик слегка продольно вдавленный, с продольными нежными складками и нечетко ограниченными некрупными точками. Срединное поле промежуточного сегмента широко продольно вдавленное в середине, с резкими поперечными складками, впереди в середине с коротким продольным килем, с нежными отстоящими волосками, загнутыми на конце. Стебелек брюшка слегка длиннее 1-го и 2-го члеников задних лапок вместе взятых, короче задней голени.
Окраска почти как у самки. Скапус целиком желтый. Заднещитик черный. Крылья прозрачные, слегка коричневатые, со светлокоричневыми и темно-коричневыми жилками. 1-й членик лапок более или менее желтый, последний членик задней лапки более или менее бурый. Длина тела 15-25 мм.
Ареал: Южная и Средняя Европа, Северная Африка, ЮгоЗападная Азия, Средняя Азия, Казахстан, Китай, Монголия и Южная Сибирь.
Распространение в Казахстане: Один из наиболее обычных и широко рпаспространенных видов в республике. Зарегистрирован в следующих местах: Западный Казахстан – Уральск, Берчогур, Темир, Рынпески, Урда, р. Эмба, Кульсары, Елтай, Индер, п-ов Мангышлак; Центральный Казахстан – 12-30 км юго-западнее Аркалыка, Державинск, 20 км южнее Караула: Юго-Восточный Казахстан – Алматы и окрестности, 10 км южнее Талгара, Тургень, Капчагай, 10 км восточнее Капчагая, 35 км северо-западнее Капчагая, 15 км восточнее Аяк-Калкана, 17 км северо-западнее Баканаса, 20 км южнее Чарына, Панжилов (Жаркент); Южный Казахстан – Кзыл-Орда, Джулек близ Чиили, Казалинск, Арал, 10 км восточнее Успеновки (Жамбылская обл.); Восточный Казахстан – Большенарымское, 25 км южнее Лепсы, пос. Алтай, 35 км южнее Аксуата, 20 км юго-западнее Ленинки, окр. Серебрянска, 15 км восточнее Зайсана, 25 км юго-восточнее Акжара. Для Казахстана вид указывают В.В.Гуссаковский (1935 – Семиречье, Зайсан) и В.Л.Казенас (1971, 1972, 1975, 1977 – Заилийский Алатау, Чарын, Или, Алма-Ата, Талды-Курган, Панфилов, Баканас, Капчагай, Аяк-Калкан, 15-20 км вост. Аяк-Калкана).
Биология и экология. Самки делают лепные гнезда из комочков влажной земли, которые они скатывают с помощью жвал и передних ног на берегах водоемов. Гнездо состоит из нескольких (по нашим наблюдениям, до 21) ячеек, имеющих в среднем длину 3 см и ширину 1,4 см и расположенных параллельно друг другу или под некоторым углом. По данным С.Н.Мярцевой (1968б), гнездо имеет до 85 ячеек. Гнезда сцелифроны помещают в укромных местах: в углублениях скал, крупных камней, в нишах обрывов, в постройках человека (на чердаках, под карнизами крыш, под мостами и в других подобных местах). сверху гнездо покрыто общим толстым глиныным покрытием.
В ячейки помещаются пауки средних размеров, которых осы ловят на деревьях и кустарниках. Яйцо откладывается на 1-й экземпляр добычи. В каждой ячейке запасается до 10 экземпляров добычи. Стадия яйца продолжается 3-4 дня, личинка питается 14-15 дней, затем делает кокон и через 3-4 дня линяет, превращаясь в диапаузирующую личинку (пронимфу).
Взрослые осы питаются нектаром цветков различных растений: молочая, клоповника, тамариска, многих видов зонтичных и др.
Осы встречаются в низкогорьях, в оазисах, долинах рек. Вид обычен в населенных пунктах.
Синонимы: flavofasciatum F.Smith, 1859 (Pelopoeus); affine Maindron, 1878 (Pelopoeus), nec Fabricius, 1793$ lineatipes Cameron, 1900
Вид образует несколько подвидов: rufopictum (F.Sm.), unifasciatum (F.Sm.), atripes (F.Mor.), tibiale Cameron, koreanum Uchida, bicinctum van der Vecht, taiwanum Tsuneki, nipponicum Tsuneki. Казахстанские популяции относятся к подвиду S. deforme atripes (F.Mor.), 1888.
Описание: Самка. Передний край наличника с 2 широкими лопастевидными выступами, разделенными в середине узкой остроугольной выемкой (рис. 58е). Наличник со слабо развитым прилегающим опушением, лоб с более густым желтовато-серым опушением. Максимальная ширина лица в середине лба почти в 1,5 раза больше минимальной на темени (рис. 58ж). Промежуток между задними глазками в 1,7 раза меньше промежутка между одним из них и ближайшим глазом. Базальные членики усиков, как на рис. 58з. Воротничок переднеспинки очень короткий, сверху слабо вдавленный в середине. Среднеспинка с очень густыми нежными поперечными морщинами, с мелкими нечетко ограниченными точками. Щитик слабо выпуклый, уплощенный, без продольного вдавления, сзади с нежными продольными морщинами. Срединное поле с негрубой, неправильной изломанно-морщинистой скульптурой, с широкой, поперечно нежно морщинистой продольной бороздой. Бока среднегруди с довольно густой некрупной пунктировкой и нежной косой морщинистой скульптурой, слабо выраженной внизу. Стебелек брюшка по длине примерно равен 1-му членику задней лапки.
Голова и грудь черные. Передний край наличника буроватый. Вершина мандибул буровато-красная. Наличник с желтым пятном. Скапус спереди обесцвеченный, буроватый, отчасти желтый. Переднеспинка с узкой поперечной полосой. Тегулы бурые, с желтым пятном. Бока среднегруди сверху с желтыми пятнами. Щитик и вершина промежуточного сегмента с желтыми пятнами. Стебелек брюшка черный. Тергумы с более или менее развитыми желтыми перевязями или пятнами у заднего края. Вершинная часть бедер, голени (кроме бурых пятен или полос снизу) и лапки рыжие, отчасти буроватые. Голени более или менее желтоватые снаружи. Жгутик усиков черный, 2-й его членик отчасти рыжеватый, как и вершина 1-го. Последний членик лапок с бурой вершиной. Окраска варьирует. У наиболее темных экземпляров (окр. Семипалатинска) тегулы и промежуточный сегмент с очень маленькими желтыми пятнышками, а брюшко почти целиком черное, только 1-й тергум с желтовато-рыжим поперечным пятном. Длина тела 17-23 мм.
Самец. Передний край наличника в середине с тупоугольной выемкой и по бокам от срединных выступов с дугообразной выемкой (рис. 58и). Лицо относительно широкое (рис. 58к), максимальная ширина в середине лба примерно в 1,4 раза больше минимальной на темени. Промежуток между одним из задних глазков и ближайшим глазом в 2,2 раза шире промежутка между задними глазками. Базальные членики усиков, как на рис. 58л. Стебелек брюшка длинный, изогнутый (рис. 58м), слегка длиннее 1-го и 2-го члеников задних лапок вместе взятых. Скульптура, окраска и опушение тела почти, как у самки. Передний край наличника черный. Тегулы коричневые, с желтым пятном, у основания чернобурые. Промежуточный сегмент у основания по бокам верхней поверхности с желтым пятном. 1-й тергум с широкой перевязью, 2-6-й – с узкой, по бокам выемчатой перевязью. Бедра в вершинной части снаружи коричневые. Лапки отчасти (от основания) светло-коричневые, отчасти (в вершинной части) бурые. Жгутик усиков черный. Длина тела 16-20 мм.
Ареал: Южная и Юго-Восточная Азия, Япония, Дарьний Восток России, Корея, Китай, Монголия, Казахстан, Узбекистан, Таджикистан. Для Казахстана вид указывают Ф.Моравиц (Morawitz, 1888 – Семипалатинск) и В.В.Гуссаковский (Gussakovskij, 1934, 1938 – Зайсан).
Распространение в Казахстане: Восточный Казахстан – Семипалатинск, Зайсан; Юго-Восточный Казахстан – предгорья и низкогорья Заилийского Алатау, Иссык, Рахат, Анатольевка, Алматы, Чемолган; Южный Казахстан – хр. Каржантау (30 км южнее Ленгера).
Биология и экология: Самки делают свободные глиняные гнезда, состоящие из одной ячейки, в защищенных от осадков местах, часто в постройках человека. Мы находили гнезда в жилых помещениях под полкой книжного шкафа (между книгами и полкой), внутри свернутого ковра на балконе, в ящике письменного стола, в щели между картонными коробками под навесом и в других подобных местах. В отличие от S. destillatorium этот вид предпочитает для гнездования узкие полузамкнутые пространства. Ячейки прикреплены к субстрату базальной половиной боковой стенки, располапгаются обычно поодиночке, реже группами до 5 штук. Добыча – мелкие пауки семейств Thomisidae, Philodromidae, Salticidae и др. Взрослые осы питаются нектаром цветков растений. Нами отмечены на цветущем молочае.
Осы встречаются в поймах рек, оазисах, горных ущельях. Обычны в населенных пунктах.
Синонимы: maderospatanum Gmelin, 1790 (Sphex); ? lugubre Christ, 1791 (Sphex), nec Villers, 1789; interruptum Palisot de Beauvois, 1806 (Pelopaeus); bilineatum F.Smith, 1852 (Pelopaeus); separatum F.Smith, 1852 (Pelopaeus); pectorale Dahlbom, 1845 (Pelopoeus); transcaspicum Radoszkowski, 1886 (Pelopoeus).
Вид образует несколько подвидов: tubifex (Latreille), pictum (F.Smith), conspicillatum (A.Costa), kohli Sickmann, andamanicum Kohl, sutteri van der Vecht, formosanum van der Vecht. Казахстанские популяции относятся к подвиду S. madraspatanum tubifex (Latr.).
Описание: Самка. Мандибулы с зубцом на внутреннем крае. Наличник равномерно выпуклый, впереди уплощенный, вытянутый, с 2 широкими лопастевидными выступами, разделенными остроугольной вырезкой, и с 2 маленькими боковыми зубцами (рис. 59а,б). Максимальная ширина лица (в середине лба) приблизительно в 1,4 раза больше минимальной (на темени). Базальные членики усиков, как на рис. 59в. 3-й членик длиннее 4-го. Воротничок переднеспинки относительно узкий, с глубокой продольной выемкой в середине. Среднеспинка матовая, с густыми, довольно нежными поперечными морщинами, с нечетко выраженной пунктировкой и неправильной микроскульптурой. Щитик равномерно выпуклый, с продольным вдавлением в середине, с продольными морщинами. Заднещитик выпуклый, без вдавления, с продольными морщинами. Бока среднегруди только внизу с заметной морщинистой скульптурой, с грубой пунктировкой, более или менее блестящие. Срединное поле промежуточного сегмента поперечно морщинистое, с широким и глубоким продольным вдавлением, с нежным отстоящим коротким опушением. Бока промежуточного сегмента с густыми косыми морщинами. Задние тазики спереди равномерно закругленные. Стебелек брюшка длинный, длиннее, чем 1-й, 2-й и 3-й членики задних лапок.
Тело черное, с желтым рисунком. Мандибулы черные, на вершине буровато-красные. Скапус черный, спереди отчасти желтый. Жгутик усиков черный, к вершине буроватый. Верх переднеспинки, тегулы впереди, пятна на боках среднегруди под тегулами, щитик, заднещитик, боковые полосы на срединном поле промежуточного сегмента, небольшие боковые и большое заднее пятно на промежуточном сегменте, стебелек брюшка, вершины передних и средних бедер, задние вертлуги, базальная часть задних бедер, передние и средние голени, базальная часть задних голеней, 1-й членик всех лапок (кроме вершины) светло-желтые. Лапки на большей части черно-бурые. Крылья с желто-коричневым затемнением. Жилки рыжевато-коричневые. Отстоящие волоски груди желтовато-серые или белые. Длина тела 15-22 мм.
Самец. Передний край срединной лопасти наличника дугообразно выемчатый между 2 угловатыми выступами (рис. 59г). Лицо с довольно густым желтовато-серым прилегающим опушением. Максимальная ширина лица приблизительно в 1,3 раза меньше минимальной ширины на темени или на наличнике (рис. 59д). Базальные членики усиков, как на рис. 59е. Воротничок переднеспинки узкий, с глубокой продольной выемкой в середине. Стебелек брюшка длинный (рис. 59з), явно длиннее, чем 3 базальные членики задних лапок вместе. Скульптура, окраска и опушение тела почти, как у самки. Заднещитик почти без продольных морщин. Жилки крыльев темно- или светло-коричневые. Длина тела 1319 мм.
Ареал: Южная и Юго-Восточная Европа, Северная Африка, Южная, Юго-Восточная и Юго-Западная Азия, Кавказ, Средняя Азия, Южный Казахстан.
Распространение в Казахстане: Южный Казахстан – Казалинск, 15 км южнее Казалинска, Сюткент, Чардара, Муймак, Учсай близ Аральского моря, Яны-Курган, 30 км южнее Ленгера.
Биология и экология: Вид более теплолюбивый, чем S. destillatorium. Встречается в пустынной зоне, но обязательно при наличии водоемов (в оазисах, по долинам рек и около других водоемов, в населенных пунктах, в низкогорьях). Самки делают из влажной глины (грязи) лепные свободные многоячейковые гнезда. Ячейки, по С.Н.Мярцевой (1972а), имеют длину от 19 до 27 мм, ширину – от 5 до 9 мм. Добыча – пауки сем. Argiopidae, Theridiidae, Tetragnathidae и Thomisidae. В одной ячейке запасается до 27 экземпляров пауков (Мярцева, 1972а, Iwata, 1976). Развитие одного поколения в благоприятных условиях происходит за 25-30 дней (Мярцеваб 1968). В гнездах паразитируют осыблестянки родов Stilbum и Chrysis. Взрослые осы питаются нектаром цветков различных растений: Limonium sp., Euphorbia sp., Tamarix spp. и др.
Описание: Самка. Наличник у основания выпуклый, впереди и по бокам уплощенный, в середине переднего края с узкой выемкой, с почти прямыми передне-боковыми краями (рис. 59и,к). Прилегающее опушение наличника слпабее развито, чем на боковых частях лба, выше наличника. Длинные волоски светло-серые. Максимальная ширины лица почти в 1,5 раза шире минимальной (на темени). Промежуток между одним из задних глазков и ближайшим глазом примерно в 1,25 раза шире промежутка между задними глазками. Базальные членики усиков, как на рис. 59л. Воротничок переднеспинки слегка выемчатый сверху (см. спереди) (рис. 59м). Среднеспинка с густыми поперечными, довольнол нежными морщинами и нечеткой редкой пунктировкой. Щитик уплощенный, слабо выпуклый, сзади с нежными продольными морщинами. Срединное поле промежуточного сегмента с широкой продольной бороздой, с густыми поперечными складками (морщинами), с короткими и длинными отстоящими волосками. Бока среднегруди с нечеткой пунктировкой, неправильной шероховатой поперечно-морщинистой нежной скульптурой, с сетчатой микроскульптурой, полуматовые. Бока промежуточного сегмента с редкими, нечетко ограниченными точками, нежной косой морщинистой скульптурой и сетчатой микроскульптурой. Стебелек брюшка равномерно изогнутый (см. сбоку), относительно короткий. Его длина примерно равна длине 1-го членика задних лапок и ¼
2-го.
Тело черное, с желтым рисунком. Мандибулы черные, с красной вершиной. Пятно на наличнике, скапус, верх переднеспинки, пятно на тегулах, пятнышки на боках среднегруди ниже тегул, пятно на щитике, 5 пятен на промежуточном сегменте, стебелек брюшка на большей части, 1-й тергум (кроме основания), перевязи на 2-5-м тергумах, 6-й тергум на большей части, боковые пятна на 5-м и 6-м стернумах, вершинная часть бедер, передние и средние голени, вершинная часть задних голеней, 1-й членик передних и средних лапок (на большей части) желтые. Крылья слегка затемненные, с коричневыми жилками. Длина тела 15-17 мм.
Самец, вероятно, имеет схлдную скульптуру, окраску и опушение тела. Автору не известен.
Ареал: Туркмения, Узбекистан, Южный Казахстан.
Распространение в Казахстане: Вид известен из Западного ТяньШаня (хр. Каржантау, р. Бадам, 30 км южнее Ленгера).
Экология и биология: Встречается в горных ущельях (в низкогорьях), в оазисах, долинах рек, населенных пунктах. Биология неизвестна. Вероятно, самки делают глиняные лепные гнезда и запасают в них пауков.
Диагностические признаки: Усиковые ямки расположены около середины лица и отделены от наличника промежутком, ширина которого обычно меньше ширины ямок. Усики самцов часто с плакоидами. Скапус короткий. 1-й членик жгутика обычно длиннее, чем 2-й (кроме некоторых видов Isodontia и Prionyx). Наличник широкий, шире своей высоты. Внутренний край мандибул с 1-2 зубцами или вершина двух-, трехзубцовая. Мандибулярные ямки замкнуты парамандибулярным выступом гипостомы. Гипостомальный киль полный, затылочный – неполный. Воротничок переднеспинки короткий и высокий. Срединное поле промежуточного сегмента редко ограниченное, дыхальцевая борозда обычно отсутствует. Эпистернальная борозда длинная, доходит или полчти доходит до передне-вентрального края мезоплеврона (за редким исключением). Маргинальная ячейка на вершине закругленная или усеченная. Субмаргинальных ячеек 3. 1-я возвратная жилка обычно впадает во 2-ю субмаргинальную ячейку, 2-я – соответственно в 3-ю. Самка с псаммофором или без него. Средние тазики почти соприкасаются. 2 апикально-вентральные щетинки 5-го членика задних лапок широкие, уплощенные. Коготки с 2-5 зубцами на внутреннем крае. Плантулы отсутствуют. Стебелек брюшка состоит только из 1-го стернума. Церки отсутствуют.
Триба включает в себя 5 родов: Sphex, Isodontia, Palmodes, Chilosphex и Prionyx.
Р.М.Бохарт и А.С.Менке (Bohart and Menke, 1963, 1976) разделили трибу на 2 подтрибы: Sphecina и Prionyxina. В подтрибе Sphecina (Sphex и Isodontia) базальная сторона 2-й субмаргинальной ячейки равна или короче передней стороны, тогда как в подтрибе Prionyxina (Palmodes, Chilosphex и Prionyx) базальная сторона всегда длиннее передней.
В Казахстане обнаружены представители 3 родов трибы: Sphex, Palmodes и Prionyx.
Типовой вид – Sphex flavipennis Fabricius, 1793
Синонимы: Sphaex Scopoli, 1772; Ammobia Billberg, 1882; Proterosphex Fernald, 1905.
Диагностические признаки: Усики самца обычно с плакоидами. Свободный край наличника самки обычно дугообразный, у самца – широко усеченный или выемчатый. Верхняя губа самци обычно со срединным продольным килем, у самца плоская. Мандибулы самки длинные, в середине расширенные, обычно сильно изогнутые, с зубцом на внутреннем крае, у самца короче и тоньше. Воротничок переднеспинки с утолщением, отделен от среднеспинки перетяжкой. Промежуточный сегмент со срединным полем или без него, обычно с дыхальцевой бороздой по бокам. Базальная сторона 2-й субмаргинальной ячейки обычно меньше передней стороны; передняя сторона 3-й субмаргинальной ячейки обычно короче, чем передняя часть задней стороны (до возвратной жилки). Передние ноги самки с хорошо развитым гребнем длинных уплощенных щетинок. 1-й членик передних лапок асимметричный. Стебелек брюшка обычно прямой, короткий. 8-й тергум самца с церками. 8-й стернум самца обычно треугольный.
Биология: Самки гнездятся в земле, обычно на участках с более или менее плотной почвой. Гнезда одно- или многоячейковые, глубиной обычно до 15-20 см. Добычей являются личинки и имаго кузнечиков (Tettigoniidae, Katydidae), реже сверчков (Gryllidae). В одной ячейке от 1 до 6-7 экземпляров. Яйцо откладывается, как правило, на 1-й экземпляр добычи. Весь запас пищи личинка съедает за 10-12 дней, после чего строит кокон. Зимует предкуколка.
Род распространен всесветно. Включает около 120 видов. В Казахстане известно 5 видов.
Литература: Гуссаковский, 1930, 1934, 1935; Казенас, 1969, 1971, 1972, 1975, 1978; Мярцева, 1964; Berland, Bernard, 1949 (1947), Kohl, 1885, 1890, 1895; F.Morawitz, 1891; Olberg, 1966; Roth, 1925; Tsuneki, 1963; Yasumatsu, 1938.
Определительная таблица видов рода Sphex L.
1 (2). Дорсальная поверхность промежуточного сегмента с 4-9 мощными поперечными ребрами. Стебелек брюшка длиннее 2-го членика задних лапок. Основание брюшка ♀ красное, у ♂ черное. Передний край наличника ♀ посредине с вырезкой. ……………………… .…………………………………………………S. (S.) pruinosus Germ.
2 (1). Дорсальная поверхность промежуточного сегмента без мощных поперечных ребер, с более нежной скульптурой. Стебелек брюшка короче 2-го членика задних лапок. (Отстоящие волоски наличника, лба и груди светлые, серебристые или золотистые).
3 (4). Стебелек брюшка не длиннее своей ширины. Переднеспинка и среднеспинка ♀ только со слабым прилегающим опушением. Крылья ♀ с желтым оттенком. Брюшко ♂ черное. Наличник ♂ без вырезки. 8-й стернум ♂ по бокам с бовольно длинными волосками. ………………..…………………………………….S. (S.) afer Lep.
4 (3). Стебелек брюшка в длину больше, чем в ширину.
5 (6). Брюшко целиком черное. ……………………..S.(S.) funerarius Guss. 6 (5). Брюшко более или менее красно-рыжее.
7 (8). Переднеспинка и среднеспинка с хорошо заметным прилегающим опушением. Дорсальная поверхность промежуточного сегмента четко поперечно морщинистая. Крылья ♀ с сильным желтым затемнением. Наличник ♂ без вырезки. …………..S.(S.) flavipennis F.
8 (7). Переднеспинка и среднеспинка почти без прилегающего опушеня. Дорсальная поверхность промежуточного сегмента лишь с едва заметными поперечными морщинками. Крылья ♀ со слабым желтым оттенком. Передний край наличника ♂ посредине с широкой неглубокой вырезкой. …………………………..S.(S.) rufocinctus Br.
Синонимы: tristis Kohl, 1885; ? plumipes Radoszkowski, 1886, nec Drury, 1773; pachysoma Kohl, 1890.
Подвид: sordidus Dahlbom, 1845
Описание: Самка. Наличник без вырезки или с незначительной вырезкой на переднем крае в середине. Наличник и лоб с густым прилегающим серебристым или золотистым опушением и длинными светлыми отстоящими волосками. Внутренние края глаз почти параллельные (рис. 60а). Воротничок переднеспинки в середине не вдавленный. Грудь со слабо развитым, едва заметным опушением и длинными светлыми волосками. Заднещитик не двухбугровый. Промежуточный сегмент сверху без поперечных крупных складок. Стебелек брюшка короткий (рис. 60б), короче 3-го членика задних лапок, не длиннее своей ширины. Средние и задние бедра с длинными бурыми волосками снизу (их максимальная длина почти равна половине ширины бедра)ю Брюшко черное или с красно-рыжим основанием. Крылья желтоватые. Длина тела 21-32 мм.
Самец. Наличник без вырезки или с маленькой вырезкой на переднем крае (рис. 60в), с голой треугольной зоной в середине. Сенсорные поля (плакоиды) на 4-6-м члениках жгутика примерно одинаковые, ограниченные килями (рис. 60г), слегка вдавленные, намного уже половины ширины члеников. Глаза явно сближаются книзу. Промежуток между задними глазками почти в 1,3 раза шире промежутка между одним из них и ближайшим глазом (рис. 60д). Стебелек брюшка длиннее своей ширины (рис. 60е), почти равен 3-му членику задних лапок. Прилегающее опушение тела очень короткое, слабо развитое, но ясно заметное и на брюшке. 8-й стернум брюшка по бокам с густыми светлобурыми волосками. Брюшко матовое. Голова, грудь и брюшко черные. Длинные волоски головы коричневато-серые, на груди - серые. Крылья с почти одинаковым на всей поверхности буроватым затемнением, передние слегка желтоватые. Длина тела 21-24 мм.
Вид имеет в Казахстане 2 различающиеся по самкам формы, которые, вероятно, можно рассматривать как подвиды (однако для окончательного суждения необходим дополнительный материал). Первая характеризуется целиком черным брюшком, более коротким стебельком (рис. 60б) и сильнее выраженной желтой окраской крыльев. Обнаружена в Зайсанской котловине и на севере Балхаш-Алакольской впадины. Вторая имеет рыжевато-красное основание брюшка, слегка более длинный (но не длиннее своей ширины) стебелек и менее выраженную желтую окраску крыльев. Обитает в более южных и западных районах Казахстана.
Ареал: Южная Европа, Северная Африка, Юго-Западная Азия, Средняя Азия, Казахстан.
Распространение в Казахстане: Восточный Казахстан – Кумашкино (Курчум), Зайсан, пески Бозайгыркум, оз. Алакуль, Лепсы; Центральный Казахстан – Тансык, Аягуз, Мынарал; Юго-Восточный Казахстан – предгорья Джунгарского Алатау, Талды-Курган, окр. Капчагая (Или), окр. Алматы, Чарын, о. Барсучий на Капчагайском водохранилище, Баканас, 20 км восточнее пос. Чингильды, Аяк-Калкан, предгорья Киргизского хребта; Южный Казахстан – пески Алко-куль-кум, пуст. Кызылкум, Чардара, Сев. Приаралье, оз. Камышлыбаш, Баламурун (близ Чиили), Арысь, Сюткент, Кантемировка; Юго-Западный Казахстан – п-ов Мангышлак.
Экология и биология: Обычен в лессовых степях и невысоких предгорьях (Гуссаковский, 1935). На равнинах встречается главным образом по долинам рек. Самки роют неглубокие норы в плотном грунте и запасают в них кузнечиков (Tettigoniidae). Взрослые осы ночуют открыто на растениях, иногда образуют скопления (Казенас, Тобиас, 1992).
Синонимы: bicolor Dahlbom, 1845, nec Fabricius, 1775; cinereorufocinctus Dahlbom, 1845; sellae Gribodo, 1873; rufodorsatus De Stefani, 1887.
Описание: Самка. Передний край наличника дугообразный или слегка усеченный в середине. Внутренние края глаз почти параллельные (рис. 61а). Мандибулы, как на рис. 61б. Усики удлиненные. Базальные и вершинные членики, как на рис. 61в,г. Переднеспинка сверху в середине продольно вдавленная, слегка приплюснутая, с густым прилегающим опушением. Среднеспинка с густым прилегающим опушением. Стебелек брюшка длиннее своей ширины (рис. 61д). 1-й членик передних лапок с 11 щетинками снаружи и 4 шипиками изнутри (рис. 61е). Промежуточный сегмент с четкими, почти правильными поперечными морщинами. Волоски и прилегающее опушение наличника золотистые или серебористые. Наличник черный или бурый, в середине впереди более или менее рыжеватый. Переднеспинка черная; густое прилегающее опушение на ней (как и на среднеспинке) латунно-блестящее. Волоски груди светло-серые, слегка желтоватые. Крылья с золотисто-желтым оттенком, в вершинной части бурые. Стебелек брюшка (за редким исключением) и 1-3-й тергумы красно-рыжие. Голени, лапки, передние бедра, отчасти средние и задние бедра красно-рыжие. Шипы ног в основном краснорыжие. Щитик, заднещитик, дорсальная поверхность промежуточного сегмента обычно более или менее красно- или буро-рыжие. Верх мезоплеврона обычно рыжий. Длина тела 26-32 мм.
Самец. Наличник без выемки на переднем крае или с маленькой выемкой. 7-й и 8-й членики усиков сверху с узкими вогнутыми плакоидами (сенсорными полями) (рис. 61ж). Глаза слегка сближаются книзу. Промежуток между задними глазками почти в 1,3 раза шире промежутка между одним из них и ближайшим глазом. Лицо с густым серебристым прилегающим опушением и светлыми желтовато-серыми волосками. Голова и грудь со светлыми волосками. Переднеспинка с продольным вдавлением в середине, с густым прилегающим опушением. Среднеспинка с хорошо развитым прилегающим опушением. Точки довольно густые, но неглубокие. Щитик и заднещитик с прилегающим опушением, микропунктировкой и продольным вдавлением.. Дорсальная поверхность промежуточного сегмента с нежными густыми поперечными морщинами, более четкими по бокам. Стебелек брюшка длиннее 3-го членика задних лапок (рис. 61з). 7-й и 8-й стернумы с довольно густым бурым опушением.
Тело черное. Брюшко в базальной части более или менее краснорыжее. Ноги черные. Стебелек брюшка черный. Крылья желтоватые, но желтый оттенок слабее, чем у самки. Длина тела 20-28 мм.
Вид образует 2 формы. 1-я отличается следующими признаками: переднеспинка в середине слегка продольно вдавленная, волоски и прилегающее опушение лица ♀ золотистое, переднеспинка и среднеспинка ♀ с густым латунно-блестящим прилегающим опушением; шипы ног в основном красно-рыжие, передние бедра красно-рыжие; крылья с яркожелтым оттенком, плакоиды ♂ узкие (не шире 1/3 ширины члеников). 2-я форма характеризуется выпуклой переднеспинкой ♀, отчасти буроватыми длинными волосками наличника ♀, латунно-серебристым прилегающим опушением лица, почти целиком черными передними бедрами, менее отчетливым желтым оттенком крыльев, черными или бурыми шипами ног и более широкими плакоидами ♂ (приблизительно равными половине ширины соответствующих члеников). 1-я форма, которую мы считает типовым подвидом, в Казахстане обнаружена в следующих местах: Алматы, Талгар, предгорья Киргизского хр. и хр. Каржантау. 2-я форма найдена в юго-восточной части Бетпакдалы, Прибалхашье, долине р. Или, в низовьях р. Каскелен, Чу-Илийских горах, пустыне Кызылкум, долине р. Сыр-Дарьи, на п-ове Мангышлак.
Ареал: Южная Европа, Северная Африка, Юго-Западная Азия, Средняя Азия, южная половина Казахстана.
Распространение в Казахстане: Южный Казахстан – о. Барсакельмес, Куланды (Аральское море), пуст. Кызылкум (Чардара); ЮгоВосточный Казахстан – долина р. Или (Аяк-Калкан, Басщий, Капчагай, Баканас, 17 км сев.-зап. Баканаса), Западное Прибалхашье – 5-10 км южнее Мынарала, предгорья и низкогорья Заилийского Алатау (Алматы, Талгар, Чилик, 10 км сев.-зап. Чемолгана, пер. Курдай); предгорья и низкогорья Киргизского хребта (18 км южн. Каменки, 7 км южн. Подгорного); предгорья и низкогорья хр. Каратау (Кентау, пер. Куюк, Беркара), низкогорья хр. Каржантау (30-40 км южнее Ленгера), пуст. Бетпакдала (60 км сев.-зап. Фурмановки); Юго-Западный Казахстан – п-ов Мангышлак. Для Казахстана вид указывают Ф.Ф.Моравиц (Morawitz, 1891 – ст. Киргизская) и В.Л.Казенас (1969, 1972, 1975 – Южный и ЮгоВосточный Казахстан).
Экология и биология: Вид встречается в пустынях и полупустынях, в невысоких горах с аридными ландшафтами, по долинам рек, в оазисах в пустынной зоне. Самки гнездятся в плотной глинистой и щебнисто-глинистой почве. Добыча – крупные кузнечики (Tettigoniidae) и сверчки (Gryllidae). В одной ячейке запасается 3-4 экземпляра добычи (Berland, Berland, 1947). Взрослые осы питаются нектаром цветков различных растений (Limonium sp., Euphorbia sp., Ziziphora bungeana, аккурай, мордовник и др.).
Описание (по В.В.Гуссаковскому): Самка. Форма тела, скульптура головы и груди, как у S. rufocinctus Br. Среднеспинка с мелкой и густой пунктировкой. Промежуточный сегмент матовый, шероховатый, с нежными поперечными морщинами. Стебелек брюшка немного шире, чем у S. rufocinctus, но длиннее, чем у S. afer Lep., длиннее своей ширины. 1-й членик передних лапок с 10 щетинками (редко 11). Тело черное; передние голени на вершине, мандибулы в середине рыжие, края 1-го и 2-го сегментов брюшка буроватые. Ноги отчасти красно-рыжие. Крылья прозрачные, на вершине затемненные. Тело со светлым опушением. Лицо с серебристым опушением и длинными белыми волосками. Длина тела 1722 мм.
Самец. Среднеспинка пунктированная, как у S. rufocinctus; тело более стройное, чем у S. afer. Стебелек брюшка, как у самки. Окраска, как у самки, но ноги целиком черные. Крылья на вершине затемненные, на основании почти прозрачные, слегка желтоватые; края брюшных сегментов буроватые, тело со светлым опушением. Длина тела 16-18 мм.
Ареал: Центральный и Западный Китай, Монголия, Алтай.
Распространение в Казахстане: В.В.Гуссаковский (Gussakovskij, 1934) указывает вид для Зайсанской котловины. Автору этот вид не известен.
Экология и биология: Образ жизни неизвестен. Самки, вероятно, охотятся на кузнечиков и делают гнезда в земле.
Синонимы: vicinus Lepeletier, 1845 (Sphex); scioensis Gribodo, 1879; rothneyi Cameron, 1889; retractus Nurse, 1903.
Описание: Самка. Передний край наличника посредине с вырезкой (рис. 62а,б). Базальные членики усиков, как на рис. 62в. Внутренние края глаз слегка сближаются книзу (рис. 62а). Заднещитик без седловидного вдавления. Промежуточный сегмент сверху очень густо и нежно поперечно морщинистый и с 4-9 крупными поперечными валикообразными складками. Стебелек брюшка (рис. 62г) длиннее 2-го членика задних лапок. Мандибулы и ноги более или менее красно-рыжие. Базальные тергумы брюшка красно-рыжие. Длина тела 20-28 мм.
Самец. Усики относительно длинные. 5-8-й или 5-9-й членики усиков со срединными сенсорными полями (плакоидами), окаймленными 2 резкими параллельными килями. Заднещитик без высоких бугров. Дорсальная поверхность промежуточного сегмента обычно с грубыми поперечными складками. Задняя поверхность промежуточного сегмента с прилегающим серебристым опушением. Стебелек брюшка несколько длиннее 2-го членика задних лапок. 8-й стернум брюшка по сторонам в длинных, но тонких волосках. Брюшко обычно целиком черное. Крылья почти бесцветные, с буроватым вершинным краем. Длина тела 16-22 мм. Ареал: Южная Европа, Северная Африка, Юго-Западная Азия, Средняя Азия, Казахстан, Юго-Восточная Азия.
Распространение в Казахстане: пустыня Кызылкум, Чардара, Джетысай, Тускан, Яны-Курган, Чарын, хр. Каратау, низкогорья хр. Каржантау – 30 км южнее Ленгера; п-ов Мангышлак.
Экология и биология: Пустынный вид. Встречается в невысоких горах, в лессовых предгорьях, в пустынях всех типов, по долинам рек и в культурной полосе. Биология не изучена. Самки, по всей вероятности, гнездятся в земле и охотятся на кузнечиков.
Синонимы: maxillosus Fabricius, 1793, nec Poiret, 1787 (Sphex); ? triangulum Brulle, 1833, nec Villirs, 1793 (Sphex); ? leuconotus Brulle, 1833 (Sphex); obscurus Fischer-Waldheim, 1843, nec Schrank, 1802, nec Fabricius, 1804 (Sphex); pedibusnigris Zanon, 1925.
Название mavromoustakisi Beaumont, 1947 (Sphex) имеет подвидовой статус.
Описание: Самка. Наличник с едва заметной пологой вырезкой на переднем крае (рис. 62д), с длинными светло-серыми, слегка желтоватыми волосками и серебристо-белым прилегающим опушением. Внутренние края глаз параллельные. Виски хорошо развитые. Базальные членики усиков, как на рис. 62е. Переднеспинка слегка продольно вдавленная, без прилегающего опушения. Дорсальная поверхность промежуточного сегмента с нечетко выраженными поперечными складками, с очень нежной сетчатой микроскульптурой, без грубых поперечных складок. Стебелек брюшка короче 2-го членика задних лапок, длиннее своей ширины, длиннее 3-го членика жгутика усиков. Ноги отчасти (передние на голенях и лапках) красно-рыжие. Брюшко отчасти красное. Крылья грязно-серые, слегка желтоватые. Длина тела 18-26 мм.
Самец. Передний край наличника посредине с широкой неглубокой вырезкой. Усики с широкими плакоидами (сенсорными полями) на средних члениках (рис. 62ж). Отстоящие волоски головы и груди светлые. Брюшко с красным основанием. Стебелек длиннее 3-го членика задних лапок, значительно длиннее своей ширины (рис. 62з). 8-й стернум брюшка по бокам с длинными волосками. Длина тела 16-22 мм.
В пустыне Сексеулдала (близ горы Джпмбул) в Юго-Восточном
Казахстане найдено несколько экземпляров самцов, имеющих целиком черное брюшко и широкие плакоиды на 3-6-м члениках жгутика. Несколько сходных самцов обнаружено и в Кыргызстане (в окрестностях Ташкумыра). ПО-видимому, можно выделить их в самостоятельный подвид.
Ареал: Почти вся Европа, Северная Африка, Юго-Западная Азия, Средняя Азия, Казахстан, Западная Сибирь, Западный Китай.
Распространение в Казахстане: Южный Казахстан – о. Возрождения (Николая I), о. Барсакельмес, 5-8 км юго-зап. Чардары, Кзыл-Орда, Баламурун (близ Чиили), Джулек (близ Чиили), Хорхут, Ильчибай, Кара-Узяк, Мешеули (близ Чиили), Акжар – Майлибаш (близ Казалинска), Казалинск, Акыр-Тобе, Ванновка, пер. Куюк (хр. Каратау), 20 км южнее Ленгера (хр. Каржантау), Жамбыл (Джамбул), 35-50 км сев.-зап. Капчагая, 20 км вост. Капчагая, 20 км юго-вост. Чингильды, окр. Талгара, 5 км южн. Талгара, 12 км вост. Капчагая (бывш. Или), 35 км сев. Чилика, Бартогай, Аяк-Калкан, 15 км вост. Аяк-Калкана, Басший, 17 км сев.-зап. Баканаса, 10 км южн. пос. Тургень, р. Кокпатас, пески Сарытаукум, Отар, низовья р. Курты, Балтабай, низовья р. Чарын, низовья р. Каскелен, хр. Турайгыр, Лепсы, Мынарал, хр. Архарлы, хр. Малайсары, Сарканд, Кзыл-Агач; Юго-Западный Казахстан – пос. Онды, 25 км юго-вост. Таучика, хр. Каратау (п-ов Мангышлак); Западный Казахстан – Рын-пески, Индерборский, Январцево, Харькин, Уральск, Темир, Берчогур; Центральный Казахстан – 170 км сев. Балхаша, 15 км юго-зап. Киймы, 20 км сев.-вост. Караганды, Чулак-Эспе, р. Ишим; Северный Казахстан – Наурзум; Восточный Казахстан – Семипалатинск, 23 км сев.-зап. Семеновки, Маканчи, 7 и 40 км юго-вост. с. Алтай, Щербакты, Рыбачье, 45 км южн. Каратогая (оз. Зайсан), 5-10 км сев.-вост. Каратала, 6 км юго-вост. Приозерного, 15-35 км южн. Аксуата, 10 км юго-зап. Акжара, Березовка, 10 км сев.-вост. Малой Владимировки, 5 км южн. пос. Долонь, Семеновка. Для Казахстана вид указывают Ф.Моравиц (Morawitz, 1891 – Рынпески, Букеевская степь) и В.Л.Казенас (1969, 1971, 1972, 1975, 1978).
Биология и экология: Эврибионтный вид, встречающийся в пустынях и полупустынях разных типов, в степях и на открытых участках в лесной и лесостепной зонах; в горах поднимается до лугово-лесного пояса. Самки гнездятся в земле. Гнездо состоит из главного хода длиной до 15 см и яйцевидных горизонтальных ячеек длиной 3-3,5 см, число которых колеблется от 2 до 6. Гнезда располагаются поодиночке или небольшими скоплениями. Добыча – кузнечики родов Phaneroptera, Tettigonia, Platycleis, Decticus и др. В каждой ячейке запасается от 3 до 5 экземпляров. Стадия яйца продолжается 3-4 дня, стадия личинки – до 18 дней, стадия куколки – до 24 дней. Вид поливольтинный. Зимует пронимфа в коконе. В качестве врагов известны мухи рода Metopia. Взрослые осы питаются нектаром цветков различных растений (вероника, зизифора, тимьян, мирикария, крестовник, клоповник, кермек, мята, селитрянка, тысячелистник, синеголовник, тамариск, боялыш, многие зонтичные и др.). Ночуют осы открыто на растениях.
Типовой вид: Chlorion occitanicus Lepeletier et Serville, 1828
Диагностические признаки: Наличник самки более или менее уплощенный, его передний край слегка вогнутый или почти прямой, с выемкой в середине или синусоидовидным изгибом по бокам, у самца синусоидально изогнутый или со срединной выемкой. Верхняя губа у самки с 2 широко расставленными выступами на переднем крае, у самца с дугообразно выступающим или усеченным передним краем, реже с 2 треугольными выступами. Мандибулы у самки с 2 зубцами около середины внутреннего края, у самца с единственным субапикальным внутренним зубцом. Ротовой аппарат короткий. Передние ноги самки с хорошо развитым гребнем длинных, часто уплощенных щетинок. Апикально-вентральные уплощенные щетинки последнего членика средних и задних ног очень широкие, промежуток между ними обычно уже щетинок. Стебелек брюшка по длине равен общей длине 2-4-го члеников задних лапок. Последний стернум самки конический или сильно килеобразный. 8-й стернум самца треугольный или почти прямоугольный. Вальва пениса в вершинной части с зубцами вдоль вентрального края. Апикальная лопасть вольселлы короткая и широкая.
Распространение: Голарктика. В роде 20 видов, распределенных примерно поровну между Палеарктикой и Неарктикой. В Казахстане известно 5 видов.
Биология: Самки устраивают гнезда в земле, ходы для которых роют сами. Гнезда многоячейковые. Добыча – прямокрылые сем. Gryllacrididae и Tettigoniidae, главным образом Decticinae.
Литература: Казенас, 1968, 1969; Kohl, 1890, 1895; Roth, 1963; Tsuneki, 1963.
1 (4). Брюшко ♀ целиком красное. Брюшко ♂ черное; 8-1 стернум его треугольной формы.
2 (3). Волоски головы и груди ♀ беловатые. Среднеспинка ♂ матовая, с нежной микроскопической сетчатой скульптурой, неправильно морщинистая, со светло-коричневым опушением………………... ……..…………………………………………….P. melanarius (Mocs.)
3 (2). Волоски головы и груди ♀ черные. Среднеспинка ♂ блестящая, с нежными поперечными морщинами. Опушение тела черное. ……..
………………………………………………….……P. pusillus (Guss.)
4 (1). Брюшко ♀ черное или отчасти красное. 8-й стернум брюшка ♂ на конце закругленный, с выемкой или прямой.
5 (6). Среднеспинка густо пунктированная и с неправильной микроскопической скульптурой («шагренированная»), у ♂ отчасти морщинистая, матовая. Ноги ♀ на большей части красные. Брюшко ♂
лишь у основания красное или целиком черное. Крылья слегка затемненные. ……………………………………P. strigulosus (Costa)
6 (5). Среднеспинка с другой скульптурой. Ноги ♀ черныею
7 (10). Передний край боковых лопастей наличника ♀ сильно угловато вытянутый, выемка между средней и боковыми лопастями наличника глубокая. Среднеспинка ♂ с двойной пунктировкой на блестящем фоне: с нежной и густой у основания, с крупной и редкой по всей поверхности; кое-где с морщинками. Брюшко ♂ черное.
8 (9). Брюшко ♀ серовато-черное; задние края сегментов слегка красноватые. 8-й стернум ♂ с выемкой на конце. .……P. orientalis (Mocs.)
9 (8). Брюшко ♀ с красным основание. 8-й стернум ♂ прямоугольный, на конце прямой. ……………………………………P. minor (F.Mor.)
10 (7). Передний край боковых лопастей наличника ♀ закругленный, выемка между средней и боковыми лопастями наличника неглубокая. Среднеспинка ♂ с двойной пунктировкой на матовом фоне, морщинистая. …………………………...P. occitanicus (Lep. et Serv.) 11 (12). Брюшко с красным основанием. …………………………………... ………………………………P. occitanicus occitanicus (Lep. et Serv.)
12 (11). Брюшкл сплошь черное. ……………………………………………
…………...…………………………. P. occitanicus puncticollis (Kohl)
Синонимы: anatolicus Kohl, 1888 (Sphex); picicornis F.Morawitz, 1890 (Sphex).
Описание: Самка. Боковые лопасти наличника угловато выступающие, отделены от срединной лопасти глубокой выемкой (рис. 62и). Лицо с густым латунно-серебристо-блестящим опушением, с длинными желтовато-белыми волосками. Глаза слегка сближаются книзу, их внутренний край почти прямой. Ширина лица на темени почти в 1,2 раза больше, чем на наличнике. Промежуток между задними глазками слегка уже, чем промежуток между одним из них и ближайшим глазом. Переднеспинка с густой микропунктировкой и желтовато-белым, серебристоблестящим густым прилегающим опушением, с отдельными мелкими точками. Среднеспинка с неравномерной, в целом густой мелкой пунктировкой и нежными косыми (впереди по бокам), нечетко выраженными морщинами, с отдельными более крупными точками, с густым (особенно по бокам) светлым опушением. Щитик сильно выпуклый, с микропунктировкой, отдельными более крупными точками, без нежных морщин, в середине продольно вдавленный, по краям и в середине с густым прилегающим светлым опушением. Дорсальное поле промежуточного сегмента с четкими поперечными (по бокам скошенными) острыми складками, в середине (особенно впереди и сзади) уплощенновдавленное. Средняя часть его полуматовая, с нежным отстоящим опушением; боковые части (особенно сзади) голые и блестящие. Стебелек брюшка примерно в 1,2 раза короче 1-го членика задних лапок. Брюшко красно-рыжее, иногда отчасти буроватое. Стебелек черный. Ноги черные, с прилегающим светлым опушением. Коготки красно-рыжие. Лапки более или менее буроватые. Крылья слегка коричневатые, в вершинной части с буроватым затемнением. Длина тела 20-27 мм.
Самец. Передний край наличника в середине слегка выемчатый (рис. 62к). Глаза сильно сближаются книзу. Ширина лица на уровне задних глазков примерно в 1,6 раза больше его максимальной ширины. Наличник и лицо с густым серебристым прилегающим опушением и отдельными светлыми, желтовато-серыми длинными волосками. Промежуток между задними глазками примерно в 1,1 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка с густой, очень мелкой пунктировкой и отдельными, нечетко ограниченными более крупными точками, с прилегающим коричневато-серым коротким опушением (обычно более или менее стертым). Среднеспинка полуматовая, с неправильной густой поперечно-морщинистой, довольно нежной скульптурой и нечетко ограниченными некрупными точками, с полуотстоящим коротким коричневато-серым опушением (довольно слабо развитым) и отдельными длинными светлол-серыми волосками. Щитик с нежными продольными морщинами на всей поверхности и редкой пунктировкой. Срединное поле промежуточного сегмента с довольно грубыми острыми поперечно-косыми морщинами, с очень слабо развитым коротким отстоящим опушением. Стебелек брюшка почти равен по длине 1-му членику задних лапок. 8-й стернум брюшка треугольной формы (рис. 62л). Брюшко целиком черное. Ноги черные. Коготки рыжевато-красные. Крылья буроватые, в вершинной части темнее. Длина тела 14-20 мм.
Ареал: Южная Европа, Северная Африка, Юго-Западная Азия, Средняя Азия, Казахстан.
Распространение в Казахстане: Южный Казахстан – окр. ст. Чокусу, 6 км зап. пос. Аманоткель, 29 км сев.-зап. Аральска, 22 км зап. Фурмановки, 8 км зап. Чардары, 18 км сев.-зап. Кантемировки; ЮгоВосточный Казахстан – Или, Баканас, 17 км сев.-зап. Баканаса, Сугур (близ Чилика), 10 км сев.-зап. Чемолгана; Восточный Казахстан – 5 км сев.-вост. Улькен-Каратала. Для Казахстана вид указывается
В.Л.Казенасом (1969, 1972, 1975).
Биология и экология: Пустынный вид. Встречается в песчаных, глинистых и каменистых пустынях, на сухих лугах в долинах рек пустынной зоны. Возможно, зимуют имаго (есть ранневесенние и позднеосенние находки). Образ жизни неизвестен, однако можно предположить, что самки устраивают гнезда в земле и охотятся на кузнечиков. Имаго питаются нектаром цветков различных растений (молочай, ластовень, кермек, кузиния, ак-курай, зизифора, тамариск и др.).
Описание: Самка. Срединная лопасть наличника узкая, с почти прямым или слегка выемчатым передним краем; боковые лопасти угловато выступающие, отделенные от срединной лопасти глубокой и широкой выемкой (рис. 63а). Внутренние края глаз слегка сближаются книзу. Ширина лица на темени примерно в 1,15 раза больше, чем на наличнике. Промежуток между задними глазками примерно равен по ширине промежутку между одним из них и ближайшим глазом. Лицо с густым серебристо-серым прилегающим опушением (обычно отчасти стертым) и длинными бурыми волосками, только наличник впереди с широкой голой каймой. Переднеспинка без срединной выемки, равномерно выпуклая, с редкой мелкой пунктировкой и редкой микропунктировкой, блестящая, с нечеткой, очень нежной микроскульптурой. Среднеспинка с отдельными мелкими точками и неравномерной микропунктировкой, с нечеткой сетчатой микроскульптурой или почти гладкая, блестящая, в середине почти плоская (или слегка вдавленная). Щитик со слабым продольным вдавлением, с отдельными мелкими точками, гладкий и блестящий. Заднещитик бугровидно выпуклый, с нечеткой микропунктировкой и поперечными слабо выраженными морщинами. Дорсальная поверхность промежуточного сегмента с поперечными густыми морщинами, с неправильной шероховатой микроскульптурой, полуматовая, с нежными короткими отстоящими волосками, лишь сзади по бокам голая и более блестящая. Стебелек брюшка примерно в 5 раз длиннее своей ширины, почти в 1,3 раза короче 1-го членика задних лапок. Ширина 2-й субмаргинальной (радиальной) ячейки на радиальной жилке почти в 1,4 раза больше ширины 3-й. Голова и грудь черные. Тегулы сзади рыжевато-бурые. Крылья у основания прозрачные, светлые, в середине слабо, в вершинной части сильнее затемненные. Брюшко у основания (от стебелька) рыжевато-красное (1-3-й тергумы, бока 4-го, 3-5-й стернумы). Нолги черные. Лапки рыжевато-буроватые. Коготки красно-рыжие.
Длина тела 14-20 мм.
Самец. Передний край срединной лопасти наличника почти прямой (рис. 63б) или с выемкой. Наличник с узкой гладкой каймой у переднего края, с густой микропунктировкой на остальной поверхности и серебристым прилегающим опушением, с отдельными длинными чернобурыми волосками. Глаза сильно сближаются книзу. Лицо на темени примерно в 1,6 раза шире, чем на наличнике. Промежуток между задними глазками по ширине примерно равен промежутку между одним из них и ближайшим глазом. Переднеспинка с отдельными мелкими точками, с неравномерной микропунктировкой, с едва заметной сетчатой микроскульптурой, блестящая. Среднеспинка с неравномерной пунктировкой (точки разной величины), в середине и по краям с густой и мелкой пунктировкой и неправильной микроскульптурой, полуматовая, на остальной поверхности блестящая, с гладкими промежутками между точками. Щитик с редкой пунктировкой, с сетчатой микроскульптурой. Заднещитик полуматовый, с нежными поперечными морщинами. Дорсальная поверхность промежуточного сегмента с острыми поперечными густыми морщинами, почти без микроскульптуры, блестящая, с нежными отстоящими волосками, у боковых краев голая. Стебелек брюшка по длине примерно равен 1-му членику задних лапок. 8-й стернум с почти параллельными или дугообразно изогнутыми боковыми краями, на заднем крае без выемки или с маленькой выемкой (рис. 63в). Тело, ноги и усики черные. Передние крылья со слабым бурым затемнением, задние светлее, в вершинной части обе пары с более сильным затемнением. Жилки бурые. Лапки буроватые. Коготки красно-рыжие. Длина тела 1218 мм.
Ареал: Юго-Западная Азия, Средняя Азия, Южный Казахстан.
Распространение в Казахстане: Чу-Илийские горы, Кендыктас, восточные отроги Заилийского Алатау – хр. Турайгыр близ р. Чарын, хр.
Каратау – пер. Куюк, п-ов Мангышлак.
Биология и экология: пустынно-полупустынный вид. Преимущественно встречается на щебнисто-глинистых склонах невысоких гор в пустынной зоне. Образ : жизни не изучен. Самки, вероятно, делают одно- или многоячейковые гнезда в плотном грунте и охотятся на кузнечиков.
Синонимы: proditor Lepeletier, 1845 (Sphex); confinis Dahlbom, 1845 (Sphex); montanus F.Morawitz, 1889 (Sphex); ? solieri Lepeletier, 1845 (Sphex); mandarina F.Smith, 1856 (Sphex).
Подвиды: perplexus (F.Smith), 1856 – Китай; ? australis (Saussure), 1867 - ?; syriacus (Mocsary), 1881 – Восточное Средиземноморье, ЮгоЗападная Азия; puncticollis (Kohl), 1888 – Cредняя Азия; cyrenaicus (Gribodo), 1924 – Ливия; barbarus (Roth), 1963 – Марокко, Алжир; gaetulus (Roth), 1963 – Алжир, Тунис; ibericus (Roth), 1963 – Иберийский п-ов.
Описание типового подвида. Самка. Срединная лопасть наличника с прямым или слегка выемчатым передним краем; боковые угла (боковые лопасти) закругленные, отделены неглубокой выемкой от срединной лопасти (рис. 63г). Наличник с густым серебристым прилегающим опушением, с длинными черными волосками, впереди с голой каймой. Глаза почти параллельные, слегка сближаются книзу. Промежуток между задними глазками приблизительно в 1,1 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка с мелкой редкой пунктировкой, с густой микропунктировкой и короткими светлыми (желтовато-серыми) прилегающими волосками. Среднеспинка с редкими мелкими точками; промежутки с довольно густой микропунктировкой, нечеткой сетчатой микроскульптурой, слабо блестящая, с короткими светлыми полуприлегающими и длинными черно-бурыми волосками. Щитик с редкой мелкой пунктировкой, сетчатой микроскульптурой, с нежными продольными складками сзади, сильно выпуклый, со слабым продольным вдавлением в середине. Заднещитик тупоугольно выпуклый, с поперечными нежными морщинами, полуматовый. Дорсальная поверхность промежуточного сегмента с однородными густыми поперечными морщинами, с нечеткой микроскульптурой, нежными коричневато-серыми волосками. Стебелек брюшка короткий, примерно в 3 раза длиннее своей ширины в середине, значительно короче 1-го членика задних лапок, но длиннее 2-го. Тело черное; брюшко только с красным основанием (1-й и 2-й тергумы на большей части, частично 1-й и 2-й стернумы). Лапки отчасти буроватые. Коготки красно-рыжие. Крылья слегка буроватые. Длина тела 18-25 мм.
Самец. Передний край срединной лопасти наличника слегка дугообразно выемчатый или почти прямой (рис. 63д). Наличник с густым серебристо-серым опушением и длинными густыми черными волосками. Глаза сильно сближаются книзу. Ширина лица на темени почти в 1,3 раза больше ширины на нпаличнике. Ширина промежутка между задними глазками почти равна (или слегка больше) ширины промежутка между одним из них и ближайшим глазом. Переднеспинка с редкими некрупными точками, густой микропунктировкой, очень короткими желтовато-серыми полуприлегающими волосками и длинными черными волосками, с неправильной сетчатой микроскульптурой. Среднеспинка с довольно густой крупной пунктировкой, неравномерной (в целом густой) микропунктировкой, неправильной сетчатой микроскульптурой. Промежутки между точками местами на боках образуют неправильные нечеткие складки (морщины). Опушение слабо развитое, не прилегающее, желтовато-серое. Длинные волоски черные. Щитик со слабым срединным продольным вдавлением, с крупной, довольно густой пунктировкой, с сетчатой микроскульптурой, полуматовый, с нечеткой редкой микропунктировкой, сзади с заметными продольными морщинами. Заднещитик с неправильной поперечно-морщинистой скульптурой и неправильной микроскульптурой. Дорсальная поверхность промежуточного сегмента с довольно грубыми острыми поперечными морщинами, с неправильной микроскульптурой, микропунктировкой, нежными отстоящими волосками на всей поверхности, у основания и у вершины уплощенно-вдавленная. Стебелек брюшка явно короче 1-го членика задних лапок. Ширина 2-й субмаргинальной ячейки на радиальной жилке в 1,61,9 раза больше ширины 3-й. Голова, грудь и ноги черные. Коготки красно-рыжие. Крылья буроватые, у вершины темнее. Основание брюшка (вершина 1-го стернума, 2-й стернум на большей части, 1-й тергум на большей части, 2-й тергум по бокам) красно-рыжие. 8-й стернум на вершине с закругленными углами, обычно в середине с вырезкой, к вершине слегка суженный или с почти параллельными сторонами (рис.
63е). Длина тела 16-22 мм.
Подвид P. o. puncticollis Kohl отличается от номинативного подвида целиком черным брюшком, несколько более крупными размерами (♀ 20-28 мм, ♂ 20-25 мм) и сильнее затемненными крыльями.
Ареал: Южная Европа, Северная Африка, Юго-Западная Азия,
Средняя Азия, Казахстан, Центральная Азия, Сибирь, Восточная и ЮгоВосточная Азия.
Распространение в Казахстане: Подвид P. o. puncticollis – п-ов Мангышлак (25 км юго-вост. Таучика), пески Кызылкум, хр. Каржантау (40 км юго-вост. Ленгера), долина р. Сырдарьи (Джусалы, Чаган), хр. Каратау (пер. Куюк), долина р. Или (17 км сев.-зап. Баканаса), р. Чарын. Подвид P. o. occitanicus – Заилийский Алатау (ущ. Левый Талгар, Тургеньское ущ.), хр. Кетмень (окр. Подгорного), Джунгарский Алатау (хр. Малайсары), Зайсанская котловина (40 км сев.-вост. Кокпекты, Акжар), Прииртышье (Канонерка, Большая и Малая Владимировки), южн. предгорья Ивановского хр. (Березовка), Казахский мелкосопочник (БаянАул, 45 км сев.-вост. Караганды), Калбинский хр., Бухтарминское водохранилище (с. Алтай), Сев. Приаралье (Шокысу). Для Казахстана вид указывают П.Рот (Rjth, 1963 – Сырдарья) и В.Л.Казенас (1969, 1971, 1972, 1975).
Биология и экология: Номинативный подвид приурочен в основном к невысоким горам с пустынно-степной и степной растительностью, P. o. puncticollis – к песчаным, каменистым и глинистым пустыням и поймам рек в пустынной зоне. Самки гнездятся в земле, часто в плотном щебнисто-глинистом грунте. Добыча – кузнечики (Tettigoniidae). В гнезде 1 ячейка, в которой запасается 1 экземпляр добычи. Взрослые осы питаются нектаром цветков различных растений.
Описание: Срединная лопасть наличника с почти прямым передним краем. Боковые лопасти угловато выступающие, отделенные от срединной лопасти довольно глубокой вырезкой (рис. 63ж). Лицо с густым серебристым опушением и длинными бурыми волосками. Внутренние края глаз почти параллельные, слегка сближающиеся на уровне переднего глазка. Промежуток между задними глазками в 1,3-1,4 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка с густой микропунктировкой, густым серым прилегающим опушением, с отдельными мелкими точками. Среднеспинка с отдельными мелкими точками, с неравномерной (в целом густой) микропунктировкой, густым полуприлегающим светло-серым опушением, с отдельными более длинными отстоящими коричневыми волосками, без сетчатой микроскульптуры. Щитик без продольного вдавления, с мелкими редкими точками, с довольно густой микропунктировкой, со слабо заметной сетчатой микроскульптурой. Заднещитик полуматовый, с густым отстоящим опушением, с нечеткими поперечными нежными морщинами. Дорсальная поверхность промежуточного сегмента с нежными поперечными густыми складками (морщинами), с тонкими отстоящими довольно густыми коричневато-серыми волосками , сзади по бокам более или менее сглаженная, полуматовая, голая, с очень нежной поперечно-морщинистой микроскульптурой. Стебелек брюшка примерно в 6 раз длиннее своей минимальной ширины и в 1,3 раза короче 1-го членика задних лапок. Ширина 2-й субмаргинальной ячейки на радиальной жилке в 1,3-1,4 раза больше ширины 3-й. Голова, грудь, брюшко, ноги, усики черные. Крылья слегка буроватые, в вершинной части сильнее. Жилки рыжеватокоричневые, субкоста бурая. Длина тела 25-30 мм.
Самец. Боковые лопасти наличника тупоугольно выступающие, не закругленные (рис. 63з). Передний край срединной лопасти наличника слегка выемчатый или почти прямой. Лицо со слабо развитым светлосерым прилегающим опушением и длинными бурыми волосками. Внутренние края глаз сильно сближаются книзу. Ширина лица на темени в 1,5 раза больше ширины на наличнике. Промежуток между задними глазками в 1,1 раза больше промежутка между одним из них и ближайших глазом. Переднеспинка с довольно крупными редкими точками, с бурыми длинными волосками. Среднеспинка с однородной очень густой, но не очень крупной ячейкообразной пунктировкой (промежутки между точками образуют неправильные негрубые морщины), с микропунктировкой и микроскульптурой, шероховатая, с бурыми длинными волосками и очень нежным коротким полуотстоящим, слабо развитым светлым опушением. Щитик с некрупной изреженной пунктировкой, неправильной микроскульптурой (полуматовый), с нежными продольными морщинами, особенно в задней части. Заднещитик бугровидно выпуклый, с поперечными и продольными морщинами. Дорсальная поверхность промежуточного сегмента с густыми острыми поперечными морщинами, с нечетко ограниченными редкими точками, с микропунктировкой, нежными короткими отстоящими волосками на всей поверхности. Стебелек брюшка примерно в 1,25 раза короче 1-го членика задних лапок. 8-й стернум с вырезкой (рис. 63и). Тело черное. Лапки отчасти буроватые. Коготки красно-рыжие. Крылья буроватые, в вершинной части сильнее. Длина тела 16-25 мм.
Ареал: Юг Восточной Европы, Кавказ, Туркмения, Казахстан.
Распространение в Казахстане: П-ов Мангышлак – 25 км юго-вост. Таучика; предгорья Киргизского хр. – 8 км южнее Каменки, пуст. Сексеул-Дала – 60-70 км сев. Фурмановки; Тургайская ложбина – Наурзум; Прииртышье – Семипалатинск, 23 км сев.-зап. Семеновки, 9 км юговост. с. Долонь, 6 км юго-вост. пос. Щербакты, 8 км юго-вост. Большой
Владимировки; Казахский мелкосопочник – 70 км юго-юго-зап. БаянАула. Для Казахстана вид указывает В.Л.Казенас (1969).
Экология и биология: Пустынно-степной вид. Предпочитает участки с песчаной и глинисто-песчаной почвой. Образ жизни не известен. Вероятно, самки охотятся на кузнечиков и делают гнезда в земле.
Взрослые осы питаются нектаром цветков различных растений (мяты, вероники, цельнолистника, кермека, синеголовника, гипсофилы, зизифоры и др.).
Синонимы: ferus Dahlbom, 1843 (Sphex), nec Drury, 1782; straboni Berland, 1927 (Sphex).
Описание: Боковые лопасти наличника угловато выступающие, отделены от срединной лопасти глубокой выемкой (рис. 63к). Лицо с густым латунно-серебристым прилегающим опушением, со светлыми желтоватыми и коричневатыми длинными волосками. Глаза заметно сближаются книзу. Ширина лица на темени почти в 1,2 раза больше, чем на наличнике. Ширина промежутка между задними глазками почти в 1,1 раза меньше промежутка между одним из них и ближайшим глазом. Переднеспинка с густой микропунктировкой и коротким прилегающим светлым опушением, с отдельными слабо заметными мелкими точками. Среднеспинка с густой мелкой пунктировкой (с тенденцией к образованию нежных неправильных косых молрщин). Опушение довольно слабо развитое. Длинные волоски груди светлые, желтовато-серые. Щитик сильно продольно вдавленный в середине, с мелкой, на выпуклых участках довольно редкой пунктировкой. Заднещитик без поперечных морщин. Дорсальная поверхность промежуточного сегмента с густыми поперечными морщинами, с отстоящими нежными довольно короткими волосками на всей поверхности. 3-я субмаргинальная ячейка на радиальной жилке по ширине почти равна 2-й. Стебелек брюшка примерно в 1,2 раза короче 1-го членика задних лапок. Голова и грудь черные. Тегулы сзади рыжевато-бурые. Крылья буроватые, в вершинной части темнее. Стебелек брюшка черный. 1-й и 2-й сегменты брюшка (позади стебелька) красно-рыжие; 2-й тергум отчасти буроватый. Вершины передних и средних бедер, передние и средние голени и лапки красно-рыжие. Задние голени и лапки бурые, отчасти рыжеватые. Шипы и щетинки ног светлые (желтовато-белые). Передние шпоры рыжие, средние и задние бурые. Длина тела 17-22 мм.
Самец. Срединная лопасть наличника узкая, в середине с выемкой, отделена от боковых угловатой выемкой (рис. 63л). Лицо с не очень густым серебристо-серым прилегающим опушением, с длинными бурыми волосками. Глаза сильно сближаются книзу. Лицо на темени более чем в 1,4 раза шире, чем на наличнике. Промежуток между задними глазками почти равен по ширине промежутку между одним из них и ближайшим глазом. Переднеспинка с густой микропунктировкой, отдельными более крупными, нечетко ограниченными точками и с прилегающим светлым (коричневатым) опушением. Среднеспинка полуматовая, с густой мелкой пунктировкой и микропунктировкой, с тонкими короткими полуотстоящими коричневато-серыми волосками (особенно по бокам), с немногими буроватыми длинными волосками; промежутки между точками образуют неправильную морщинистую скульптуру. Щитик с сетчатой микроскульптурой, с довольно густыми мелкими точками, почти без продольных морщин. Заднещитик без поперечных морщин. Дорсальная поверхность промежуточного сегмента с густыми поперечными морщинами и довольно грубой микроскульптурой, полуматовая, с короткими нежными отстоящими, изогнутыми на конце волосками на всей поверхности. Стебелек брюшка немного короче 1-го членика задних лапок. 8-й стернум на конце с вырезкой (рис. 63м). Все тело черное. 1-й тергум красновато-буроватый у основания. Коготки красно-рыжие. Членики лапок на конце буровато-красноватые. Длина тела 14-20 мм.
Ареал: Южная Европа, Юго-Западная Азия, Средняя Азия, Казахстан.
Распространение в Казахстане: Долина р. Сырдарьи, хр. Каратау – 30 км сев.-зап. Ванновки, Ачисай, ущ. Костобе; Чу-Илийские горы (Айтау) – Отар; предгорья и низкогорья Джунгарского Алатау – Кзыл-Агач, хр. Малай-Сары, хр. Катутау – Коныролен; окр. Алматы; долина р. Лепсы.
Биология и экология: Вид встречается в пустынной зоне – в невысоких горах с глинисто-щебнистыми склонами, в лессовых предгорьях, по долинам рек. Самки гнездятся в земле и охотятся на крупных личинок кузнечиков (Tettigoniidae). Взрослые осы питаются нектаром цветков различных растений (ферула, прангос, боялыш, цельнолистник, молочай и др.).
Типовой вид: Ammophila kirbii Van der Linden, 1827
Синонимы: Prionyx Dahlbom, 1847; Enodia Dahlbom, 1843; Yarpactopus F.Smith, 1856; Parasphex F.Smith, 1856; Gastrosphaeria A.Costa, 1858; Pseudosphex Taschenberg, 1869; Calosphex Kohl, 1890; Neosphex Reed, 1894.
Диагностические признаки: 1-й членик жгутика самца равен или короче 2-го; членики жгутика равны или некоторые из них (обычно 3-5й) с плоскими плакоидами. Наличник самки обычно выпуклый, с прямым или дугообразным передним краем, со срединной вырезкой. Наличник самца с дугообразным или трапециевидным, иногда слабо выемчатым передним краем. Мандибулы самки обычно с 2 зубцами около середины внутреннего края, мандибулы самца с одним субапикальным внутренним зубцом. Ротовые органы иногда удлиненные. Эпистернальная борозда обычно доходит до передне-вентрального края мезоплеврона. Метаплеврон часто желобообразно вогнутый. Субмаргинальных ячеек обычно 3. 1-я и 2-я возвратные жилки впадают соответственно во 2-ю и 3-ю субмаргинальные ячейки. Передние ноги самки с хорошо развитым гребнем длинных и расширенных щетинок. Промежуток между 2 вершинными щетинками средних и задних лапок обычно больше ширины щетинок. Коготки с 2-5 базальными зубцами на внутреннем крае.. Стебелек брюшка сильно варьирует по длине. 4-5-й стернумы самца, в отличие от соседних, обычно с микроопушением. 8-й стернум самца треугольный и часто килеобразный. Вершина вальвы пениса с зубцами вдоль вентрального края; апикальная лопасть вольселлы длинная и тонкая.
Распространение: Род распространен всесветно. Известно около 60 видов, большинство из Палеарктики. В Казахстане найдено 12 видов. Биология: Самки устраивают гнезда (обычно одноячейковые) в земле, причем одни виды – до охоты, другие – после. Добыча – саранчовые (Acrididae). В каждой ячейке, как правило, запасается один экземпляр добычи. Некоторые виды известны как
эффективные регуляторы численности вредных саранчовых.
Литература: Гуссаковский, 1930; Казенас, 1968, 1969; Kohl, 1890; 1895; Tsuneki, 1963; Yasumatsu, 1938.
Определительная таблица видов рода Prionyx
1 (12). Коготки у основания с 2 зубцами.
2 (7). Тергумы брюшка у заднего края с бледно-желтыми перевязями.
3 (6). Длина тела более 18 мм. 1-я и 2-я субмаргинальные поперечные жилки у радиальной жилки не сближенные. Стебелек брюшка приблизительно равен 1-му и 2-му членикам жгутика вместе взятым.
4 (5). Шипы на ногах бледно-желтые, ноги бурые. Среднеспинка с косыми морщинами и точками между ними. Светлые перевязи брюшка широкие, по бокам включают по одному пятну оснолвной окраски. Стебелек брюшка у ♀ короче 1-го членика задних лапок, у ♂ такой же длины, как 1-й членик задних лапок. Брюшко у ♀, кроме светлых перевязей, желтовато-красное, у ♂, кроме светлых перевязей, черное. ……………………………....P. haberhaueri (Rad.)
5 (4). Шипы ног большей частью так же окрашены, как ноги. Среднеспинка густо пунктированная, без морщин. Полосы брюшка неширокие, впереди по бокам немного выемчатые. …………………..
…………….………………………………..P. nigropectinatus (Tasch.)
6 (3). Длина тела менее 18 мм. 1-я и 2-я кубитальные поперечные жилки у радиальной жилки отчетливо сближены. Стебелек брюшка длиннее, чем 2-й и 3-й членики жгутика вместе взятые. …………... ……………………………………………..………….P. niveatus (Duf.)
7 (2). Брюшко без бледно-желтых перевязей.
8 (11). Крылья слабо затемненные, желтоватые.
9 (10). Лапки, кроме обычных шипов, с густыми крепкими щетинками. Волоски головы и груди черно-коричневые. Промежуточный сегмент с лохматым длинным опушением. ……...….P. crudelis (F.Sm.) 10 (9). Лапки только с шипами. Волоски головы и груди светлые. ………
……………………………………………….....P. subfuscatus (Dahlb.)
11 (8). Крылья черно-коричневые. Лапки, кроме обычных шипов, с маленькими и слабыми щетинками. Гребень передних лапок двойной: состоит из слабых и крепких щетинок. ………………………... ………………………………………………..P. radoszkowskyi (Kohl)
12 (1). Коготки у основания с 3-4 зубцами.
13 (14). Щитик плоский, без продольного вдавления. Волоски на срединной площадке промежуточного сегмента короче диаметра переднего глазка. Боковые поверхности промежуточного сегмента ♀ с продольным килем. …………………………P. lividocinctus (Costa)
14 (13). Щитик выпуклый, с продольным вдавлением в середине. Волоски на срединной площадке промежуточного сегмента длиннее диаметра переднего глазка. Боковые поверхности промежуточного сегмента ♀ без киля.
15 (16). Тергумы брюшка на конце с бледно-желтой перевязью, на 1-м тергуме она широко прерванная. 4-й членик челюстных щупиков заметно длиннее 6-го. Наличник ♀ посредине с вырезкой. 4-й и 5-й стернумы брюшка без густого опушения. 1-й членик передних лапок ♀ снаружи обычно с 9 длинными шипами. ……P. kirbyi (Lind.)
16 (15). Тергумы брюшка без бледно-желтых перевязей. 4-й членик челюстных щупиков короче 6-го или равен ему.
17 (18). Брюшко красное; если с черными пятнами, то лишь на 5-м и 6-м тергумах. Грудь с обильным серебристым опушением, скрывающим скульптуру. Срединная площадка промежуточного сегмента по бокам с голыми участками. …...……P. viduatus viduatus (Christ.) 18 (17). 2-4 последних сегмента брюшка черные.
19 (20). Стебелек брюшка более или менее красный. Грудь ♀ почти без прилегающего опушения. 4-й и 5-й стернумы брюшка ♂ с темнозолотистым опушением. Усики, ноги и стебелек брюшка относительно тонкие. Срединное поле промежуточного сегмента с почти одинаковыми по длине нежными волосками. Крылья слегка затемненные. Пунктировка среднеспинки крупная (точки в 3-4 раза уже переднего глазка). …….……………………….……P. songaricus Ev.
20 (19). Стебелек брюшка черный. Грудь ♀ с прилегающим опушением. 4-й и 5-й стернумы брюшка ♂ без опушения или с темно-бурым опушением. Усики, ноги и стебелек брюшка более толстые. Срединное поле промежуточного сегмента с разными или одинаковыми по длине волосками. Крылья почти не затемненные. Пунктировка среднеспинки более мелкая.
21 (22). 4-й членик челюстных щупиков несколько длиннее 6-го. Передний край наличника ♀ с очень слабой выемкой. Бока среднегруди с густым серебристым опушением. 4-й и 5-й стернумы брюшка ♀ без густого опушения. Срединное поле промежуточного сегмента с почти одинаковыми по длине волосками. …………………………
……….……………………………… P. viduatus argentatus (Mocsary)
22 (21). 4-й членик челюстных щупиков короче 6-го. Передний край наличника ♀ с заметной выемкой. Бока среднегруди с негустым серебристым опушением. 4-й и 5-й стернумы брюшка ♀ с густым темным опушением. Срединное поле промежуточного сегмента с разными по длине волосками. ……………………..P. nudatus (Kohl)
Cиноним: vittatus (Kohl), 1884 (Enodia)
Описание: Самка. Передний край наличника слегка дугообразно выемчатый или почти прямой (рис. 62а). Средняя часть лба (с усиковыми ямками) сильно вдавленная. Наличник выпуклый, с отогнутой, расширенной по бокам каймой, на всей поверхности с густым прилегающим латунно-блестящим опушением и длинными светлыми желтоватосерыми волосками. Глаза почти параллельные (едва заметно сближаются книзу). Усики, как на рис. 64б. Голова и грудь с густым светлозолотистым (латунно-блестящим) прилегающим опушением, скрывающим скульптуру. Среднеспинка в середине голая, с косыми негрубыми морщинами и редкими точками. Щитик сильно выпуклый, с продольным вдавлением в середине. На голых выпуклых участках видны густая микропунктировка и некрупные точки. Заднещитик равноменрно выпуклый, но без бугра или выступа в середине. Скульптура срединного поля промежуточного сегмента скрыта под густым полуприлегающим опушением. 1-я и 2-я радиомедиальные жилки спереди только незначительно сближенные. Стебелек брюшка (рис. 64г) короче 1-го членика задних лапок (рис. 64в) приблизительно на ½ длины 2-го членика. Коготки двухзубцовые. Мандибулы в базальной части желтовато-рыжие. Скапус и педицеллус рыжие. Голова и грудь на большей части черные. Тегулы рыжие. Ноги на большей части рыжие. Шипы ног беловатые. Тазики более или менее черные. 1-й членик передних лапок с 7 длинными тонкими бурыми щетинками и 2 беловато-рыжими плоскими широкими щетинками на наружном крае и 7-9 короткими беловатыми шипами. Брюшко красно-рыжее, с широкими беловато-желтыми перевязями, включающими по 2 буровато-рыжих пятна. Шипы ног беловатые. Коготки рыжевато-бурые. Крылья прозрачные, слегка буроватые, с коричневыми и рыжими жилками. Длина тела 23-27 мм.
Самец. Наличник равномерно выпуклый, без обособленной каймы, с дугообразно выступающим передним краем, слегка широко выемчатым в середине (рис. 64д). Лицо с длинными светло-серыми волосками и прилегающим опушением. Глаза сближаются книзу. Ширина головы примерно в 2,4 раза больше ширины лица в середине. Промежуток между задними глазками примерно в 1,1 раза уже, чем между одним из них и ближайшим глазом. Базальные жилки жгутика усиков удлиненные (рис. 64е). Переднеспинка с редкой мелкой пунктировкой, сетчатой микроскульптурой и светло-серым прилегающим опушением. Среднеспинка с косыми морщинами, крупными ячееобразными точками, светло-серыми длинными волосками и слабо развитым прилегающим коротким опушением. Щитик равномерно выпуклый, слегка продольно вдавленный в середине, с редкими мелкими точками, микропунктировкой и опушением, особенно в середине и по краям. Заднещитик сильно выпуклый, но без бугра или выступа. Срединное поле промежуточного сегмента довольно грубо поперечно морщинистое, с довольно густыми изогнутыми отстоящими волосками, по длине приблизительно равными диаметру переднего глазка, только по бокам с узкой голой зоной. Стебелек брюшка (рис. 64з) по длине почти равен 1-му членику задних лапок (рис. 64ж), примерно в 2 раза длиннее 2-го членика. Голова, грудь, усики и ноги черные. Брюшко черное, с дважды выемчатыми желтовато-белыми перевязями у заднего края. Крылья слегка коричневатые, с бурыми и коричневыми жилками. Длина тела 16-22 мм.
Ареал: Юго-Западная Азия, Средняя Азия, Казахстан.
Распространение в Казахстане: Предгорья Западного Тянь-Шаня – Белые Воды; низкогорья Киргизского хр. – окр. Подгорного; долина р. Или – бывш. пос. Или; вост. отроги Заилийского Алатау – р. Чарын; сев. предгорья Джунгарского Алатау – Кызыл-Агач; Зайсанская котловина – 40 км зап. Черняевки; Прииртышье – Семипалатинск.
Экология и биология: Вид встречается в пустынной зоне, обычно в невысоких горах с щебнисто-глинистой почвой и по долинам рек, в степной зоне – на участках с песчаной почвой. Образ жизни неизвестен. Самки, по всей вероятности, для гнезд роют норки в земле и охотятся на саранчовых.
Синонимы: albisectus Lepeletier et Serville, 1828 (Sphex); sjoestedti Cameron, 1908 (Sphex); curvilineatus Cameron, 1912 (Sphex); congoensis Berland, 1927 (Sphex); alluaudi Berland, 1927 (Sphex); chudeaui Berland, 1927 (Sphex).
Подвиды: marginatus (F.Smith), 1856 (Parashex) – Африка.
Описание: Передний край наличника с маленькой треугольной вырезкой (рис. 64и,к). Наличник сильно выпуклый, с широкой уплощенной отогнутой каймой, с густым серебристым , слегка желтоватым прилегающим опушением и длинными желтоватыми светло-серыми волосками. Низ лба (с усиковыми ямками) сильно вдавленный, верх лба с продольным срединным вдавлением в виде широкой борозды. Глаза явно сближаются книзу (рис. 64и). Ширина головы приблизительно в 2,7 раза больше ширины лица в середине лба. Промежуток между задними глазками в 1,15 раза меньше промежутка между одним из них и ближайшим глазом. 4-й членик нижнечелюстных щупиков заметно длиннее 6-го. Переднеспинка утолщенная, гладкая и блестящая, с редкими точками и редкой микропунктировкой. Опушение слабо развитое, но хорошо заметное у заднего края. Среднеспинка с довольно редкой мелкой пунктировкой и очень нежной неправильной микроскульптурой. Опушение слабо развитое, хорошо заметное лишь у тегул. Отстоящие волоски разные по длине, самые длинные в 2-2,5 раза длиннее диаметра переднего глазка. Щитик гладкий и блестящий, выпуклый, с широким продольным вдавлением в середине, с редкими точками, по краям с опушением. Заднещитик равномерно продольно выпуклый, гладкий и блестящий, с редкими некрупными точками, без срединного вдавления. Срединное поле промежуточного сегмента с пунктировкой и нежной поперечно-морщинистой скульптурой, более четкой в средней части, с отстоящими короткими и длинными светлыми волосками (короткие в 3 раза короче диаметра переднего глазка, длинные примерно равны ему). Бока среднегруди с давольно редкой пунктировкой и тенденцией к образованию нежных морщин, с довольно густой микропунктировкой. Опушение груди относительно слабо развитое, густое на плечевых буграх, близ средних тазиков и в задней части боков промежуточного сегмента. 1-й членик передних лапок с 9-10 длинными уплощенными бурыми щетинками на наружном крае. Коготки 3-4-хзубцовые. Стебелек брюшка (рис. 64л) по длине почти равен 1-му членику задних лапок вместе с ½ 2го членика. Мандибулы бурые, в базальной части красноватые. Голова, усики, грудь и ноги черные. Членики лапок более или менее буроватые. Стебелек брюшка черный. 1-й тергум, вершина 1-го стернума, 2-й, 3-й и 6-й чегменты брюшка красно-рыжие, отчасти буроватые. Тергумы, по крайней мере 2-4-й, с узкой желтовато-белой перевязью у заднего края. Длина тела 13-19 мм.
Самец. Наличник сильно выпуклый, без вырезки на переднем крае
(рис. 64м), с густым прилегающим серебристым опушением и длинными белыми волосками. Глаза сильно сближаются книзу. Ширина лица у переднего глазка примерно в 1,5 раза шире, чем на наличнике. Промежуток между задними глазками почти равен или слегка уже, чем промежуток между одним из них и ближайшим глазом. Базальные членики жгутика усиков не удлиненные (рис. 64и). Ширина головы почти в 2 раза больше ширины лица впереди переднего глазка. Верх переднеспинки с довольно редкой и не очень крупной пунктировкой. Скульптура, окраска и опушение тела почти как у самки. Срединное поле промежуточного сегмента с отстоящими светлыми волосками разной длины. Наиболее длинные из них примерно в 3 раза длиннее наиболее коротких. Стебелек брюшка слегка длиннее 1-го и 2-го члеников задних лапок. 3-й тергум обычно только по бокам красно-рыжий. 1-й тергум со светлыми узкими поперечными полосками по бокам. 7-й тергум целиком черный. Длина тела 11,5-16 мм.
Ареал: Южная и Средняя Европа, Африка, Юго-Западная Азия, Средняя Азия, Казахстан, Китай, Монголия.
Распространение в Казахстане: Долина р. Урал – Уральск, окр. Индерборского; бассейн р. Уил – Бабатай, Караулкельды; Рын-пески; Тургайская ложбина – Наурзум; Южное Предуралье – Берчогур, Актюбинск; Казахский мелкосопочник – 40 км юго-зап. Аркалыка, р. Ишим; Сев. Приаралье – 29 км сев.-зап. Аральска; пуст. Бетпакдала – Кокашок, 70 км сев.-зап. Фурмановки; Сев. Прибалхашье – 15 км юго-зап. Балхаша, Мынарал; долина р. Или – Учарал, Или (бывш.), 17 км сев.-зап. Баканаса, окр. Капчагая, Николаевка, 25 км сев. Чингильды; Прииртышье – Большая Владимировка, с. Щербакты, Семеновка, Долонь, Малая Владимировка; Зайсанская котловина – Каратал, 45 км южн. Каратогая, Черняевка, Приозерное; предгорья Джунгарского Алатау – Андреевка, пер. Архарлы; предгорья Заилийского Алатау – Алматы, Чемолган; Алакульская впадина – Маканчи, 25 км сев. Рыбачьего. Для Казахстана вид указывают Ф.Моравиц (Morawitz, 1891 – Рын-пески) и В.Л.Казенас
(1969, 1972, 1975).
Биология и экология: Пустынно-степной вид. Самки гнездятся преимущественно на участках с песчаной или супесчаной почвой. Гнездо одноячейковое. Добыча – крупные личинки саранчовых, в частности, представителей родов Chorthippus, Stauroderus, Oedipoda, Omocestus, Calliptamus, Dociostaurus, Notostaurus и др. В гнезде запасается 1-2 экземпляра добычи. В качестве врагов известны мухи родов Sphecapata, Hilarella, Paragusia и др. Взрослые осы питаются нектаром цветков различных видов растений, в частности крестовника, зизифоры, лука, кермека, вероники, дикой моркови, гипсофилы, солнцецвета, клоповника, молочая, подмаренника, кендыря, тамариска, солодки и др.
Синонимы: isselii Gribodo, 1880 (Priononyx); obliquestriatus Mocsary, 1883 (Enodia); graecus Mocsary, 1883 (Enodia).
Подвиды: oasis (Tsuneki), 1971 (Sphex) - Монголия; apakensis (Tsuneki), 1971 (Sphex) – Сев. Китай.
Описание: Самка. Наличник сильно выпуклый, с широкой сглаженной отогнутой каймой впереди, с глубокой выемкой в середине переднего края (рис. 65а,б). Середина лба (с усиковыми ямками) сильно вдавленная. Глаза явно сближаются книзу (рис. 65а). Ширина лица перед передним глазком приблизительно в 1,2 раза больше минимальной. Промежуток между задними глазками слегка (примерно в 1,1 раза) шире промежутка между одним из них и ближайшим глазом. Лицо с густым прилегающим латунно-блестящим опушением. Усик как на рис. 65в. Переднеспинка вверху в середине гладкая и блестящая, с редкой мелкой пунктировкой, с редкой микропунктировкой, почти без продольного углубления в середине, на большей части с довольно густым опушением. Среднеспинка с густой довольно крупной пунктировкой, в средней части гладкая и блестящая, с редкой микропунктировкой, по бокам и впереди с густой микропунктировкой и густыми короткими отстоящими волосками; кроме того, у боковых краев с густым прилегающим опушением. Длина отстоящих волосков примерно равна диаметру переднего глазка. Щитик на большей части плоский, гладкий и блестящий, с редкой мелкой пунктировкой и микропунктировкой, в середине не вдавленный. Срединное поле промежуточного сегмента с нежными косыми складками, без точек, по бокам почти голое, в середине с полуприлегающими волосками, длина которых равна диаметру глазков или слегка короче. Бока среднегруди с густым прилегающим опушением, только сзади голые. Бока заднегруди голые. Бока промежуточного сегмента с продольным килем. 1-й членик передних лапок с 5 длинными уплощенными бурыми щетинками на наружном крае. Коготки 4-хзубцовые. Стебелек брюшка (рис. 65г) примерно равен 1-му членику задних лапок. Голова, грудь, усики и ноги черные. Мандибулы в базальной половине буровато-рыжие, передний край наличника рыжеватый. Скапус спереди буроватый. Лапки к вершине рыжеватые. Короткие шипы ног светлые, беловатые. Брюшко на большей части рыжее. 4-5-й тергумы сверху и 6-й сегмент целиком черные. Тергумы с обесцвеченной буроватой каймой сзади. Стебелек брюшка темно-рыжий, буроватый. Крылья слегка коричневатые, с рыжевато-коричневыми жилками. Длина тела 15-20 мм.
Самец. Передний край наличника в середине слегка (иногда едва заметно) выемчатый, часто почти прямой (рис. 65д,е). Лицо с густым се- ребристо-белым прилегающим опушением. Передняя часть наличника голая. Ширина лица перед передним глазком примерно в 1,5 раза больше, чем на наличнике. Голова спереди шире лица на уровне передних глазков примерно в 2 раза. Базальные членики усиков, как на рис. 65ж. Промежуток между задними глазками приблизительно в 1,3 раза шире, чем между одним из них и ближайшим глазом. Стебелек брюшка длиннее 1-го членика задних лапок примерно на 1/3 длины 2-го. Скульптура, опушение и окраска тела почти как у самки. Наличник целиком черный, слегка буроватый на переднем крае. Усики черно-бурые. Стебелек брюшка черный; 1-й тергум и 2-й сегмент на большей части рыжие. Длина тела 13-17 мм.
Ареал: Южная Европа, Северная Африка, Юго-Западная Азия, Средняя Азия, Казахстан, Китай, Монголия.
Распространение в Казахстане: Пуст. Кызылкум – ур. Тентексай (пески Алкокулькум), 5-10 км вост. Чардары; южные предгорья хр. Каратау – 30 км юго-зап. ст. Арысь; низкогорья хр. Каржантау – 30 км южн. Ленгера; низкогорья хр. Каратау – 6 км юго-зап. Лугового, 18 км сев.-зап. Кантемировки; пуст. Мойынкум – 30 км южнее ст. Чу; долина р. Или – Аяк-Калкан, 17 км сев.-зап. Баканаса; низкогорья хр. Заилийского Алатау – пер. Курдай, Каскеленское ущ.; предгорья Джунгарского Алатау – Кзыл-Агач; предгорья Калбинского хр. – 8 км южнее Каменки.; Прииртышье – 8 км юго-вост. Большой Владимировки; Прикаспийская низменность – Ащисай близ пос. Кульсары; Сев. Приаралье – ст. Чокусу, 29 км сев.-зап. Аральска, 10 км севю-зап. и 10-15 км юго-зап. ст. Камышлыбаш.
Биология и экология: Пустынно- сухостепной вид. Обычен в лессовых предгорьях и низкогорьях, глинистых и песчаных пустынях. Самки гнездятся в земле и охотятся на саранчовых (в частности, Dociostaurus tartarus Uv.). Отмечено питание взрослых ос на цветках зизифоры, кермека, тамариска, молочая, мяты, подмаренника и других растений.
Синоним: eatoni (E.Saunders), 1910 (Sphex).
Подвид: lugens (Kohl), 1889 (Sphex) – Армения, Иран, Афганистан, Средняя Азия.
Описание: Самка. Наличник с поперечным дугообразным килем близ переднего края, с расширенной по бокам плоской каймой, перед килем вогнутый, с маленькой тупоугольной выемкой в середине переднего края (рис. 65з). Лицо с густым желтовато-белым прилегающим опушением; наличник с несколькими черными отстоящими волосками. Внутренние края глаз почти параллельные. Низ лба по бокам от усиковых ямок вдавленный. Лицо приблизительно в 2,2 раза уже головы (см. спереди). Промежуток между задними глазками примерно в 1,2 раза уже промежутка между одним из них и ближайшим глазом. Усики, как на рис. 65и. Верх головы, переднеспинка и среднеспинка с густым серебристым (отчасти латунно-блестящим) прилегающим опушением, скрывающим скульптуру. Щитик сильно выпуклый, с широким продольным вдавлением в середине, сзади с продольными морщинами. Заднещитик выпуклый, в виде 2 бугров, разделенных продольным углублением, с поперечными морщинами. Бока груди с морщинами, имеющими разное направление, с более или менее гладкими промежутками между ними. Дорсальная поверхность промежуточного сегмента с не очень грубыми поперечными морщинами, с полуприлегающими золотисто-бурыми волосками. Стебелек брюшка в 1,1-1,2 раза длиннее 2-го членика задних лапок, образует с 1-м тергумом острый угол (см. в профиль – рис. 65к). Ноги, кроме черных шипов, с нежными темными щетинками и волосками. Тело черное, отчасти бурое. Крылья с бурым затемнением, особенно темным близ конца маргинальной (радиальной) ячейки. Усики и ноги черные, отчасти красновато-буроватые. Тазики, вертлуги и бедра с прилегающим серебристым опушением. Длина тела 25-30 мм.
Самец. Передний край наличника слегка широко выемчатый в середине (рис. 65л). Наличник вытянутый, сильно выпуклый, спереди вдавленнный, со сглаженной треугольной зоной впереди. Лицо (включая большую часть наличника) и верх головы с густым прилегающим серебристым (слегка желтоватым) опушением, скрывающим скульптуру. Наличник с редкими отстоящими черными волосками. Глаза почти параллельные (рис. 65л). Голова спереди почти в 2,3 раза шире минимального расстояния между глазами. Промежуток между задними глазками примерно в 1,1 раза шире промежутка между одним из них и ближайшим глазом. Переднеспинка сверху с продольным вдавлением. Переднеспинка и среднеспинка с густым прилегающим серебристым опушением, скрывающим скульптуру, и буроватыми длинными волосками. Щитик с продольным вдавлением, с продольными морщинами и ячееобразными точками. Заднещитик сильно выступающий в виде 2 бугров, разделенных довольно узким вдавлением. Срединное поле промежуточного сегмента с довольно грубой поперечно-морщинистой (отчасти ячеистой) скульптурой, с неправильной микроскульптурой, с отстоящими бурыми волосками. Бока среднегруди неправильно-ячеисто морщинистые. Стебелек брюшка почти в 2 раза длиннее 3-го членика задних лапок, но в 1,6 раза короче 1-го. Ноги, кроме шипов, с нежными волосками, без щетинок. 7-й стернум брюшка сзади по бокам угловато выступающий. Тело, ноги, усики черные. Мандибулы буровато-красные. Крылья с бурым затемнением, задние в апикальной части почти без затемнения. Длина тела 23-25 мм.
Ареал: Северная Африка, Юго-Западная Азия, Средняя Азия, Южный Казахстан.
Распространение в Казахстане: Пуст. Кызылкум; Присырдарьинская равнина – Кзыл-Орда, Джулек близ Чиили. Для Казахстана вид приводит В.В.Гуссаковский (1933 – «Перовск»).
Биология и экология: Редкий пустынный вид. Биология неизвестна. Самки, вероятно, гнездятся в более или менее рыхлом грунте и охотятся на крупных саранчовых.
Синонимы: ? dives Lepeletier, 1845 (Sphex); ? nivosus F.Smith, 1856 (Sphex); maracandicus Radoszkowski, 1877 (Podium).
Описание: Проксимальный зубец мандибул значительно крупнее среднего (рис. 66а). Передний край наличника без срединной вырезки, в середине прямой, дугообразно изогнутый по бокам (рис. 66б). Наличник поперечно выпуклый, с отогнутой широкой каймой, с густым прилегающим желтовато-белым опушением и косо отстоящими длинными белыми волосками. Глаза почти параллельные (рис. 66б), слегка расходятся книзу. Ширина лица в середине лба в 2,2 раза меньше ширины головы спереди. Промежуток между задними глазками слегка шире промежутка между одним из них и ближайшим глазом. Лицо до задних глазков в густом опушении. 1-й членик передних лапок с 11 длинными плоскими бурыми (на вершине обесцвеченными) щетинками и 11 короткими шиповидными рыжеватыми щетинками на наружном крае. Средние и задние лапки с рыжевато-бурыми шипами. Коготки с 2 зубцами. Переднеспинка без продольного вдавления в середине, с густым прилегающим опушением. Среднеспинка с густым прилегающим опушением, под опушением с густой микропунктировкой и редкой мелкой пунктировкой. Щитик равномерно выпуклый, с едва заметным продольным вдавлением в середине, с редкой мелкой пунктировкой и микропунктировкой, в середине блестящий, по краям с густым прилегающим белым опушением. Заднещитик в середине поперечно-бугровидно выпуклый, с густым прилегающим опушением почти на всей поверхности. Форма 2-й и 3-й субмаргинальных и маргинальной (радиальной) ячеек, как на рис. 66в. Срединное поле промежуточного сегмента с густым белым прилегающим опушением, почти целиком скрывающим поперечно-косую морщинистую скульптуру. Стебелек брюшка короткий (рис. 66г), почти в 2 раза короче 1-го членика задних лапок, почти равен по длине 2-му членику задних лапок (рис. 66д). Тело на большей части рыжее, голова сверху темно-бурая (почти черная), переднегрудь и среднегрудь сверху бурые. Брюшко рыжее, с желтовато-белыми дважды выемчатыми перевязями у заднего края сегментов, в выемках перевязей буровато-рыжие. Ноги рыжие. Шипы ног рыжие, на вершине лапок бурые. Длина тела 2532 мм.
Самец. Глаза слегка сближаются книзу (рис. 66е). Наличник сильно выпуклый, с гладкой голой каймой, слегка выемчатой в середине переднего края (рис. 66ж). Ширина головы (спереди) почти в 2,5 раза больше ширины лица в середине лба. 1-й членик жгутика длиннее скапуса (рис. 66з). Опушение и скульптура тела почти как у самки. Промежуток между задними глазками приблизительно в 1,1 раза шире промежутка между одним из них и ближайшим глазом. Щитик сильно выпуклый, слегка продольно вдавленный в середине. Стебелек брюшка, как на рис. 66и, короче 1-го членика задних лапок, но явно длиннее 2-го. Голова и грудь на большей части черные. Мандибулы красно-рыжие, с красно-бурой вершиной. Передняя часть наличника рыжая. Скапус бурый, отчасти рыжий. Жгутик усиков черно-бурый. Тегулы рыжие, отчасти бурые. Крылья прозрачные, жилки бурые и коричневые. Стебелек брюшка черный или бурый. 1-й тергум на большей части черно-бурый или бурый, с желто-рыжим пятном, по краям светло-желтый. 2-6-й тергумы с дважды выемчатыми беловато-желтыми перевязями у заднего края. Ноги черные или черно-бурые, с прилегающим белым опушением.
Коготки красно-рыжие.
Ареал: Африка, Юго-Западная Азия, Средняя Азия, Казахстан.
Распространение в Казахстане: Пуст. Кызылкум. В Зайсанской котловине (40 км зап. Черняевки) найдено 2 экземпляра ос, относящихся, вероятно, к этому виду, но значительно отличающихся деталями окраски от типичных представителей. Не исключено, что они принадлежат к самостоятельному виду. Для Южного Казахстана вид указывается в работах О.И.Радошковского (1877) и В.Л.Казенаса (1969).
Экология и биология: Пустынный вид. Встречается в песках. В кпачестве добычи в Северной Африке отмечена саранча Schistocerca peregrina Oliv. (Berland, 1956).
Синонимы: albopectinatus Taschenberg, 1869 (Enodia); ? suavis F.Morawitz, 1893 (Sphex).
Подвид: ettingol (Tsuneki), 1971 (Sphex) – Монголия.
Описание: Самка. Передний край наличника с неглубокой широкой дугообразной выемкой (рис. 66к,л) или почти прямой в середине. Лицо с густым серебристо-белым прилегающим опушением и длинными волосками. Глаза слегка расходятся вверх и вниз от середины лба (рис.
66к). Базальные членики усиков удлиненные (рис. 66м). Голова почти в 2 раза шире лица в середине его высоты. Промежуток между задними глазками приблизительно равен по ширине промежутку между одним из них и ближайшим глазом. Переднеспинка с редкой мелкой пунктировкой, очень нежной сетчатой неправильной микроскульптурой, с короткими прилегающими серебристо-белыми и длинными отстоящими белыми волосками. Среднеспинка в средней части голая, гладкая и блестящая, с редкими мелкими точками; по бокам, у переднего и заднего краев с густым прилегающим серебристым опушением и отдельными отстоящими белыми волосками. Щитик очень сильно выпуклый, на большей части голый, гладкий и блестящий, с отдельными мелкими точками, в середине продольно вдавленный. Срединное поле промежуточного сегмента с густыми нежными поперечными морщинами и нежной неправильной микроскульптурой, более или менее скрытыми под полуприлегающими волосками, длина которых не больше диаметра переднего глазка. Бока и низ груди с густым прилегающим опушением. 1-я и 2-я поперечные радио-медиальные жилки у радиальной жилки сближенные (рис. 67а). 1-й членик передних лапок с 8-10 длинными щетинками. Стебелек брюшка (рис. 67б) примерно в 1,5 раза короче 1-го членика задних лапок. Коготки двухзубцовые. Голова и грудь черные. Мандибулы (кроме черно-бурой вершины), передний край наличника, скапус отчасти (спереди), тегулы, ноги на большей части рыжие. Стебелек брюшка черный. Брюшко рыжее, тергумы с беловато-желтыми,дважды выемчатыми (по бокам) перевязями у заднего края; 2-5-й стернумы с прерванными в середине, выемчатыми по бокам светлыми перевязями. Тазики, вертлуги и основания бедер более или менее черные или черно-бурые. Крылья светлые, прозрачные, с бурыми и рыжевато-коричневыми жилками. Шипы средних и задних лапок в основном рыжевато-бурые или буровато-рыжие, у некоторых экземпляров светлые, беловатые, более или менее обесцвеченные. Длина тела 13-20 мм.
Самец. Передний край буроватой полупрозрачной каймы наличника слегка дугообразно выемчатый (рис. 67в). Глаза сильно сближаются книзу (рис. 67г). Промежуток между задними глазками приблизительно равен по ширине промежутку между одним из них и ближайшим глазом. Лицо с густым прилегающим серебристо-белым опушением и длинными белыми волосками. Переднеспинка с густым прилегающим опушением. Среднеспинка с шероховатой (нежной неправильноморщинистой) скульптурой, густой микропунктировкой, неправильной микроскульптурой и густым прилегающим опушением, отчасти скрывающим скульптуру. Щитик выпуклый, с продольным срединным вдавлением, с очень нежными продольными неправильными морщинами, с очень нежными точками, по краям опушенный. Срединное поле промежуточного сегмента с густыми поперечными нежными морщинами, микропунктировкой, нечеткой неправильной микроскульптурой и опушением. Стебелек брюшка (рис. 67д) длиннее 1-го членика задних лапок приблизительно на половину длины 2-го. Ноги с серовато-белым прилегающим опушением. Голова и грудь черные. Мандибулы у основания и на вершине бурые, в середине рыжевато-желтые. Передний край наличника буровато-рыжеватый. Усики черные, отчасти буроватые. Ноги черные, лапки буроватые. Шипы ног светлые, беловатые, отчасти обесцвеченные. Коготки буровато-рыжие. Стебелек брюшка черный. 1-й тергум рыжевато-бурый (иногда почти черный), остальные рыжие. 1-6-й тергумы с дважды выемчатыми спереди узкими, часто более или менее редуцированными перевязями; 5-й стернум с бурым краем. Стернумы брюшка желто-рыжие, 1-3-й – с узкими беловато-желтыми перевязями у заднего края (иногда также 6-й и 7-й). Длина тела 12-18 мм.
Ареал: Северная Африка, Юго-Западная Азия, Средняя Азия, Казахстан.
Распространение в Казахстане: Пуст. Кызылкум – 20 км южнее Яны-Кургана, 6 км юго-зап. Чардары; Присырдарьинская равнина – Байгакум (близ Чиили), Куваи (близ Казалинска), Баламурун, Тамды, Чардара; пески Большие Барсуки – Челкар; о. Барсакельмес; п-ов Мангышлак – ур. Бас-Кудук, Созды; низовья р. Чу; пуст. Мойынкум – 15 км сев.вост. Кенеса; Илийская долина – 8 км южнее Жаркента (бывш. Панфилов), окр. Аяк-Калкана, ур. Мын-Булак близ Аяк-Калкана, 45 км вост. пос. Чингильды, бывш. Или, Басший, 17 км сев.-зап. Баканаса; хр. Чулак – ущ. Челбар; горы Богуты, р. Чарын; пуст. Сарытаукум – 20 км сев. пос. Айдарлы; пуст. Бетпакдала; Сев. и Зап. Прибалхашье – Мын-Арал, Балхаш, 50 км сев. Балхаша; Зайсанская котловина – 30-40 км сев.-зап. Акжара, окр. Аксуата.
Биология и экология: Вид обитает в песчаных пустынях и полупустынях. В пустынной зоне встречается также по долинам рек (на песках). Самки делают одноячейковые гнезда в земле, охотятся на саранчовых, в частности, Dociostaurus tartarus Uv., Leptopternis gracilis (Ev.) и др.
Описание: Самка. Передний край наличника с вырезкой (рис. 67е). Наличник с густым прилегающим опушением, скрывающим скульптуру, и длинными отстоящими серебристо-серыми волосками. Опушение лба слабее развито, особенно вверху. Лоб с широкой срединной бороздой в верхней части, с редкой мелкой пунктировкой и густой микропунктировкой. Глаза явно сближаются книзу (рис. 67ж). Ширина лица перед передним глазком в 1,25 раза больше минимальной ширины на наличнике. 3-й членик усиков в 1,5 раза длиннее 4-го (рис. 67з). 4-й членик нижнечелюстных щупиков короче 6-го. Переднеспинка сверху гладкая и блестящая, с редкими мелкими точками и редкой микропунктировкой. Среднеспинка с густой некрупной пунктировкой, довольно сильно выпуклая, в середине без борозды, без прилегающего опушения, но с отстоящими волосками разной длины. Щитик выпуклый, гладкий и блестящий, с редкими мелкими точками, в середине слегка продольно вдавленный. Бока среднегруди с густой крупной пунктировкой, с нежными косыми морщинами сзади, у заднегруди, с пятном прилегающего серебристого опушения близ средних тазиков. Бока промежуточного сегмента вверху спереди с несколькими грубыми складками, сзади с пятном густого прилегающего опушения. Срединное поле промежуточного сегмента с мелкой пунктировкой и нежными поперечными морщинами и отстоящими волосками разной длины. (Самые длинные волоски в 2,5-3 раза длиннее диаметра переднего глазка.) 1-й членик передних лапок снаружи с 6-7 длинными шипами. Маргинальная, 2-я и 3-я субмаргинальные ячейки, как на рис. 67и. Стебелек брюшка длинный (рис. 67к), длиннее 1-го членика задних лапок на 1/3 длины 2-го. Коготки 4хзубцовые. Голова, грудь, усики и ноги черные. Мандибулы бурые, в середине красноватые. Лапки бурые, с буровато-рыжими или отчасти обесцвеченными светлыми шипами. Стебелек брюшка черный, следующие 3 сегмента красно-рыжие; 4-6-й – черные, 4-й часто у основания более или менее красно-рыжий; 4-й и 5-й стернумы с большим бурым пятном очень нежного вельветообразного опушения (рис. 67к). Крылья слегка буроватые, с бурыми и коричневыми жилками. Длина тела 13-16 мм.
Самец. Передний край наличника слегка изогнутый, в середине почти прямой (рис. 67л), иногда с едва заметной выемкой. Ширина лица перед передним глазком приблизительно в 1,5 раза больше минимальной ширины на наличнике (рис. 67м). Наличник с голой треугольной гладкой голой зоной впереди, с редкими некрупными точками, вверху и по бокам с густым прилегающим опушением и длинными светло-серыми волосками. Верх лба перед передним глазком со слабо развитым прилегающим опушением, с редкой мелкой пунктировкой и микропунктировкой. Длинные волоски приблизительно в 3 раза длиннее диаметра переднего глазка. Усики, как на рис. 67и. Промежуток между задними глазками почти равен по ширине промежутку между одним из них и ближайшим глазом или слегка меньше. Скульптура, окраска и опушение тела почти как у самки. Стебелек брюшка (рис. 67о) по длине почти равен 1-му и 2-му членикам задних лапок. 1-й и 2-й тергумы, вершина 1-го стернума и 2-й стернум красно-рыжие. 4-й и 5-й стернумы матовые, бурые, с очень нежным вельветообразным опушением. Длина тела 12-15 мм.
Ареал: Южная Европа, Северо-Восточная Африка, Юго-Западная Азия, Средняя Азия, Казахстан.
Распространение в Казахстане: Пуст. Кызылкум – 8 км зап. Чардары; долина р. Сыр-Дарьи – Соло-Тюбе близ Кзыл-Орды, Байгакум близ Чиили; п-ов Мангыщлак – Бейнеу, 25 км юго-вост. Таучика; Сев. Приаралье – Аманоткель, Камышлыбаш; предгорья хр. Каратау – Жамбыл; хр. Киндыктас – Суганды; Юго-Вост. Бетпакдала – 70 км сев.-зап. Фурмановки; низкогорья хр. Каржантау – 30 км южнее Ленгера; Зап. Прибалхашье – 5-10 км южнее Мынарала; Казахский мелкосопочник – 20 км юго-зап. Аркалыка, 10 км юго-зап. пос. Шийли, 10 км зап. пос. Ашутасты, 25 км сев.-вост. пос. Амангельды, 12 км сев. Атбасара, 20 км сев.вост. Державинска, 10-30 км сев.-вост. Киймы, Аягуз, Кургальджино, Баян-Аул, 10 км сев. Баян-Аула, 70-90 км юго-зап. Баян-Аула; долина р. Урал – 130 км сев. Гурьева (Атарау), Индерборский; Рын-пески; Южн.
Предуралье – 5-15 км сев.-вост. Актюбинска; предгорья Джунгарского Алатау – Андреевка; низкогорья Джунгарского Алатау – Копал, пер. Алтынемель, хр. Чулак; низкогорья и предгорья Заилийского Алатау – хр. Сюгаты (Согеты), пер. Курдай, 13 км сев. пос. Масанчи, Тургень, Талгар, 10 км сев.-зап. Чемолгана; долина р. Чу – окр. Фурмановки; Илийская долина – 65 км зап. Баканаса, Тескенсу, 17 км сев.-зап. Баканаса, Или (бывш.), 35 км сев.-зап. Капчагая, 10 км юго-зап. Жаркента (бывш. Панфилов); Алакольская впадина – Маканчи; Прииртышье – 7-12 км сев. и 8 км сев.-зап. Семипалатинска, 8 км юго-вост. Большой Владимировки, 9 км сев.-зап. пос. Долонь, Белогорье, Щербакты; Зайсанская котловина – 10 км сев.-вост. Каратала, 120 км вост. Зайсана, Приморское, 64 км южн. Аксуата; низкогорья Казахстанского Алтая – 30 км вост. пос. Алтай, Раздольное; Северо-Казахстанская равнина – 100 км зап. Павлодара.
Экология и биология; Пустынно-степной вид. Встречается во всех типах пустынь, полупустынь и степей, по долинам рек и в низкогорьях. Самки делают одноячейковые гнезда в земле и запасают в них саранчовых. Взрослые осы питаются нектаром цветков различных растений. Отмечено питание на тамариске, молочае, ак-курае, зизифоре, подмареннике, кермеке, кендыре, мордовнике, селитрянке, синеголовнике, веронике, доннике, мяте, гипсофиле, гречихе, морковнике и др. Самцы и самки ночуют открыто на растениях.
Синоним: ? melanotus (F.Morawitz), 1890 (Sphex)
Описание: Самка. Наличник с расширенной по бокам плоской каймой, сильно выпуклый, с редкими мелкими точками, нежной микроскульптурой, блестящий, с длинными черными волосками, без прилегающего опушения, впереди с неглубоким треугольным вдавлением, слегка тупоугольно выемчатым передним краем (рис. 68а). Глаза почти параллельные (рис. 68а). 1-й членик жгутика усиков удлиненный (рис. 68б). Промежуток между задними глазками слегка шире промежутка между одним из них и ближайшим глазом (или почти равен ему). Верх лба с узкой глубокой бороздой. Среднеспинка гладкая и блестящая, с довольно редкой (впереди по бокам густой) мелкой пунктировкой. Щитик с очень глубоким срединным продольным вдавлением, разделен на 2 сильно выпуклых бугра, гладкий и блестящий, с редкой мелкой пунктировкой вверху и впереди, сзади с продольными и косыми нежными морщинами. Заднещитик с неправильной морщинистой скульптурой и пунктировкой. Срединное поле промежуточного сегмента с густопоперечно-морщинистой скульптурой и нечеткой мелкой пунктировкой, с густыми бурыми длинными отстоящими волосками. Стебелек брюшка короткий, не длиннее 2-го членика задних лапок, расширяющийся кзади, образует с 1-м тергумом острый угол. 1-й членик передних лапок с 11 (?) щетинками и с волосками на наружном крае. Тело и придатки черные. Мандибулы в вершинной половине буроватые. Крылья с плотным бурым затемнением, с отдельными более светлыми участками, с черными и бурыми жилками. Длина тела 20-23 мм.
Самец. Наличник равномерно выпуклый, с широко, но неглубоко дугообразно выемчатым передним краем, с выступающими боковыми углами (рис. 68в), с широкотреугольной гладкой голой зоной впереди и очень узкой каймой, на большей части с густой пунктировкой (точки разной величины), с длинными отстоящими черными волосками. Глаза не очень сильно сближаются книзу. 3-7-й (8-й) членики жгутика с плакоидами (ограниченными килями), которые не уже ½ ширины члеников. Усики, как на рис. 68г,д. Среднеспинка с густой, довольно крупной пунктировкой. Точки в среднем в 4 раза уже переднего глазка. Промежутки между точками имеют тенденцию к образованию коротких, неправильно изогнутых морщин. Щитик сильно выпуклый, с широким, но неглубоким срединным вдавлением, с тонкой продольной складкой в середине (как у самки), с нежными продольными (по бокам косыми) морщинами в задней части. Скульптура, окраска и опушение тела почти как у самки. Стебелек брюшка (рис. 68е) почти в 2 раза короче 1-го членика задних лапок. Волоски головы и груди черные, отчасти буроватые. Крылья однообразно затемненные (бурые), с черными жилками. Тегулы сзади буровато-рыжие. Длина тела 16-20 мм.
Ареал: Средняя Азия, Иран (?), Южный Казахстан.
Распространение в Казахстане: Долина р. Или – Аяк-Калкан
(бывш.).
Экология и биология: Пустынный вид. В пустынной зоне встречается и по долинам рек. Биология неизвестна. Вероятно, самки гнездятся в плотном песчаном грунте и охотятся на саранчовых.
Вид не изучен автором.
Ареал: Иран, Средняя Азия, Южный Казахстан.
Распространение в Казахстане: Предгорья хр. Каратау – Байгакум (близ Чиили), Баламурун; пески Малые Барсуки, долина Сырдарьи. Экология и биология: Возможно, как у P. radoszkowskyi.
Синоним: tenuicornis F.Morawitz, 1890 (Sphex).
Описание: Самка. Наличник равномерно выпуклый, с широкой уплощенной каймой, с узкой выемкой в середине переднего края (рис. 68ж), с длинными серыми волосками и относительно слабо оразвитым прилегающим серым опушением. Срединный зубец мандибул маленький. Середина лица (с усиковыми ямками) сильно вдавленная. Глаза сближаются книзу (рис. 68з). Базальные членики усиков , как на рис. 68и. Ширина головы примерно в 2,6 раза больше ширины лица в середине. Промежуток между задними глазками слегка шире промежутка между одним из них и ближайшим глазом (примерно в 1,1 раза). Верх лба с глубокой и широкой бороздой. Лицо с мелкой, слабо заметной редкой пунктировкой и сетчатой микроскульптурой. Воротничок переднеспинки утолщенный, гладкий и блестящий сверху, с редкими крупными точками. Среднеспинка в середине сильгно выпуклая, по бокам, особенно сзади, вдавленная, с густой (в средней части редкой) крупной пунктировкой, без прилегающего опушения. Щитик сильно выпуклый, на большей части гладкий и блестящий, с редкими точками, в середине с широким продольным вдавлением. Срединное поле промежуточного сегмента широко уплощенно-вдавленное в середине, с довольно грубыми поперечными морщинами, с довольно густыми короткими отстоящими волосками (их длина примерно равна ½ диаметра переднего глазка), только сзади по бокам голое. Бока среднегруди с морщинистопунктированной поверхностью. Бока заднегруди и промежуточного сегмента с резкими косыми морщинами. Прилегающее опушение на груди почти не развитое. Бока среднегруди перед средними тазиками вдавленные. Стебелек брюшка (рис. 68к) немного длиннее 1-го членика задних лапок. 1-й членик передних лапок с 9 длинными тонкими уплощенными щетинками и 5 короткими шипами на наружном крае. Коготки 4хзубцовые. Голова и грудь черные. Мандибулы в середине краснорыжие. Скапус отчасти буроватый. Тегулы бурые, с черным пятном. Стебелек брюшка, 1-й тергум, 2-4-й сегменты брюшка рыжеватокрасные. Вершина брюшка черная. Ноги черные, отчасти буроватые. Коготки рыжевато-светло-коричневые. Вершины лапок рыжеватые. Крылья со светло-бурым равномерным затемнением. Жилки бурые и коричневые. Длина тела 14-17 мм.
Самец. Наличник равномерно выпуклый, его передний край в середине почти прямой (рис. 68л). Ширина лица перед передним глазком в 1,7 раза больше минимальной (на наличнике). Голова (см. спереди) почти в 2 раза шире лица перед передним глазком (рис. 68м). Наличник с узкой голой и гладкой каймой впереди, с редкими мелкими точками, довольно густой микропунктировкой, прилегающим светло-серым опушением (особенно вверху) и длинными отстоящими светло-серыми волосками. Базальные членики усиков, как на рис. 68н. Переднеспинка гладкая и блестящая, с редкими мелкими точками. Среднеспинка с крупной пунктировкой, редкой в середине и сзади, густой впереди и по бокам. Точки в 3-4 раза уже переднего глазка. (Промежутки между точками гладкие и блестящие.) Щитик выпуклый, в середине слегка продольно вдавленный, с редкими точками. Заднещитик равномерно поперечно выпуклый, с крупными поперечно-удлиненными точками. Срединное поле промежуточного сегмента в средней части с продольными, сзади поперечными, по бокам косыми и поперечными негрубыми сглаженными морщинами, с мелкой, довольно редкой пунктировкой и тонкими отстоящими светло-серыми волосками (они по длине примерно равны диаметру переднего глазка). Стебелек брюшка (рис 68о) слегка короче, чем 1-й и 2-й члениеки задних лапок, взятых вместе. Коготки 4хзубцовые. Голова, грудь, усики и ноги черные. Вершины лапок бурые. Стебелек брюшка бурый (иногда красный), на вершине рыжеватый. 1-3й тергумы, вершинная часть 1-го стернума, 2-й и 3-й стернумыы рыжевато-коричневые. Остальная часть брюшка черная. Крылья с бурым затемнением. Жилки бурые и темно-коричневые. Длина тела 12-16 мм.
Ареал: Юго-Западная Азия, Средняя Азия, Казахстан, Западный Китай.
Распространение в Казахстане: Пуст. Кызылкум – 5-8 км юго-зап.
и 8 км зап. Чардары; долина р. Или – 17 км сев.-зап. Баканаса, Каратуранга, Или (бывш.), 35 км сев.-зап. Капчагая.
Экология и биология: Обитает в глинисто-песчаных и песчаных пустынях. Встречается также на сухих лугах в поймах рек в пустынной зоне. Биология не изучена. Вероятно, самки устраивают одноячейковые гнезда в земле и охотятся на саранчовых. Отмечено питание взрочлых ос на ак-курае, селитрянке, молочае и тамариске.
Синонимы: soror Dahlbom, 1845 (Sphex); nigritus Lucas, 1848 (Sphex), nec Fabricius, 1878; desertorum Eversmann, 1849 (Sphex); subfuscatus Eversmann, 1849 (Sphex), nec Dahlbom, 1845; chrysopterus Ruthe et Stein, 1857 (Enodia); anthracinus A.Costa, 1867 (Gastrosphaeria).
Подвиды: ? nankumiensis (Laidlaw), 1929 (Sphex) – Индия; ? rhodesianus (Arnold), 1936 (Chlorion) – Родезия, Танзания; albovillosulus (Giordani Soika), 1942 (Sphex) – Сомали; ? rukwaensis (Arnold), 1959 (Sphex) – Танзания.
Описание: Самка. Наличник с глубокой полукруглой вырезкой на переднем крае (рис. 69а), с поперечными килями, отходящими от вырезки, широко вдавленный по бокам, с редкой пунктировкой, густой микропунктировкой, золотистым прилегающим опушением и длинными желтовато-серыми волосками. Лицо широкое. Голова (см. спереди) примерно в 2 раза шире лица в середине высоты. Внутренние края глаз почти параллельные, слегка расходятся книгу (рис. 69б). Базальные членики усиков, как на рис. 69в. Переднеспинка с нежными неправильными косыми морщинами и густой, нечетко выраженной мелкой пунктировкой, с шероховатой микроскульптурой. Щитик выпуклый, с мелкими редкими точками, густой микропунктировкой, сзади с нежными неправильно изогнутыми морщинами. Заднещитик сильно выпуклый, с поперечными неправильными морщинами, с неправильной шероховатой микроскульптурой, полуматовый. Срединное поле промежуточного сегмента с неправильной ячееобразной, по бокам нежной поперечно-морщинистой скульптурой, с шероховатой неправильной микроскульптурой и с длинными светло-коричневыми волосками. Бока груди с четкой морщинистой скульптурой, более или менее блестящие. 1-й членик передних лапок с 6 длинными бурыми щетинками на наружном крае. Форма 1-й и 2йсубмаргинальных ячеек как на рис. 69и. 1-й тергум образует со стебельком почти прямой угол (несколько менее 900). Стебелек короткий (рис. 69г), слегка короче 2-го членика задних лапок. Голова, грудь, брюшко, ноги и усики черные. Волоски головы и груди желтоватосерые. Крылья темно-желтые, с буроватой апикальной частью. Жилки рыжевато-коричневые. Длина тела 16-21 мм.
Самец. Наличник равномерно выпуклый, с плавной, едва заметной выемкой на переднем крае (рис. 69д), с густым прилегающим светлым опушением и довольно густыми отстоящими бурыми волосками (их длина примерно в 1,5 раза больше диаметра переднего глазка). Глаза относительно слабо сближаются книзу (рис. 69е). Базальные членики усиков, как на рис. 69ж. 3-6-й членики жгутика усиков с широкими плакоидами на передней поверхности. Промежуток между задними глазками примерно в 1,2 раза уже промежутка между одним из них и ближайшим глазом. Коготки 2-хзубцовые. Скульптура, опушение и окраска почти как у самки. Переднеспинка сверху выпуклая (рис. 69з). Морщины на срединном поле более четкие, чем у самки. Форма маргинальной и субмаргинальных ячеек, как на рис. 69и. Стебелек брюшка (рис. 69л) по длине равен 2-му членику задних лапок и ½ 3-го, в 1,5 раза короче 1-го, образует с 1-м тергумом почти прямой угол (рис. 69к). Голова, грудь, брюшко, усики и ноги черные. Волоски груди бурые. Крылья желтые, с буроватой вершинной частью, с рыжевато-коричневыми и желтокоричневыми жилками. Длина тела 13-19 мм.
Ареал: Южная и Средняя Европа, Северная Африка, ЮгоЗападная Азия, Средняя Азия, Казахстан, Китай, Монголия, Восточная Азия.
Распространение в Казахстане: Присырдарьинские пустыни – Сары-Агач, Мешеули (близ Чиили), Кзыл-Орда; пуст. Мойынкум – Каргалы-Куль, Акыр-Тобе; о. Барсакельмес, о. Возрождения (Аральское море); Рын-пески, пески Большие Барсуки – Челкар, Малые Барсуки, пуст. Кызылкум – Чардара; низкогорья хр. Каржантау – 30 км южн. Ленгера; предгорья хр. Каратау – Жамбыл; п-ов Мангышлак; предгорья и низкогорья Заилийского Алатау – Алматы, Тургень, Талгар, Алма-Арасан, Чемолган, ущ. Левый Талгар; пуст. Сарытаукум – 20 км сев. пос. Айдарлы; Илийская долина – Учарал, Каратуранга, 17 км сев.-зап. Баканаса, Баканас, Или (бывш.), Капчагай, Аяк-Калкан, низовья р. Чарын; предгорья Джунгарского Алатау – Талды-Корган; Алакульская впадина – 25 км сев. Рыбачьего; Зайсанская котловина – 40 км сев.-вост. пос. Кокпекты, 45 км южн. Каратогая, 40 км зап. Черняевки, 20 км вост. Каратала, 26 км юго-вост. Приозерного; Прииртышье – Щербакты, Канонерка, Большая и Малая Владимировки, Семеновка, пос. Алтай; Сев. и Зап. Прибалхашье – Балхаш, Мын-Арал; Тургайская ложбина – Наурзум; долина р. Урал – Уральск, Индерборский, Елтай, Гребенщиков; хр. Мугоджары – Берчогур, р. Шет-Иргиз.
Экология и биология: Пустынно-степной вид. В пустынной зоне обычен на сухих лугах речных долин и в низкогорьях. Самки гнездятся в земле и охотятся на саранчовых (зарегистрированы представители родов Calliptamus, Oedipoda, Celes, Chorthippus, Locusta и др.). Самцы и самки проводят ночь на растениях. Отмечено питание имаго на цветках тамариска, молочая, донника, мяты, кермека, селитрянки, крестовника, клоповника, вероники, зизифоры и других растений.
Синонимы: pubescens Fabricius, 1793 (Sphex); canescens Dahlbom, 1843 (Enodia); micans Eversmann, 1849 (Sphex); granti W.F.Kirby, 1900 (Sphex); platynotus Matsumura, 1912 (Sphex); ? zanoni Gribodo, 1925 (Priononyx); mocsaryi Kohl, 1885 (Sphex).
Подвиды: argentatus (Mocsary), 1883 (Enodia) – “sw. USSR”; pollens (Kohl), 1885 (Sphex) – Вост. Средиземноморье.
Описание: Самка. Наличник выпуклый, по бокам вогнутый, с почти прямым передним краем (рис. 70а) или с маленькой треугольной вырезкой в середине (рис. 70б,в), с густым прилегающим серебристобелым опушением и длинными белыми волосками. Лицо относительно узкое, его ширина в середине примерно в 2,7 раза меньше ширины головы (см. спереди). Глаза от середины лица слегка расходятся вниз и вверх (рис. 70а). Промежуток между задними глазками примерно в 1,1 раза шире промежутка между одним из них и ближайшим глазом. Усики как на рис. 70г. Среднеспинка с мелкой пунктировкой, редкой в средней части, более густой по бокам и у основания, без складок, с густым прилегающим серебристо-белым опушением и отстоящими волосками. Щитик сильно выпуклый, слегка продольно вдавленный в середине, с редкими мелкими точками, гладкий и блестящий, по краям с густым прилегающим опушением. Заднещитик поперечно выпуклый, гладкий и блестящий вверху, с густым опушением по краям. Срединное поле промежуточного сегмента с густыми одинаковыми по длине (равной примерно диаметру переднего глазка) полуприлегающими волосками, отчасти скрывающими скульптуру (неправильные морщины, грубая микроскульптура, нечетко ограниченные точки). Жилкование передних крыльев, как на рис. 70д. Коготки 4-хзубцовые. Бока груди и ноги с густым прилегающим серебристо-белым опушением. Стебелек брюшка (рис. 70е) по длине примерно равен 2-му членику задних лапок и 2/3 3го, но короче 1-го. Голова, грудь, усики и ноги черные. Стебелек брюшка черный (иногда бурый), остальная часть буровато-рыжая. 5-й и 6-й тергкмы часто с черными пятнами. Крылья прозрачные, с бурыми или коричневыми жилками. Отстоящие волоски головы и груди белые. Длина тела 17-24 мм.
Самец. Наличник выпуклый, почти без вырезки на переднем крае (рис. 70ж) или с едва заметной вырезкой, с густым прилегающим опушением и длинными белыми волосками (как все лицо до переднего глазка). Глаза сильно сближаются книзу (рис. 70з). Ширина головы (см. спереди) более чем в 2 раза больше, чем ширина лица перед передним глазком. Ширина промежутка между задними глазками почти равна ширине промежутка между одним из них и ближайшим глазом. Стебелек брюшка немного длиннее 1-го членика задних лапок. Скульптура, окраска и опушение тела почти как у самки. Усики к вершине буроватые. Тегулы сзади полупрозрачные, буроватые. Стебелек брюшка черный, по бокам более или менее красно-рыжий. 7-й и 8-й стернумы более или менее чкрно-бурые. Края сегментов с полупрозрачной каймой, иногда с очень узкой поперечной желтой, прерванной в середине перевязью перед каймой. Длина тела 15-20 мм.
Вид в Казахстане представлен 2 подвидами. Подвид P. v. argentatus (Mocs.) = P. v. mocsaryi (Kohl) отличается от типичного подвида окраской брюшка и более мелкими размерами тела. У самки 2-4-й тергумы, нижне-боковые части 4-6-го, задняя часть 1-го стернума, 2-5-й стернумы, срединное пятно на 6-м красно-рыжие. 4-6-й тергумы сверху и 6-й стернум почти целиком черные. У самца стебелек, 4-7-й тергумы черные, 4-й и 5-й стернумы рыжевато-бурые; 6-й и 7-й стернумы черные, остальная часть брюшка красно-рыжая. Длина тела самки 16-19 мм, самца – 14-17 мм. (Описание окраски и размеры тела типичного подвида приведены в описании вида).
Ареал: Южная и Средняя Европа, Африка, Юго-Западная Азия, Средняя Азия, Казахстан, Западный Китай, Монголия.
Распространение в Казахстане: P. v. viduatus - Пуст. Кызылкум – 5-8 км зап. Чардары, Присырдарьинские пустыни – Коксу, Туркестан; о. Барсакельмес; Приаралье – Чокусу, Камышлыбаш; пуст. Мойинкум; пов Мангышлак; пуст. Бетпакдала; Сев. Прибалхашье – г. Балхаш; долина р. Или – 17 и 65 км зап. Баканаса, 35 км сев.-зап. Капчагая, Аяк-Калкан, Каратуранга, Или (бывш.), Чингильды, 20 и 45 км вост. Чингильды, 8 км южн. Жаркента (бывш. Панфилов), низовья р. Курты, Егеньсу; пуст. Сарытаукум – 20 км сев. пос. Айдарлы; Казахский мелкосопочник – 170 км сев. Балхаша, 8 км юго-зап. Каркаралинска; 50-70 км юго-зап. БаянАула; Тургайская ложбина – 25 км зап. пос. Шийли (р. Каинды), 10-20 км сев.-вост. Амантогая; пески Большие Барсуки – Челкар; Прикаспийская низменность – Ащисай (близ пос. Кульсары); Прииртышье – 20 км сев.-зап. Майского, окр. Большой Владимировки; Зайсанская котловина – 72 км юго-вост. пос. Кокпекты.
P. v. argentatus – Пуст. Кызылкум, Присырдарьинские пустыни _ Джулек (близ Чиили); долина р. Или – Или (бывш.), Аяк-Калкан, 17 км сев.-зап. Баканаса, Тескенсу; п-ов Мангышлак; долина р. Урал – Индерборский; Уральск, Чинаров, Харькин, Елтай, Гребенщиков; хр. Мугоджары – Берчогур; Южн. Предуралье – Актюбинск; Рын-пески; Прибалхашье – Мын-Арал; Тургайская ложбина – 10 км юго-зап. пос. Шийли; Казахский мелкосопочник – 20 и 45 км сев.-вост. Державинска, 15 км сев.-вост. Амантогая, 20 км юго-зап. Аркалыка, 70-90 км юго-зап. и 10 км сев. Баян-Аула; Прииртышье – Белогорье, 9 км сев.-зап. пос. Долонь, Щербакты.
Экология и биология: Пустынно-степной вид. Встречается во всех типах пустынь, предпочитая участки с глинисто-песчаной почвой. В степной зоне чаще встречается на участках с песчаной почвой. Обычен на открытых участках по долинам рек. Самки делают одноячейковые гнезда в земле. По наблюдениям на полуострове Мангышлак, основной добычей там являются самки атбасарской кобылки Dociostaurus kraussi Ingen. Сначала оса охотится, затем роет норку, поместив добычу на низкое растение. Втащив кобылку в гнездо, откладывает на нее яйцо и закрывает норку. Личинка, вышедшая из яйца, съедает кобылку за 7-8 дней, делает кокон и в нем зимует. Ночуют взрослые осы на растениях. Отмечено питание их на цветках гораниновии, гелиотропа, молочая, аккурая, тамариска, лука, селитрянки, кермека, клоповника, качима, донника, мяты, синеголовника, зизифоры, крестовника, мордовника, полыни, вероники, тимьяна и других растений.
Диагностические признаки: Усиковые ямки расположены вблизи середины лица и обычно отделены промежутком от лобно-
наличникового шва. Усики самцов без плакоидов. 1-й членик жгутика усиков почти всегда длиннее 2-го. Наличник обычно широкий. Мандибулы с 1-3 зубцами на внутреннем крае, редко без зубцов. Мандибулярная ямка обычно закрытая парамандибулярным выступом гипостомы. 3й членик максиллярных пальп симметричный. Гипостомальный киль полный. Затылочный киль внизу прерванный. Воротничок – переднеспинки от короткого до длинного. Срединное поле промежуточного сегмента обычно имеется и обозначено другой скульптурой. Дыхальцевая борозда отсутствует. Эпистернальный шов обычно длинный, доходит или почти доходит до передне-вентрального края плеврона. Передние крылья обычно с 3 субмаргинальными ячейками. Обе возвратные жилки впадают, за редким исключением, во 2-ю субмаргинальную ячейку. Самка с псаммофором. Средние тазики сближенные, часто соприкасаются. Две апикально-вентральные щетинки 5-го членика задних ног уплощенные, разделены промежутком, в 3 и более раз более широким, чем щетинки (редко в 2 раза). Внутренний край коготков простой или с 1-2 зубцами у основания. Лапки без плантул (у Parapsammophila обычно с ними). Стебелек брюшка состоит из 1-го стернума, у Ammophila также и из 1-го тергума. 8-й тергум без церок. Вольселла с подвижно сочлененным дигитусом.
В трибе 6 родов, из которых в фауне Казахстана представлены 4: Parapsammophila, Eremochares, Podalonia и Ammophila.
Типовой вид – P. miles (Taschenberg), 1869 [ = Ammophila cyani- pennis Lepeletier, 1845].
Cиноним: Ceratosphex Rohwer, 1921
Диагностические признаки: Внутренние края глаз прямые, сближаются книзу у самца и почти параллельные у самки. Лобная линия слабая или отсутствует. Вершинные членики усиков иногда с тилоидами. Усиковые ямки отделены от наличника промежутком, ширина которого не меньше диаметра ямки. Наличник впереди у самки с прямой на большей части срединной лопастью, у самца закругленной или усеченной. Мандибулы с 1-2 зубцами на внутреннем крае у самки и с 1-4 зубцами у самца. Ротовые органы обычно короткие. Мандибулярные ямки закрытые у самки и открытые у самца. Воротничок короткий, с почти вертикальной передней поверхностью. Стебельковая ямка вентрально широко окаймлена стернумом промежуточного сегмента. Мезостернальная область иногда вогнута для прикладывания передних тазиков. Эпистернальная ьорозда от субалярной ямки идет прямо вниз, не пересекая скробальную ямку. Маргинальная ячейка на вершине закругленная. 2-я возвратная жилка изредка впадает в 3-ю субмаргинальную ячейку (P. algira). 1-я или 2-я возвратные жилки иногда интерстициальные. Передняя лапка самки с гребнем длинных, иногда уплощенных щетинок. Членики лапки на вершине удлиненные, особенно 1-й. Средние тазики соприкасаются друг с другом. Средние голени обычно с 2 шпорами. Лапки обычно с плантулами. Коготки обычно с 2 базальными зубцами. Стебелек брюшка состоит только из 1-го стернума, который соприкасается со 2-м стернумом. Дыхальца 1-го тергума расположены у середины тергума. Гоностиль самца не раздвоенный на вершине.
Биология не известна. Самки гнездятся в земле.
Род распространен в Старом Свете. Известно около 20 видов, большинство – из Африки. В Казахстане 1 вид.
Литература: Kohl, 1906; Menke, 1966
Определительная таблица видов рода Parapsammophila
1 (1). Среднегрудь впереди закругленная, без бугровидного выступа. Членики передних лапок на конце с длинным выступом. Ноги отчасти светло-красные. Большая часть головы и груди с густым снежно-белым прилегающим опушением……..…P. turanica F.Mor.
Синоним: lutea auct., non Taschenberg, 1869
Описание6 Самка. Передний край срединной лопасти наличника ограничен по бокам угловатыми выступами, слегка, почти равномерно дугообразно выпуклый (рис. 71а), иногда в середине слегка выемчатый. Внутренние края глаз почти параллельные (рис. 71б). Лицо широкое (рис. 71б). Промежуток между задними глазками слегка уже промежутка между одним из них и ближайшим глазом. Переднеспинка равномерно выпуклая, без продольного углубления. Воротничок примерно в 2 раза шире своей длины. Среднеспинка с неравномерной мелкой пунктировкой, впереди и по бокам в передней половине с густой пунктировкой, иногда с тенденцией к образованию поперечных моорщин, в середине и в задней половине с более редкой пунктировкой, с гладкими блестящими промежутками. Прилегающее опушение головы и груди хорошо развитое. Светлые длинные отстоящие волоски довольно густые. Щитик выпуклый, в середине продольно вдавленный, на большей части гладкий и блестящий, с редкими точками, по краям с густым прилегающим опушением. Срединное поле промежуточного сегмента с густыми поперечными (слегка скошенными) морщинами, в средней части с коротким прилегающим опушением и длинными отстоящими белыми волосками, по бокам голое, с правильными негрубыми морщинами. 1-4-й членики передних лапок асимметричные, с длинными крепкими щетинками. Проксимальный шип на вершине выступа 1-го членика более толстый и короткий, чем остальные. Среднегрудь снизу впереди без бугра. 2-я возвратная жилка впадает во 2-ю субмаргинальную ячейку близ начала 2-й радиомедиальной жилки (рис. 71в). 1-й членик стебелька брюшка слегка короче 1-го и 2-го члеников задних лапок . Голова и грудь черные. Мандибулы на большей части рыжевато-желтые. Наличник (кроме верхнебоковых частей) красно-рыжий. Усики черные, только скапус краснорыжий. Тегулы рыжие, жилки крыльев рыжевато-коричневые, у основания крыла рыжие. Крылья не затемненные. Брюшко красно-рыжее, в вершинной части буроватое. Ноги рыжие. Длина тела 20-27 мм.
Самец. Передний край наличника без вырезки в середине и без боковых зубцов (рис. 71г). Глаза сильно сближаются книзу (рис. 71д). Ширина лица перед передним глазком в 1,6-1,7 раза больше ширины внизу (на наличнике). Промежуток между задними глазками примерно в 1,1 раза шире промежутка между одним из них и ближайшим глазом. Лицо с густым серебристым прилегающим опушением и длинными белыми отстоящими волосками. Переднеспинка равномерно выпуклая, без продольного углубления. Ширина воротничка примерно в 1,7 раза больше его длины. Среднеспинка с густой мелкой пунктировкой, в задней поло- вине более редкой, в передней – более густой, так что промежутки, сливаясь, образуют здесь неправильные складки. Опушение и волосяной покров головы и груди почти как у самки. Щитик выпуклый, в середине слегка продольно вдавленный. Срединное поле промежуточного сегмента у основания выпуклое, на всей поверхности с довольно густыми поперечными (слегка косыми) морщинами, на большей части с короткими прилегающими и длинными отстоящими волосками, по бокам с голыми участками, покрытыми правильными негрубыми морщинами. Низ и бока груди и тазики с густым прилегающим серебристо-белым опушением и длинными косо отстоящими белыми волосками. Окараска почти как у самки. Наличник черный, с рыжевато-бурым передним краем. Усики черные, только вершина скапуса рыжевато-бурая. Брюшко краснорыжее, в вершинной части затемненное. 1-й членик стебелька черный, 2й сверху черный, но по бокам рыжевато-бурый. Ноги рыжие, кроме большей части тазиков и задних вертлугов. Длина тела 20-25 мм.
Ареал: Северная Африка, Юго-Западная Азия, Средняя Азия, Южный Казахстан.
Распространение в Казахстане: Присырдарьинские пустыни – Баламурун (близ Чиилит), Джулек (близ Чиили); пуст. Кызылкум; долина р. Или – 35 км сев. Чилика (Аяк-Калкан), 50 км сев.-вост. Чилика (15 км вост. Аяк-Калкана).
Экология и биология: Обитает в песчаных пустынях. Самки гнездятся в плотном песке. Добыча неизвестна, возможно, относится к прямокрылым.
Типовой вид – Ammophila violaceipennis Lepeletier, 1845.
Синоним: Psammophila Dahlbom, 1842.
Диагностические признаки: Внутренние края глаз прямые, обычно параллельные у самок и сближающиеся книзу у самцов. Лобная линия вдавленная или отсутствует. Скапус овальный. 1-й членик жгутика длиннее, чем 2-й. Вершинные членики у самцов часто с тилоидами.
Усиковые ямки у самок почти соприкасаются с наличником, у самцов отделены промежутком, равным примерно диаметру ямок. Передний край наличника самок с широкой, усеченной срединной лопастью, ограниченной по бокам острыми углами, или дугообразно изогнутый. Передний край наличника самцов прямой или закругленный, иногда слегка широко выемчатый. Верхняя губа обычно шире своей длины. Мандибулы самок с 1 большим зубцом в середине внутреннего края, мандибулы самцов тоже с 1зубцом на внутреннем крае. Ротовые органы длинные. Мандибулярные ямки закрыты гипостомой у самок и некоторых самцов. Воротничок переднеспинки короткий. Срединное поле промежуточного сегмента имеется или не выражено. Стебельковая впадина широко окаймлена вентрально стернумом промежуточного сегмента. Эпистернальная борозда идет почти прямо вниз от субалярной ямки, минуя скробальную ямку. Маргинальная ячейка не верщине заостренная или притупленно-закругленная. Форма 2-й субмаргинальной ячейки варьирует. Передние лапки самок с гребнем шипов или щетинок, наружные вершины члеников лапок обычно удлиненные. Внутренняя вершина передних тазиков иногда с зубцом или шипом. Средние тазики сближенные. Коготки обычно простые, но иногда с базальным зубцом на внутреннем крае. Аролиум у самок иногда более или менее редуцированный. Стебелек брюшка состоит только из 1-го стернума, вершина которого достигает 2-го стернума. Стебелек образует с 1-м тергумом (см. в профиль) угол от45 до 900. 1-й тергум сильно расширяющийся к вершине, особенно у самок; его дыхальца расположены обычно перед его серединой. Боковые стороны 8-го стернума сближаются к вершине, которая обычно имеет выемку. Вальва пениса самцов обычно на вершине с зубчиками на вентральном крае. Гоностиль не раздвоенный.
Биология: Самки делают простое одноячейковое гнездо в земле. Охотятся обычно перед изготовлением гнезда. Добыча – гусеницы бабочек (Lepidoptera), главным образом сем. Noctuidae. В каждом гнезде запасается одна гусеница.
Род имеет всесветное распространение. Известно около 70 видов. В Казахстане найдено 8 видов.
Литература: Гуссаковский, 1930; Казенас, 1968; Мярцева, 1963б; Kohl, 1906; Roth, 1928, 1929; Tsuneki, 1978b.
1 (10). Срединное поле промежуточного сегмента голое, морщинистое.
Передние тазики на вершине с шипом, иногда очень коротким.
2 (7). Брюшко с красным основанием.
3 (6). Коготки у основания без зубчика.
4 (5). Чернаяокраска брюшка с синим металлическим блеском. ………...
…...………………………………………………….P. atrocyanea (Ev.)
5 (4). Черная окраска брюшка без синего металлического блеска. ………
……………………………………………………….....P. alpina (Kohl)
6 (3). Коготки у основания с зубчиком. ……………….……..P. affinis (K.)
7 (2). Брюшко сплошь черноею Коготки у основания без зубчиков или с очень маленьким зубчиком.
8 (9). Брюшко с синим металлическим блеском. Среднеспинка с косыми морщинами. …………………….………………….P. atrocyanea (Ev.)
9 (8). Брюшко без синего металлического блеска. Среднеспинка без морщин. ……………………………….…………….P. ebenina (Spin.)
10 (1). Срединное поле промежуточного сегмента сплошь покрытое волосками. Передние тазики без шипа.
11 (16). Аролии хорошо развитые. Волоски груди целиком или частично светлые. Предпоследний членик передней лапки ♀ не симметричный.
12 (13). Среднеспинка с нежной и редкой пунктировкой, промежутки между точками не уже точек. Опушение среднеспинки короткое (волоски короче, равны или слегка длиннее диаметра переднего глазка). ……………………………………………...……P. fera (Lep.)
13 (12). Среднеспинка с более густой и более крупной пунктировкой, промежутки между точками уже точек. Волоски на среднеспинке значительно длиннее диаметра переднего глазка.
14 (15). Бока среднегруди с прилегающим опушением. ….P. tydei (Guill.)
15 (14). Бока среднегруди без прилегающего опушения.…P. luffi (Saund.)
16 (11). Аролии почти не развитые. Волоски груди черные. Предпоследний членик передней лапки ♀ симметричный………………………
……………………………………………………….. P. hirsuta (Scop.)
Синонимы: lutaria auct., non Fabricius, 1787; ariasi Mercet, 1906 (Ammophila).
Описание: Самка. Передний край наличника широко дугообразный, без зубцов (рис. 71е). Наличник с редкой мелкой пунктировкой, с микропунктировкой и очень нежной сетчатой микроскульптурой, впереди более или менее сглаженный, с длинными отстоящими волосками, без прилегающего опушения. Внутренние края глаз прямые и параллельные (рис. 71ж). Промежуток между задними глазками приблизительно в 1,6 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка с редкой мелкой пунктировкой, гладкая и блестящая. Среднеспинка с редкой мелкой пунктировкой, гладкая и блестящая, с очень нежной сетчатой микроскульптурой. Щитик выпуклый, гладкий и блестящий, с редкими мелкими точками, в середине продольно вдавленный, сзади с нежными продольными морщинами. Заднещитик сильно выпуклый, слегка продольно вдавленный в середине, с мелкими редкими точками и морщинами. Срединное поле промежуточного сегмента голое, с косыми поперечными морщинами, с неправильной микроскульптурой. Передние тазики с маленьким шипом сзади на внутреннем крае (рис. 71з). Коготки с зубчиком (рис. 71и). 1-й членик передних лапок с 7-8 крепкими щетинками, из которых проксимальная на вершине апикального выступа короче и шире остальных (рис. 71к). Лапки с пульвиллами. Стебелек брюшка в 1,3-1,4 раза короче 1-го членика задних лапок, почти в 1,2 раза длиннее 2-го. Голова, грудь, ноги, усики, стебелек и вершина брюшка черные. Мандибулы, лапки и тегулы более или менее бурые. 1-й тергум, вершинная часть 1-го стернума, 2-й сегмент и большая часть 3-го красно-рыжие. Волоски головы и груди черные. Крылья буроватые, задние светлее. Жилки коричневые и бурые. Коготки буровато-красные. Длина тела 14-21 мм.
Самец. Передний край наличника очень слабо широко выемчатый, почти прямой (рис. 71л). Глаза сильно сближаются книзу (рис. 71м). Наличник и лоб с прилегающим серым опушением и длинными отстоящими волосками. Промежуток между задними глазками в 1,3-1,4 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка и среднеспинка с довольно густыми мелкими, слегка продольно удлиненными точками и сетчатой нежной микроскульптурой. Щитик на всей поверхности с нежными продольными морщинами, впереди более или менее сглаженный, с густой пунктировкой. Скульптура заднещитика, срединного поля промежуточного сегмента и боков среднегруди и промежуточного сегмента, как у самки. Стебелек брюшка по длине примерно равен 1-му членику задних лапок и почти в 2 раза длиннее 2-го. Окраска тела почти как у самки. Волоски головы и груди черно-бурые и серые. Вершина 1-го стернума и 1-го тергума, 2-й и 3-й сегменты брюшка красно-рыжие. Крылья буроватые. Жилки коричневые и бурые. Длина тела 13-20 мм.
Ареал: Почти вся Европа, Северная Африка, Юго-Западная Азия, Средняя Азия, Казахстан, Китай, Монголия, Сибирь, Дальний Восток России.
Распространение в Казахстане: П-ов Мангышлак, плато Устюрт, Рын-пески, долина Урала; долина Сыр-Дарьи, присырдарьинские пустыни, пуст. Кызылкум, предгорья и низкогорья хр. Каратау, Киргизский хр., Таласский Алатау, низкогорья и предгорья Заилийского Алатау, предгорья и низкогорья Джунгарского Алатау, хр. Кетмень, Кунгей Алатау, Южное Прибалхашье и долина р. Или, пуст. Сексеулдала, Казахский мелкосопочник, Тургайская ложбина, Прииртышье, Казахстанский Алтай, Зайсанская котловина, низкогорья и предгорья хр. Саур, хр. Тарбагатай, Североказахстанская равнина , т.е. в Казахстане почти повсеместно.
Экология и биология: Эврибионтный вид. Встречается от лесной зоны до пустынь, в горах полнимается до субальпийских лугов. Предпочитает биотопы степного характера. Самки охотятся на гусениц совок (Noctuidae), живущих в земле. Гнездо устраивается до охоты. Оно имеет только одну ячейку, в которую помещается одна гусеница.
Описание: Самка. Передний край наличника с маленькой треугольной выемкой в середине (рис. 72а). Промежуток между задними глазками почти в 2 раза уже промежутка между одним из них и ближайшим глазом. Голова и грудь с черными отстоящими волосками, с редкой пунктировкой и сетчатой морщинистой микроскульптурой. 1-й членик жгутика только незначительно длиннее, чем 2-й, почти равен 3му. Лицо квадратное. Бока среднегруди почти без морщин. Промежуточный сегмент впереди с нежной косой морщинистой скульптурой. Лапки с отчетливыми пульвиллами. Радиальная (маргинальная) ячейка на конце закругленная, сравнительно короткая. Щетинки гребня передних лапок самки короткие, асимметрия 2-4-го члеников слабая. 1-й членик с выступом на конце. Коготки без зубца. Передние тазики без шипа. Стебелек брюшка короче, чем у P. affinis. Тело черное. 1-й, 2-й и передняя половина 3-го тергума красно-рыжие. Крылья слабо затемненные, буроватые. Длина тела 10-13 мм.
Самец. Наличник удлиненный, кпереди сильно суженный, со слегка дугообразно выемчатым передним краем, с закругленными переднебоковыми углами (рис. 72б). Наличник с густым прилегающим серебристо-серым опушением и длинными светло-серыми и черно-бурыми волосками. Верх лба и темя с длинными черными волосками (их длина в 34 раза больше диаметра переднего глазка). Глаза сближаются книзу (рис. 72б). Ширина лица перед передним глазком в 1,5 раза больше минимальной ширины лица на наличнике. Промежуток между задними глазками в 2 раза уже промежутка между одним из них и ближайшим глазом. Среднеспинка с редкой мелкой пунктировкой и густой сетчатой микроскульптурой. Щитик с такой же скульптурой и негрубыми продольными морщинами в задней половине. Срединное поле промежуточного сегмента голое, с очень нежной ячеисто-морщинистой скульптурой (с тенденцией к преобладанию косых морщин). Грудь с черно-бурыми и бледно-серыми длинными волосками. Маргинальная и субмаргинальные ячейки, как на рис. 72в. Стебелек брюшка примерно в 1,2 раза короче 1го членика задних лапок и в 1,5 раза длиннее 2-го. Пульвиллы развитые. Коготки без зубца. Голова, грудь, усики и ноги черные. Крылья слегка буроватые, апикальнее маргинальной ячейки несколько темнее. Стебелек брюшка черный. 1-й тергум, 2-й сегмент и базальная часть 3-го сегмента красно-рыжие. Коготки красно-рыжие. Длина тела 10-14 мм.
Ареал: Средняя и Южная Европа, Марокко, Юго-Западная Азия, Кавказ, Таджикистан, Юго-Восточный Казахстан, Кыргызстан.
Распространение в Казахстане: Заилийский Алатау – ущ. Правый Талгар, 2000 м над ур. м.
Биология и экология: Горный вид, встречающийся в сухолуговых биотопах. Биология неизвестна. Вероятно, самки роют норки в земле и устаивают в них простые одноячейковые гнезда, в которых запасают гусениц ночниц (Noctuidae).
Синоним: ? psilocera Kohl, 1888 (Ammophila)
Описание: Самка. Передний край наличника дугообразно выступающий, без зубцов и выемок (рис. 72г). Наличник слабо выпуклый, в середине уплощенный, с неравномерной, в целом густой пунктировкой и очень нежной сетчатой микроскульптурой. Точки разной величины (в 38 раз уже переднего глазка). Внутренние края глаз почти параллельные (рис. 72д). Ширина головы (см. спереди) в 1,75 раза больше ширины лица в середине. 1-й членик жгутика усиков почти в 2 раза длиннее 2-го. Промежуток между задними глазками почти в 1,7 раза уже промежутка между одним из них и ближайшим глазом. Лицо без прилегающего опушения, с длинными отстоящими черными волосками. Лоб вверху и темя уплощенно-вдавленное. 2-4-й членики передних лапок слабо асимметричные. 1-й членик с 8 уплощенными щетинками. 1-я щетинка на апикальном выступе укороченная, шипообразная, утолщенная и изогнутая. Переднеспинка вверху в середине гладкая и блестящая, с редкой мелкой пунктировкой, по бокам и впереди с более или менее выраженными нежными поперечными морщинами. Среднеспинка с косыми нежными морщинами и мелкой густой (в средней части и сзади редкой) пунктировкой, с четкой двойной бороздой в середине. Бока среднегруди с косо- (отчасти ячеисто-) морщинистой скульптурой. Щитик сильно выпуклый, впереди сглаженный, с редкими мелкими точками, сзади с нежными продольными морщинами, в середине слегка продольно вдавленный. Заднещитик сильно выпуклый, с нежными косыми морщинами. Срединное поле промежуточного сегмента голое, с очень нежной сетчато-морщинистой скульптурой (с преобладанием поперечных и косых морщин), в задней половине слегка широко продольно вдавленное, вблизи основания с поперечным, более или менее сглаженным вдавлением. Маргинальная и субмаргинальные ячейки, как на рис. 72е. 1-й членик передних лапок с выступом на конце (рис. 72ж). Передние тазики с шипом на конце внутреннего края (рис. 72з). Коготки без зубца (рис. 72и). Пульвиллы маленькие (рис. 72к). Стебелек брюшка относительно короткий. Его длина слегка больше длины 2-го членика, приблизительно в 1,5 раза меньше длины 1-го. Голова, грудь, брюшко, ноги и усики черные, брюшко с синим металлическим оттенком. Мандибулы в вершинной половине буроватые. Ноги отчасти буроватые. Крылья бурые, задние светлее, особенно в задней части. Тегулы черные, сзади бурые. Волоски головы и груди черно-бурые. Длина тела 15-24 мм.
Самец. Наличник широкий, плоский, внизу отогнутый косо вперед, с почти прямым в середине передним краем (рис. 72л). Глаза сильно сближаются книзу (рис. 72м). Ширина лица перед передним глазком почти в 1,5 раза больше минимальной ширины лица на наличнике. Промежуток между задними глазками в среднем в 1,25 раза уже промежутка между одним из них и ближайшим глазом. Лицо с густым серебристым прилегающим опушением и длинными отстоящими серовато-белыми волосками. Переднеспинка с довольно густыми мелкими точками; среднеспинка с густыми (в средней части редкими) мелкими точками, по бокам с нежными сглаженными поперечными морщинами, иногда слабо заметными. Щитик с нежными продольными морщинами, в середине слегка уплощенно-вдавленный. Заднещитик сильно выпуклый, с неправильной морщинистой скульптурой. Срединное поле промежуточного сегмента с нежной сетчатой микроскульптурой, голое, в середине широко продольно вдавленное, с нежной поперечно-морщинистой скульптурой, по бокам с косыми морщинами. Бока среднегруди внизу густо ячееобразно пунктированные, с тенденцией к образованию морщин, с длинными отстоящими волосками и очень короткими нежными полуприлегающими волосками. Форма маргинальной и субмаргинапльных ячеек как на рис. 72н. Стебелек брюшка значительно короче 1-го членика задних лапок (примерно в 1,3 раза), но длиннее 2-го (примерно в 1,4 раза). Голова, грудь и усики черные. Жгутик усиков отчасти бурый. Мандибулы с желто-рыжей предвершинной частью и бурой вершиной. Тегулы отчасти бурые. Голени, передние лапки, средние лапки у основания, вершина передних бедер и средние бедра снаружи рыжие. Стебелек брюшка черный. Брюшко на большей части красно-рыжее. 1-й тергум с черной полосой сверху. 5-й тергум сверху, 6-й и 7-й черные. Длина тела 13-19 мм.
Ареал: Юг Восточной Европы, Северная Африка, Кавказ, ЮгоЗападная Азия, Средняя Азия, Казахстан, Китай, Монголия.
Распространение в Казахстане: Пуст. Кызылкум, присырдарьинские пустыни – Байгакум (близ Чиили), пески Большие Барсуки, п-ов Мангышлак, Рын-пески, Прибалхашье – 5 км юго-зап. Балхаша, 20 км сев. Мын-Арала.
Экология и биология: Пустынный весенний вид. Встречается в песчаных, глинисто-песчаных и солончаковых пустынях.. Самки делают одноячейковые гнезда в знмле и помещают в них гусениц ночниц (Noctuidae).
Синоним: micipsa Morice, 1900 (Psammophila)
Описание: Самка. Наличник равномерно выпуклый, в середине слегка уплощенно-вдавленный, с узкой, слегка расширенной по бокам каймой, с дугообразным (в середине почти прямым) передним краем (рис. 73а). Пунктировка наличника мелкая и редкая: микропунктировка тоже редкая; сетчатая микроскульптура отчетливая. Отстоящие волоски редкие. Прилегающее опушение отсутствует. Внутренние края глаз почти параллельные (рис. 73б), слегка сближаются кверху. Промежуток между задними глазками примерно в 1,3 раза уже промежутка между одним из них и ближайшим глазом. Верх переднеспинки в середине с маленьким продольным вдавлением, гладкий и блестящий. Среднеспинка с редкими мелкими точками, гладкая и блестящая, впереди в середине с 2 сближающимися кзади углубленными бороздками. Щитик сзади с нежными продольными морщинами, впереди сглаженный, с редкими мелкими точками, в середине продольно вдавленный. Задгнещитик сильно выпуклый, с неправильной нежной морщинистой скульптурой, иногда более или менее сглаженный. Срединное поле промежуточного сегмента полуматовое, с нежными поперечными и косыми морщинками, более резко выраженными по бокам, с сетчатой неправильной микроскульптурой, голое. Бока среднегруди с морщинистой скульптурой и редкими, нечетко ограниченными мелкими точками. Маргинальная и субмаргинальные ячейки как на рис. 73в. Передние тазики на вершине внутренней стороны с коротким шипом (рис. 73г). 1-й членик передних лапок с 8 уплощенными крепкими щетинками (рис. 73д). 1-4-й членики асимметричные, с лопастевидным выступом на конце. Коготки с маленьким зубчиком у основания (рис. 73е). Пульвиллы маленькие. Голова, грудь, усики, ноги и брюшко черные, без синего металлического блеска. Коготки буровато-рыжие. Волоски тела черные. Жгутик усиков, лапки и мандибулы буроватые. Крылья (особенно передние) с бурым затемнением. Жилки в основном бурые. Стебелек брюшка примерно в 1,4 раза короче 1-го членика задних лапок и почти в 1,5 раза длиннее 2-го. Длина тела 16-20 мм.
Самец. Передний край наличника слегка дугообразно выемчатый в середине. (рис. 73ж). Глаза сильно сближаются книзу (рис. 73з). Ширина лица на наличнике почти в 2 раза уже ширины лица перед передним глазком. Промежуток между задними глазками в 1,2-1,3 раза уже, чем между одним из них и ближайшим глазом. Нижняя часть лба и наличник с густым прилегающим серебристо-белым опушением и длинными отстоящими черными волосками. Среднеспинка с густыми мелкими, про- дольно удлиненными точками; промежутки между ними имеют тенденцию к образованию морщин. Сетчатая микроскульптура отчетливая. Щитик с продольно-морщинистой скульптурой, в середине слегка продольно вдавленный. Заднещитик с неправильной морщинистой скульптурой. Скульптура срединного поля промежуточного сегмента, как у самки, но несколько резче. Передние тазики с коротким шипом на вершине переднего края. Коготки с очень маленьким зубчиком у основания. Стебелек брюшка немного длиннее 1-го членика задних лапок, более чем в 2 раза длиннее 2-го. Ноги с очень коротким светло-серым прилегающим опушением. 1-3-й тергумы брюшка на большей части, середина 4-го и 5-го тергумов с густым коротким прилегающим серебристо-серым опушением. Окраска тела почти как у самки. Отстоящие волоски головы, груди и ног черно-бурые. Крылья слабее затемненные, чем у самки, особенно задние. Жилки бурые и коричневые. Длина тела 14-18 мм.
Ареал: Юг Восточной Европы, Средиземноморье, Северная Африка, Юго-Западная Азия, Средняя Азия, Казахстан.
Распространение в Казахстане: Пуст. Кызылкум – 50 км зап. Баиркума; Присырдарьинская равнина – Байгакум (близ Чиили); предгорья и низкогорья хр. Каратау – Ачисай, 20 км южн. Чулак-Кургана, 30 км сев.вост. Сузака, 50 км зап. Джамбула; пус. Мойинкум – Акыр-Тобе; низовья р. Чу; вост. Приаралье; пуст. Сексеулдала; пески Большие Барсуки; п-ов Мангышлак; Илийская долина – Или (бывш.), Баканас, 50 км сев.зап. Баканаса, 10 км сев.-вост. и 10 км южн. Капчагая, 10 км сев.-вост. Аяк-Калкана; Вост. Прибалхашье – Карачаган, 35 км юго-зап. пос. Лепсы; Сев. Прибалхашье – окр. Балхаша; Казахский мелкосопочник – Сары-Арка, 60 км вост. Джезказгана, 100 км сев.-вост. Балхаша.
Экология и биология: Пустынный вид. Обычен в песчаных и глинистых пустынях. Активность имаго приходится главным образом на конец весны и начало лета. Самки делают одноячейковые гнезда в земле. Постройка гнезда обычно предшествует охоте. Добыча – гусеницы подгрызающих совок (Noctuidae). Развивается одно поколение в году.
Синонимы: abeillei Marquet, 1881 (Ammophila); polita Mocsary, 1883 (Psammophila); morawitzi Ed.Andre, 1886 (Ammophila).
Описание: Самка. Наличник равномерно выпуклый, с узкой, расширенной по бокам каймой, с широко дугообразно изогнутым передним краем (рис. 73и), с мелкой редкой пунктировкой, более густой вверху. Верх наличник а и низ лба с коротким, слабо развитым, прилегающим опушением и отстоящими волосками разной длины (наиболее длинные в 2,5 раза длиннее диаметра переднего глазка, наиболее короткие – короче его). Глаза почти параллельные (рис. 73и). Промежуток между задними глазками почти в 1,4 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка гладкая и блестящая, с редкой мелкой пунктировкой, без срединного вдавления. Среднеспинка с редкими мелкими точками, редкой микропунктировкой, гладкая и блестящая. Щитик сзади и по бокам с продольными морщинами, на большей части гладкий и блестящий, с отдельными мелкими точками, в середине широко уплощенный. Заднещитик в середине гладкий и блестящий, с крупными точками, по бокам шагренированный, с очень нежными поперечными морщинами. Срединное поле промежуточного сегмента неправильно, довольно грубо, отчасти ячеисто морщинистое (с преобладанием поперечных и косых морщин), с отстоящими светло-серыми волосками. Бока среднегруди с крупной густой пунктировкой и более или менее сглаженными косыми неправильными морщинами. Передние тазики без шипа на вершине (рис. 73к). Коготки без зубца. Пульвиллы хорошо развитые. 1-й членик передних лапок с 6-7 крепкими уплощенными щетинками (рис. 73л). Стебелек брюшка по длине примерно равен 2-му членику задних лапок, значительно короче 1-го членика. Голова, грудь, усики, ноги, стебелек и вершина брюшка черные. Ноги, жгутик, скапус, тегулы, мандибулы, передний край наличника обычно отчасти бурые. Волоски гололвы и груди светло-серые, на наличнике и висках с примесью бурых. 1-й сегмент брюшка (кроме стебелька), 2-й, 3-й и базальная часть 4го красновато-рыжие. Иногда брюшко почти целиком красно-рыжее. Длина тела 13-16 мм.
Самец. Передний край наличника слегка выемчатый или почти прямой (рис. 73м). Наличник слабо выпуклый, впереди слегка отогнут кпереди. Наличник и лоб на большей части с густым прилегающим серебристым опушением. Глаза сильно сближаются книзу (рис. 73м). Промежуток между задними глазками почти в 1,2 раза уже промежутка между одним из них и ближайшим глазом. Верх переднеспинки с редкой мелкой пунктировкой и густым прилегающим серебристо-белым опушением. Среднеспинка с густой (по крайней мере впереди) мелкой пунктировкой и прилегающим, не очень густым серебристо-белым опушением. Отстоящие волоски среднеспинки по длине примерно равны диаметру переднего глазка или немного длиннее и короче. Бока среднегруди с не очень густой пунктировкой, вверху с нежными морщинами, с прилегающим опушением. Щитик на большей части сглаженный, сзади с продольными морщинами. Заднещитик сильно поперечно-бугровидно выпуклый, с отдельными точками, на большей части сглаженный. Срединное поле промежуточного сегмента с неправильной ячеистоморщинистой скульптурой, с длинными отстоящими волосками. Метаплевры и бока промежуточного сегмента с резко выраженными морщинами. Передние тазики без острого шипа. Стебелек брюшка почти в 1,2 раза длиннее 1-го членика задних лапок. Голова, грудь, ноги, усики, стебелек и вершина брюшка черные. Брюшко от вершины стебелька до середины 4-го тергума и основания 5-го стернума красновато-рыжее. Мандибулы в середине светло-рыжие. Ноги и усики отчасти буроватые. Волоски головы и груди светло-серые и белые, только на лбу и висках с примесью бурых. Ноги и брюшко с очень коротким прилегающим инеевидным опушением. Крылья в вершинной части слегка буроватые. Жилки светло-коричневые, слегка рыжеватые. Длина тела 12-15 мм.
Ареал: Южная и Средняя Европа, Северная Африка, Юго-
Западная Азия, Средняя Азия, Казахстан.
Распространение в Казахстане: Сев. Приаралье – Аральск, 30 км юго-вост. Аральска; пуст. Кызылкум; Присырдарьинская равнина; предгорья Заилийского Алатау – Алматы, хр. Богуты; Илийская долина – низовья р. Чарын, 10-20 км сев.-вост. Капчагая, окр. Капчагая; сев. предгорья Джунгарского Алатау – 25 км сев. пос. Чингильды, 20 км сев.-зап. Капчагая; Южн. Прибалхашье – 17 км сев.-зап. Баканаса; Сев. Прибалхашье – 20 км зап. Балхаша, окр. Балхаша; южн. предгорья хр. Тарбагатай – Маканчи; Зайсанская котловина – 45 км южн. Каратогая, 20 км вост. Каратала; Казахский мелкосопочник – 170 км сев. Балхаша, 10 км зап. пос. Ашутасты; Тургайская ложбина – Наурзум, 10 км юго-зап. пос. Шийли (р. Караторгай); пески Большие и Малые Барсуки; Южн. Предуралье – Кандыгач (Октябрьск), Уил, Саралжин, Бабатай; долина р. Урал – 50 км сев. Атырау (Гурьева); Рын-пески.
Экология и биология: Пустынно-степной вид, предпочитает биотопы с глинисто-песчаными почвами; голых песков, по-видимому, избегает. Биология не изучена. Самки, по всей вероятности, делают в земле одноячейковые гнезда и охотятся на гусениц бабочек. Отмечено питание взрослых ос на клоповнике, крестовнике, качиме, доннике, солянках, мари, гораниновии, мордовнике, тамариске, кермеке и подмареннике.
Синонимы: arenaria Fabricius, 1787 (Sphex), nec Linnaeus, 1758; arenosa Gmelin, 1790 (Sphex); argentea N.Kirby, 1798 (Ammophila); macrogaster Dahlbom, 1831 (Sphex); viatica auct., nec Linnaeus, 1758; ebenina Lepeletier, 1845 (Ammophila), nec Spinola, 1838.
Подвиды: mervensis (Radoszkoeski), 1887 (Ammophila) – о-ва Средиземного моря, Средняя Азия, Казахстан; nepalensis (Zavattari), 1909 (Ammophila) – Непал.
Описание: Самка. Наличник сильно выпуклый, со слегка изогнутым между боковыми зубцевидными углами передним краем (рис. 74а), с очень узкой гладкой каймой впереди, с довольно густой крупной пунктировкой. Промежутки между токами блестящие, с микропунктировкой, с очень неправильной микроскульптурой (особенно вверху). Внутренние края глаз почти прямые, параллельные или слегка сближающиеся книзу (рис. 74б). Промежуток между задними глазками примерно в 1,5 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка с густой (вверху редкой) пунктировкой. Промежутки между точками гладкие и блестящие. Среднеспинка с густыми, слегка продольно удлиненными точками, в средней части более редкими, чем по бокам, с гладкими блестящими промежутками. Щитик на большей части гладкий и блестящий, с редкими точками, в задней части с нежными продольными морщинами. Передние тазики без зубца (рис. 74в). 1-й членик передних лапок как на рис. 74г. Заднещитик в середине сзади килевидно приподнятый. Срединное поле промежуточного сегмента с неправильной ячеисто-морщинистой шероховатой скульптурой, с длинными отстоящими черными волосками. Бока среднегруди грубо ячееобразно пунктированные. Стебелек брюшка слегка длиннее 2-го членика задних лапок. 1-й членик задних лапок примерно в 1,5 раза длиннее стебелька. Коготки без зубцов. Пульвиллы не развитые (рис. 74д). Форма маргинальной и субмаргинальных ячеек как на рис. 74е. Голова, грудь, ноги и усики черные. Стебелек и вершинная часть брюшка черные. 1-й и 2-й тергумы, вершина 1-го стернума и 2-й стернум красно-рыжие, отчасти буроватые. Волоски головы и груди черные. Крылья буроватые, задние слабее. Жилки крыльев бурые и коричневые. Длина тела 16-22 мм.
Самец. Наличник со слегка выемчатым в середине передним краем (рис. 74е,ж), с очень мелкой редкой пунктировкой и сетчатой микроскульптурой, прилегающим серым опушением и длинными отстоящими черно-бурыми волосками. Лоб вверху без прилегающего опушения, с густой пунктировкой и неправильной шероховатой микроскульптурой. Глаза сильно сближаются книзу (рис. 74ж). Ширина лица перед передним глазком примерно в 1,5 раза больше минимальной ширины лица на наличнике. Переднеспинка с довольно густой пунктировкой (в середине вверху изреженной), с нежными сглаженными морщинами по бокам и впереди. Среднеспинка с густой пунктировкой, промежутки имеют тенденцию к образованию нежных сглаженных неправильных морщин, особенно по бокам. Скульптура и опушение груди почти как у самки. Срединное поле промежуточного сегмента с длинными волосками, ячеисто-морщинистое. Стебелек брюшка немного короче 1-го членика задних лапок, длиннее 2-го членика примерно в 1,8 раза. Передние тазики без шипа сзади. Голова, усики, грудь, ноги, стебелек, апикальная часть 3-го сегмента и 4-7-й сегменты черные. 1-3-й сегменты целиком или на большей части красно-рыжие. Тегулы сзади бурые. Крылья буроватые, особенно передние. Жилки бурые и коричневые. Коготки на большей части красно-рыжие, без зубчика. Волоски головы и груди черные, на груди часто с примесью светло-серых. Длина тела 12-21 мм.
В Казахстане вид представлен 2 подвидами. Описание типичного подвида приведено выше. Второй подвид – P. h. mervensis (Rad.) – отличается целиком черной окраской брюшка у самки. Найден в Южном Казахстане (хр. Каратау).
Ареал: Почти вся Европа, Северная Африка, Юго-Западная Азия, Средняя Азия, Казахстан, Китай, Монголия и Западная Сибирь.
Распространение в Казахстане: Предгорья и низкогорья хребтов Западного и Северного Тянь-Шаня – почти повсюду; долина р. Сырдарьи и прилегающие пустыни, долина р. Или и прилегающие пустыни; Казахский мелкосопочник – почти повсюду; предгорья и низкогорья Казахстанского Алтая, Саура и Тарбагатая; Прииртышье, Тургайская ложбина, Южное Предуралье.
Экология и биология: Эврибионтный вид. Встречается от пустынь до лесной зоны, а в горах – от сухих предгорий до альпийских лугов. Предпочитает открытые биотопы с изреженной растительностью. В году 2-3 поколения. Зимуют, вероятно, не только личинки, но и взрослые осы, т.к. имеются позднеосенние и ранневесенние находки имаго. Самки делают в земле (обычно в плотном грунте) одноячейковые гнезда. Добыча – гусеницы совок (Noctuidae). Отмечено питание взрослых ос на цветках молочая, зизифоры, примулы, клоповника, селитрянки, ферулы, кермека, качима, мирикарии, тысячелистника, жузгуна, дягиля и других растений.
Синоним: arenaria Luderwaldt, 1897 (Psammophila), nec Fabricius, 1787.
Описание: Самка. Наличник равномерно выпуклый. Передний край слегка дугообразно выступающий, иногда слегка выемчатый (рис. 74з). Пунктировка наличника густая, довольно крупная. Промежутки между точками с сетчатой микроскульптурой. Глаза слегка сближаются книзу, с почти прямым внутренним краем (рис. 74и). Лоб внизу с редкой пунктировкой, широко-клиновидно вдавленный вверху. Промежуток между задними глазками приблизительно в 1,7 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка в середине слегка продольно вдавленная. Среднеспинка с густой, в середине изреженной пунктировкой и редкой микропунктировкой, с длинными отстоящими волосками. Щитик в середине слегка уплощенно-вдавленный, гладкий и блестящий, с редкой пунктировкой, сзади с короткими продольными морщинами. Заднещитик сильно выпуклый, в середине гладкий и блестящий, с очень мелкими точками. Бока среднегруди морщинистые и пунктированные, без прилегающего опушения. Срединное поле промежуточного сегмента с морщинисто-пунктированной скульптурой, с длинными отстоящими волосками, с нечетко ограниченными точками. Базальные членики передних лапок как на рис. 74к. Асимметрия 14-го члеников сильная. Коготки без зубца. Пульвиллы нормально развитые (рис. 74л). форма маргинальной и субмаргинальных ячеек как на рис. 74м. Стебелек брюшка значительно (примерно в 1,25 раза) короче 1-го членика задних лапок, но длиннее 2-го более чем в 1,3 раза. Голова, грудь, усики, ноги черные. Мандибулы отчасти буровато-красные. Стебелек брюшка черный. 1-й тергум, вершина 1-го стернума, 2-й, 3-й сегменты и большая часть 4-го рыжевато-красные, отчасти буроватые. Голова с черными, грудь с черными, бурыми и светло-серыми волосками. Крылья с относительно слабым коричневым или желтовато-коричневым затемнением. Жилки коричневые и рыжевато-коричневые; коста и субкоста буроватые. Тегулы на заднем крае буроватые. Длина тела 16-21 мм.
Самец. Наличник широкий, с небольшой выемкой в середине переднего края (рис. 74н). Лоб на большей части и наличник с густым прилегающим серебристым опушением и длинными отстоящими светлыми волосками. (Несколько волосков бурые.) Глаза сильно сближаются книзу (рис. 74н). Лицо перед передним глазком примерно в 1,6-1,7 раза шире, чем на наличнике. Промежуток между задними глазками в 1,4-1,5 раза шире промежутка между одним из них и ближайшим глазом. Переднеспинка и среднеспинка с мелкой пунктировкой и длинными светло-серыми волосками, гладкие и блестящие. Промежутки между точками шире, равны или уже точек. Пунктировка в середине среднеспинки (по бокам от срединной борозды) и сзади более редкая, чем по бокам. Скульптура щитика, заднещитика, боков среднегруди и срединного поля промежуточного сегмента почти как у самки. Пульвиллы хорошо развитые. Стебелек брюшка примерно в 1,2 раза длиннее 1-го членика задних лапок или 1-го тергума, более чем в 2 раза длиннее 2-го членика. Окраска тела почти как у самки. Ноги отчасти буроватые. Коготки буроватокрасные. 1-й сегмент брюшка (кроме стебелька), 2-4-й сегменты, основание 5-го стернума и передние участки боков 5-го тергума краснорыжие. Крылья со светло-коричневым затемнением. Жилки коричневые, отчасти буроватые. Длина тела 13-20 мм.
Ареал: Почти вся Европа, Средняя Азия, Казахстан, Западная Сибирь.
Распространение в Казахстане: Сев. Приаралье – 10 км сев. Аральска; предгорья и низкогорья Западного Тянь-Шаня – 18 км южн. АкарТобе, Бурное, 30-40 км южн. Ленгера; предгорья и низкогорья Заилийского Алатау – 110 км зап. Алматы, Тургень, окр. Чемолгана, Тескенсу, Куртогай (р. Чарын); хр. Турайгыр, Бартогай (р. Чилик); предгорья Джунгарского Алатау – Сарканд, Басший; Сев. Прибалхашье – 5 км югозап. Балхаша, пуст. Бетпакдала – р. Сарысу; долина р. Или – пос. Или (бывш.), 17 км сев.-зап. Баканаса; Казахский мелкосопочник – 150-170 км сев. Балхаша, 45 и 70 км южн. Караганды, 27 км юго-зап. Целинограда, Кургальджино, 10-40 км юго-зап. Аркалыка, 10 км вост. пос. Амангельды, 15 км сев.-вост. Амантогая, 30-45 км сев.-вост. Державинска, 1020 км южн. Горняка, Рузаевка, 15 км сев.-зап. Целинограда, 60 км вост. Джезказгана, 195 км сев.-вост. Караганды, 20 км зап. Жана-Арки, Коксенгир, 80-100 км сев.-вост. Балхаша, 8 км юго-зап. Каркаралинска, 40 км сев.-вост. Кайнара, 50 км сев. Актогая, 40 км зап., 61 км сев. и 20 км южн. Караула, 12 км сев. Маинкаина, 13 км сев.-вост. Ульяновского, 5070 км юго-юго-зап. Баян-Аула; Тургайская ложбина – Аман-Карагай, Наурзум, Шийли (р. Караторгай), Иргиз; Северо-Казахстанская равнина – 20 км юго-зап. и 100 км зап. Павлодара, 150 км вост. Кустаная; Прииртышье – 9 км сев.-зап., 9 км юго-вост. и 5 км южн. пос. Долонь, Щербакты, 23 км сев.-зап. Семеновки; предгорья и низкогорья Казахстанского Алтая – 15 км сев. Самарского, 10 км юго-вост. пос. Алтай, Самaрское, 15 км сев. Алексеевки (хр. Азутау), Катон-Карагай; Зайсанская котловина – 15 км южн. Аксуата, Акжар, 7 км юго-зап. Акжара, 40 км зап. Черняевки, 5-15 км и 20 км вост. Каратала, 45 км южн. Каратогая, Рожково; предгорья хр. Тарбаготай – Маканчи, 20 км вост. Маканчи; предгорья хр. Манрак – 30 км зап. пос. Чиликты (р. Кандысу); Южное Предуралье – 5-10 км сев.-вост. Актюбинска, Саралжин, пески Большие Барсуки – Челкар; долина р. Урал – Гурьев, 10-50 км сев. Гурьева, Гребенщиков, Индерборский, Володарское.
Экология и биология: Степной вид, на юге встречается в горах и по долинам рек. Самки делают одноячейковые гнезда в земле и охотятся на гусениц совок (Noctuidae).
Вид известен автору только по работам К.Цунеки (Tsuneki, 1971) и Ф.Ф.Коля (Kohl, 1901), на основании которых приводится ниже его описание.
Описание: Самка. Наличник слабо выпуклый. Передний край его в середине прямой. Ширина лица перед передним глазком немного меньше, чем на наличнике. Ширина промежутка между одним из задних глазков и ближайшим глазом равна длине 3-го членика жгутика и на диаметр глазка шире промежутка между задними глазками. Воротничок переднеспинки по бокам закругленный, в середине сильно возвышающийся, с крутыми передней и боковыми сторонами. Среднеспинка гладкая и блестящая, с редкой пунктировкой, в середине задней половины без пунктировки. Точки разные по величине: очень грубые или умеренно крупные. Щитик выпуклый, пунктированный, без морщин. Бока среднегруди гладкие и умеренно густо, но очень грубо пунктированные, с гладкими промежутками между точками. Мезоплевры морщинистые. Срединное поле промежуточного сегмента поперечно морщинистое. Бока его косо морщинистые. Стебелек брюшка прямой, его длина примерно равна 2/3 длины 1-го членика задних лапок. Передние тазики без зубца. Пульвиллы маленькие, коготки без зубца. 1-й членик передних лапок с 8-9 крепкими длинными щетинками. Маргинальная (радиальная) ячейка передних крыльев на вершине закругленная. Голова, грудь, усики и ноги черные. Брюшко от стебелька красное. Вершинный сегмент отчасти коричневатый. Крылья слегка буроватые. Волоски тела беловатые и очень редкие. Длина тела 17 мм.
Самец. Передний край наличника в середине почти прямой. Глаза сильно сближаются книзу. Промежуток между задними глазками в 1,2 раза уже промежутка между одним из них и ближайшим глазом. Минимальная ширина лица на наличнике приблизительно равна длине 3-го и 4-го члеников усика вместе. Виски относительно узкие, примерно вдвое уже глаз (см. в профиль). 1-й и 2-й членики усиков, вместе взятые, короче, чем 3-й и 4-й. 3-й членик примерно в 3,5 раза длиннее ширины на конце. Голова с редкой пунктировкой и нежной микропунктировкой. Воротничок переднеспинки сбоку закругленный. Эпистернальный шов отчетливый. Мезоплевры сверху густо и грубо пунктированные, с длинными густыми волосками. Пунктировка переднеспинки, среднеспинки и щитика редкая и довольно грубая, несколько гуще, чем у самки; промежутки между точками сильно блестящие. Срединное поле промежуточного сегмента густо и грубо пунктированное, с тенденцией к образованию поперечных морщин. Метаплевры и бока промежуточного сегмента с продольными и поперечными морщинами. Коготки без зубца. Пульвиллы очень маленькие. Тазики, бедра и голени с густым коротким инеевидным опушением. Маргинальная и субмаргинальные ячейки как на рис. 75г. Стебелек брюшка в 1,2 раза длиннее 1-го членика задних лапок, который почти равен по длине 1-му тергуму. Голова, грудь, усики и ноги черные. Стебелек брюшка черный. Брюшко с красным очнованием, от вершины 3-го тергума черное. Волоски груди беловатые. Длина тела 14 мм.
Ареал: Точно не выяснен. Вид известен из Казахстана и Монголии.
Распространение в Казахстане: Зайсанская котловина – Зайсан.
Экология и биология: Не известны. Самки, вероятно, делают одноячейковые гнезда в земле и охотятся на гусениц совок или других бабочек.
Синонимы: madeirae Dahlbom, 1843 (Psammophila); klugii Lepeletier, 1845 (Ammophila); psammodes Lepeletier, 1845 (Ammophila); capuccina A.Costa, 1858 (Ammophila); ? spinipes F.Smith, 1878 (Ammophila); lanuginosa Marquet, 1881 (Ammophila); ? laeta Bingham, 1897 (Ammophila); ? durga Nurse, 1903 (Ammophila); errabunda Mercet, 1906 (Ammophila), nec Kohl, 1901; homogenea Mercet, 1906 (Ammophila).
Подвиды: senilis (Dahlbom), 1843 (Psammophila) – ЮжнаяЕвропа, ? Азия; argentata (Lepeletier), 1845 (Ammophila) – Сев. Африка; caspiciosa (F.Smith), 1856 (Ammophila) – Австралия; apakensis (Tsuneki), 1971 (Ammophila) – Сев. Китай.
Описание: Самка. Наличник сильно поперечно-выпуклый, с почти прямым передним краем между зубцевидными боковыми углами (рис. 75а), с редкой мелкой пунктировкой, с густым прилегающим серебристо-серым опушением в верхней половине и длинными светло-серыми волосками (с небольшой примесью черных). Лицо широкое; глаза почти параллельные (рис. 75а)ю Промежуток между задними глазками в 1,51,6 раза уже промежутка между одним из них и ближайшим глазом. Среднеспинка с густой крупной пунктировкой. Точки в 3-4 раза уже переднего глазка; промежутки между точками блестящие, с микропунктировкой и очень нечеткой сетчатой микроскульптурой. Грудь со светлосерыми длинными волосками и с небольшой примесью черно-бурых внизу. Щитик сзади с продольными морщинами. Срединное поле промежуточного сегмента с длинными отстоящими волосками, с неправильной ячеисто-морщинистой скульптурой. Форма маргинальной и 2-3й субмаргинальных ячеек как на рис. 75г. Бока среднегруди с прилегающим серебристым опушением. 1-3-й членики передних лапок асимметричные, с лопастевидным выступом на конце. 1-й членик с 3 крепкими длинными щетинками на вершине выступа и 4 длинными крепкими щетинками на наружном крае (рис. 75б). Ноги с очень коротким инееобразным прилегающим опушением. Коготки без зубцов. Пульвиллы развитые (рис. 75в). Голова, грудь, ноги и усики черные. Мандибулы красно-бурые на большей части. Стебелек брюшка почти целиком черный. 1-й тергум, вершина 1-го стернума, 2-й, 3-й сегменты и 4-й на большей части (иногда целиком) рыжевато-красные, иногда с бурыми пятнами. Изредка 5-й сегмент отчасти тоже рыжевато-красный. Верши- на брюшка черная, без металлического синего блеска. Коготки отчасти красно-рыжие. Длина тела 16-21 мм.
Самец. Наличник широкий (рис. 75д), почти равномерно выпуклый, с почти прямым в середине передним краем, с хорошо выраженными тупыми боковыми углами на переднем крае (рис. 75д), с густым прилегающим серебристым опушением и отстоящими белыми волосками. Верх головы с примесьюбурых волосков. Глаза сближаются книзу, но менее, чем у многих других видов (рис. 75д). Промежуток между задними глазками примерно в 1,25 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка и среднеспинка с густой крупной пунктировкой (точки в 3,5-4 раза уже переднего глазка), помимо длинных отстоящих светлых волосков с короткими прилегающими волосками. Бока среднегруди и промежуточного сегмента с густым прилегающим серебристым опушением, отчасти скрывающим скульптуру. Щитик более или менее сглаженный, со слабым срединным продольным вдавлением. Срединное поле неправильно ячеисто-морщинистое, с длинными отстоящими волосками. Ноги с коротким инееобразным прилегающим опушением. Тазики опушенные, как бока среднегруди. Стебелек брюшка примерно в 1,3 раза длиннее 1-го членика задних лапок и в 2,3 раза длиннее 2-го. Голова, грудь, усики, ноги, стебелек брюшка (кроме вершины) и вершина брюшка черные. 1-й сегмент (кроме большей части стебелька), 2-й, 3-й и большая часть 4-го сегмента красно-рыжие. Крылья коричневатые, жилки рыжевато-коричневые и буровато-коричневые.
Лапки буроватые. Коготки буровато-красные. Длина тела 12-20 мм.
Ареал: Южная и Средняя Европа, почти вся Африка, ЮгоЗападная Азия, Средняя Азия, Казахстан, Монголия, Зап. Китай, Австралия.
Распространение в Казахстане: Плато Мангышлак – Таучик; Присырдарьинские пустыни – Баламурун (близ Чиили), Коксу; пескиМойинкум – Акыр-Тобе, 30 км южн. Чу; о. Барсакеоьмес; пуст. Кызылкум -– 5-8 км юго-зап. Чардары; пуст. Сексеулдала – 60-70 км сев.-зап. Фурмановки; долина р. Или и Южное Прибалхашье – Баканас, 50-65 км зап. и 17 км сев.-зап. Баканаса, Айдарлы, Капчагай, 35 км сев.-зап. Капчагая,
Или (бывш.), 30-35 км сев. Чилика (Аяк-Калкан); предгорья и низкогорья Заилийского Алатау – Бартогай, горы Богуты, р. Чарын, 10 км вост. Алматы, 10 км сев.-зап. Чемолгана; предгорья и низкогорья Джунгарского Алатау – окр. Жаркента ( бывш. Панфилов), Басший, Коныролен, Егеньсу; Сев. и Зап. Прибалхашье – 5-15 км юго-зап. Балхаша, 5-10 км южн. Мынарала; Зайсанская котловина – 20 км вост. Каратала; Прииртышье – Щербакты, Семипалатинск; Казахский мелкосопочник - Балкашино, Шортанды; Тургайская ложбина – Наурзум; горы Мугоджары – Кандагач; Южн. Предуралье – Бабатай; долина р. Урал – Гребенщиков, Елтай, 50 км сев. Атырау (бывш. Гурьев), окр. Атырау; Прикаспийская низменность – Ащисай (близ пос. Кульсары).
Экология и биология: Пустынный вид. Предпочитает биотопы с рыхлой песчаной почвой. В степной зоне встречается на песках с изреженной растительностью. Самки гнездятся в уплотненном песчаном грунте. Постройка гнезда предшествует охоте. Добыча – гусеницы совок (Noctuidae). Отмечено питание взрослых ос на цветках кермека, клоповника, морковника, качима, тамариска, селитрянки, ак-курая, молочая, саусуреи, лука, жузгуна, подмаренника и чингиля.
Типовой вид – Eremochares doriae Gribodo, 1883.
Диагностические признаки: Внутренние края глаз прямые или слегка вогнутые, сильно сближаются книзу. Лобная линия отсутствует. Усиковые ямки отделены от наличника промежутком, равным по ширине примерно 2 диаметрам ямок у самки, 3 и более – у самца. Передний край наличника самки дугообразный или с широкой срединной лопастью, ограниченной по бокам вырезками. Верхняя губа шире своей длины, выпуклая у самки. Внутренний край мандибулы самки с большим зубцом (иногда еще с дополнительным маленьким зубцом), у самца без зубца или с маленьким зубцом. Мандибулярная ямка закрытая у самки и открытая у самца. Воротничок переднеспинки умеренной длины, с постепенно спускающейся к голове передней поверхностью. Стебельковая ямка почти целиком огружена тергумом промежуточного сегмента. Мезостернальная область груди с выступом впереди, направленным к передним тазикам. Эпистернальная борозда идет от субалярной ямки вниз мимо скробальной ямки. Маргинальная ячейка на вершине закругленная; 2-я субмаргинальная ячейка варьирует по форме. 2-я возвратная жилка обычно интерстициальная между 2-й и 3-й субмаргинальными ячейками. Передняя лапка самки с гребнем довольно коротких крепких шипов. Средние тазики почти соприкасаются друг с другом. Средние голени с 1 шпорой. Апикально-вентральные щетинки 5-го членика лапок умеренно широкие. Коготки с 2 базальными зубцами. Аролиумы большие. Стебелек брюшка состоит только из очень длинного 1-го стернума, который достигает основания 2-го стернума. Дыхальца 1-го тергума расположены примерно в середине длины членика. Боковые края 8-го стернума самца сближаются к вершине. Вершина его закругленноусеченная. Вершинная часть вальвы пениса с рядом зубцов на вентральном крае. Гоностиль не раздвоен на вершине.
Биология: Самки обычно гнездятся скоплениями. Норки имеют 1 ячейку на конце. Постройка гнезда предшествует охоте. Добыча – личинки саранчовых (Acrididae). В одной ячейке запасается 4-6 экземпляров добычи, причем провиантирование происходит в течение нескольких дней – по мере развития личинки.
Распространение: Представители рода обнаружены в Северной Африке, Южной Европе, Юго-Западной Азии, Средней Азии и Казахстане. Известно 5 видов. В Казахстане 2 вида.
Литература: Казенас, 1970; Смирнов, 1915; Kohl, 1906.
Определительная таблица видов рода Eremochares
1 (2). Темя и среднеспинка с очень густым опушением, скрывающим скульптуру. Минимальное расстояние между глазами равно длине 3-го членика усиков. Грудь ♀ красно-коричневая. ………………….
..…………………………………………………….E. mirabilis (Guss.)
2 (1). Темя и среднеспинка с менее густым опушением, не скрывающим скульптуру. Минимальное расстояние между глазами больше длины 3-го членика усиков. Грудь ♀ обычно черная……..E. dives (Br.)
Синонимы: melanopus Lucas, 1848 (Ammophila); festiva F.Smith, 1856 (Ammophila); elegans F., 1856 (Ammophila); nigritarsa Walker, 1871 (Ammophila); doriae Gribodo, 1882; retowskii Konow, 1887 (Parapsammophila); orichalceomicans Strand, 1915 (Ammophila).
Описание: Самка. Передний край наличника без выемки и боковых зубцов (рис. 75е), в середине с узкой голой каймой. Глаза сильно сближаются книзу (рис. 75е). Ширина лица перед передним глазком примерно в 1,5 раза больше минимальной на наличнике. Промежуток между задними глазками приблизительно в 1,1 раза шире промежутка между одним из них и ближайшим глазом. Лицо с густым прилегающим серебристым (слегка желтоватым) опушением и длинными (равными по длине) желтовато-серыми светлыми волосками. Воротничок переднеспинки сильно выпуклый, примерно в 2 раза шире своей длины, в середине с небольшим продольным углублением. Среднеспинка с густой мелкой пунктировкой и густой микропунктировкой, на большей части с густым прилегающим опушением, в середине более редким. Щитик сильно выпуклый, в середине слегка уплощенно-вдавленный. Срединное поле промежуточного сегмента на большей части изломанно-поперечноморщинистое и густо пунктированное, с густым прилегающим опушением и длинными отстоящими волосками, по бокам с правильными поперечно-косыми негрубыми морщинами, без пунктировки, голое, блестящее. Среднегрудь впереди внизу в середине с выступом. 1-й членик передних лапок с короткими крепкими шипами (они короче ширины членика) на наружной стороне, на конце вытянутый (рис. 75ж). Жилкование переднего крыла как на рис. 75и. Коготки лапок с зубчиком (рис. 75з). Стебелек брюшка (рис. 75к) по длине почти равен 1-му и 2-му членикам задних лапок вместе. Анальная ячейка заднего крыла замкнутая перпендикулярной жилкой. Голова черная. Мандибулы в базальной половине красно-рыжие. Наличник впереди отчасти рыжий. Усики черные, скапус на вершине буроватый. Грудь черная. Плечевые бугры рыжеватобурые. Тегулы буровато-рыжие. Субкоста и радиальная жилка (радиальный сектор) бурые, остальные жилки темно-коричневые, у основания крыльев рыжеватые. Стебелек брюшка черный. 1-й тергум сверху отчасти черный, в вершинной части и по бокам красно-рыжий, более или менее буроватый. 1-й стернум на вершине, 2-й и 3-й сегменты целиком, бока 4-го тергума, 4-й стернум, бока 5-го и 6-го тергумов красно-рыжие или буровато-рыжие. Остальная часть брюшка черная. Вершинная часть крыльев слегка буроватая. Ноги отчасти красно-рыжие. Длина тела 2229 мм.
Самец. Наличник равномерно выпуклый. Передний край наличника почти прямой или слегка выемчатый в середине (рис. 75л). Глаза сильно сближаются книзу (рис. 75м). Ширина лица перед передними глазком примерно в 1,8 раза меньше минимальной (на наличнике). Промежуток между задними глазками в 1,5-1,6 раза шире промежутка между одним из них и ближайшим глазом. Лоб вверху широко равномерно вдавленный. Средняя линия слабо выражена. Переднеспинка примерно в 2 раза длиннее своей ширины. Среднеспинка с густой пунктировкой и густыми светло-серыми волосками. Прилегающее опушение слабее развитое, чем у самки. Среднегрудь впереди снизу с коническим бугром. Стебелек брюшка (1-й стернум) длинный, немного длиннее 1-го и 2-го члеников задних лапок вместе. Тело черное. 1-й тергум по бокам, 2-й тергум и 2-й стернум обычно более или менее буровато-рыжеватожелтые. Брюшко с очень коротким инеевидным опушением. Передние крылья не затемненные. Длина тела 18-25 мм.
Ареал: Южная Европа (Средиземноморье), Северная Африка, Юго-Западная Азия, Средняя Азия, Казахстан.
Распространение в Казахстане: Долина р. Сырдарьи и присырдарьинские пустыни – Байгакум (близ Чиили), Кзыл-Орда; пуст. Кызылкум; о. Возрождения (Аральское море); п-ов Мангышлак – ур. БасКудук; Рын-пески; долина р. Или – пос. Или (бывш.), 8 км вост. пос. Или, 15 км вост. Аяк-Калкана; предгорья Заилийского Алатау (равнина) – 10 км сев.-зап. Чемолгана.
Экология и биология: Обычно вид встречается на участках солончаков с уплотненной песчано-глинистой почвой в долинах рек в пустынной зоне. Самки гнездятся в земле на голых участках; делают одноячейковые гнезда. Добыча – личинки саранчовых (представители родов Dociostaurus, Notostaurus, Oedipoda, Calliptamus, Oedaleus, Chorthippus, Eremippus и др.). Для вида отмечено замедленное, или прогрессивное, провиантирование (Казенас, 1970) в течение всего периода развития личинки, который длится 6-7 дней. Взрослые осы питаются на цветках различных растений (солнцецвета, ластовня, тамариска, кермека, горца и др.).
Описание: Самка. Передний край наличника дугообразный, без зубцов (рис. 76а). Мандибулы как на рис. 76б. Глаза сильно сближаются книзу (рис. 76а). Лицо узкое, вверху перед передним глазком в 2,3 раза шире, чем внизу на наличнике. Промежуток между задними глазками примерно в 1,1 раза шире промежутка между одним из них и ближайшим глазом. Лоб сильно вдавленный. Переднеспинка с продольным вдавлением на вершине, без резкого уступа спереди (рис. 76в,г). Ее длина в середине примерно в 1,4 раза меньше ширины в середине. Щитик и заднещитик сильно бугровидно выпуклые, с продольным вдавлением в середине. Срединное поле промежуточного сегмента по бокам голое, с поперечными нежными морщинами, на большей части неправильно морщинистое, с густым прилегающим опушением. Среднегрудь снизу в середине с коническим бугорком, ограничивающим сзади вогнутую поверхность. Жилкование передне-вершинной части передних крыльев как на рис. 75и. Голова, грудь и тазики с обильным прилегающим опушением, скрывающим скульптуру. 1-й членик передних лапок как на рис. 76д. Ноги и брюшко с очень коротким инееобразным опушением. Стебелек брюшка немного короче задней голени, слегка длиннее (почти равен), чем 1-й и 2-й членики задних лапок вместе. Голова, грудь и брюшко почти целиком рыжие или буровато-рыжие. Мандибулы на большей части желтовато-рыжие. Скапус рыжий, педицеллюс буровато-рыжий; жгутик бурый, снизу рыжеватый. Длина тела 24-28 мм.
Самец. Наличник равномерно выпуклый. Передний край его слегка выемчатый в середине (рис. 76е). Глаза сильно сближаются книзу (рис. 76ж). Лицо узкое (рис. 76ж). Промежуток между задними глазками примерно в 1,5 раза шире промежутка между одним из них и ближайшим глазом. Голова и грудь с отстоящими белыми волосками (как у самки), с густым прилегающим опушением. 1-й членик стебелька слегка длиннее 1-го и 2-го члеников задних лапок. Голова на большей части черная. Наличник и низ лба рыжевато-желтые. Скапус спереди светлокоричневый, рыжеватый. Грудь под прилегающим опушением бурая и черная, снизу отчасти рыжая. Ноги рыжие, голени и лапки более или менее бурые. Бедра сверху более или мене бурые. Стебелек брюшка, 1-й тергум сверху, 3 вершинных сегмента брюшка бурые или черные. Крылья прозрачные, слегка рыжевато-светло-коричневые. Субкоста более или менее бурая. Длина тела 20-26 мм.
Ареал: Иран, Узбекистан, Южный Казахстан.
Распространение в Казахстане: Пуст. Кызылкум (30 км южнее Чардары).
Экология и биология: Пустынный вид, приуроченный к пескам. Биология не изучена; вероятно, она близка к таковой у E. dives.
Типовой вид – Sphex sabulosus Linnaeus, 1758.
Синонимы: Ammophilus Latreille, 1802-1803; Miscus Jurine, 1807; Ammophilus Latreille, 1829; Coleoptera Lepeletier, 1845; Argyrammophila Gussakovskij, 1928; Apycnemia Leclercq, 1961.
Диагностические признаки: Внутренние края глаз прямые или слегка изогнутые, обычно сближающиеся книзу, но иногда почти параллельные у самок. Вершинные членики усиков самцов часто с тилоидами. Усиковые ямки у самок почти соприкасаются с наличником или отделены от него промежутком, почти равным диаметру ямок, у самцов отделены от наличника промежутком, равным от 1 до 2 диаметров ямок. Передний край наличника самки обычно с прямой или изогнутой спереди срединной лопастью, ограниченной по бокам маленьким зубцом или углом и изредка слегка выемчатой в середине. Внутренний край мандибул самки с 1-3 зубцами. Ротовые органы обычно длинные. Манжибулярные ямки закрытые (узко открытые у некоторых самцов). –Воротничок часто удлиненный. Срединное поле промежуточного сегмента выделено особой скульптурой или опушением. Стебельковая ямка обычно окаймлена снизу широко или узко стернумом промежуточного сегмента. Передняя часть мезоплевр снизу иногда вогнутая (для прикладывания передних тазиков). Эпистернальная борозда обычно имеется, не пересекает скробальную ямку. Вершина маргинальной ячейки заостренная или закругленная. Форма 2-й субмаргинальной ячейки варьирует. 3-я субмаргинальная ячейка иногда отсутствует или стебельчатая. 1-я или 2-я возвратные жилки иногда интерстициальные, но обычно впадают во 2-ю субмаргинальную ячейку. Передние лапки самки с гребнем щетинок. Членики передних лапок, особенно 1-й, обычно с выступом на наружном крае. Средние тазики соприкасающиеся или разделенные промежутком. Средние голени обычно с 2 шпорами. Коготки обычно простые, изредка с зубчиком у основания на внутреннем крае. Аролиумы от маленьких до больших. Стебелек брюшка очень длинный, состоит из цилиндрического 1-го стернума и лабо расширяющегося кзади 1-го тергума, как бы двухсегментный. Вершина 1-го стернума не достигает основания 2-го. Дыхальца 1-го тергума расположены позади его середины. 8й стернум самца обычно усеченный на вершине и часто выемчатый. Вальва пениса на вершине с зубцами (на вентральном крае). Гоностиль не раздвоенный на вершине.
Биология: Самки роют норки в земле и устраивают в них одноячейковые гнезда. Постройка гнезда обычно предшествует охоте. Добыча – гусеницы бабочек (Lepidoptera) и личинки пилильщиков (Hymenoptera, Tenthredinoidea). Обычно в каждой ячейке добыча запасается по типу массового провиантирования, но для нескольких видов характерно прогрессивное провиантирование.
Распространение: Известно около 200 видов из всех зоогеографических областей Земного шара. В Казахстане 20 видов.
Литература: Гуссаковский, 1930; Казенас, 1968; Alfieri, 1946; Kohl, 1906; Roth, 1928, 1929; Tsuneki, 1968, 1971b.
1 (2). Бока среднегруди без эпистернального шва, или он развит только вверху, за плечевыми буграми. Коготки у основания с зубчиком. Стигмы 2-го сегмента брюшка расположены в середине длины тергума. ……………………………………………A. hungarica Mocs.
2 (1). Бока среднегруди c хорошо развитым эпистернальным швом (иногда скрытым под опушением). Коготки без зубчика. Стигмы 2-го сегмента брюшка расположены в задней половине тергума.
3 (14). Длина переднеспинки приблизительно равна ее ширине, иногда меньше, но не более чем в 1,5 раза. Верхняя часть переднеспинки обычно с поперечными складками. Вдавленная часть среднегруди (эпикнемиальное поле) обычно ограничена сзади поперечным, глубоко вырезанным в середине килем (у ♂ слабо развитым).
4 (5). Затылок с высоким килем. Передняя часть переднеспинки выступает вперед, скрывая шейку, сильно выемчатая в середине. ………
……………………………………………….A. elongata Fisch.-Waldh.
5 (4). Затылок сзади с низким килем. Переднеспинка не выдвинута вперед, не скрывает шейку и без вырезки.
6 (9). Срединное поле промежуточного сегмента по бокам голое, в середине опушенное.
7 (8). Ноги более или менее красные, отчасти затемненные. Переднеспинка ♀ посредине вдавленная. Темя с бугром…………………. …...…………….………………………………….A. occipitalis F.Mor.
8 (7). Ноги черные. Переднеспинка ♀ почти не вдавленная, примерно в 1,5 раза шире своей длины. Темя ♀ без бугра………………… ………………………………………………………A. sinensis Sickm.
9 (6). Срединное поле промежуточного сегмента полностью опушенное.
10 (11). Наличник ♂ вверху сильно выпуклый, внизу далеко выступающий вперед. Срединное поле промежуточного сегмента неправильно ячеисто-морщинистое, со слабым опушением……………..
………………………………………………………A. longicollis Kohl
11 (10). Наличник ♂ другой формы. Опушение срединного поля промежуточного сегмента сильнее развитое.
12 (13). Переднеспинка и среднеспинка с хорошо развитыми четкими поперечными морщинами. Опушение тела не скрывает скульптуру. Наличник ♀ почти плоский. Переднеспинка впереди почти не вогнутая. Скапус ♂ обычно целиком красный……………………. …………………………………………………..…A. gracillima Tasch.
13 (12). Переднеспинка и среднеспинка, самое большое, с несколькими слабо заметными морщинами, скрытыми под опушением. Опушение тела густое, скрывает скульптуру. Наличник ♀ выпуклый. Переднеспинка впереди слегка вогнутая. Скапус ♂ с темными пятнами.
14 (3). Длина переднеспинки значительно меньше ее ширины, обычно со слабыми поперечными складками или без них. Низ среднегруди без поперечного киля.
15 (16). Наличник ♀ сильно выпуклый, у ♂ с большим бугром, с выемкой на переднем крае. Верхняя поверхность промежуточного сегмента ♀ с очень густым опушением, скрывающим его скульптуру. Волоски передней поверхности головы с золотистым блеском.
……………………………………………………….A. sareptana Kohl
16 (15). Наличник ♀ и ♂ другой формы, обычно уплощенный.
17 (18). Наличник ♀ впереди на значительном протяжении вдавленный и глубоко выемчатый в середине. Передняя поверхность головы с серебристым прилегающим опушением и длинными отстоящими светлыми волосками. Переднеспинка впереди в середине с продольной выемкой, мелкоморщинистая. Наличник ♂ почти плоский, широкий, слегка вдавленный в середине, с треугольной вырезкой впереди. Членики лапок относительно широкие, отделены друг от друга перетяжками. ……………………...….A. adelpha Kohl
18 (17). Наличник ♀ не вдавленный и без глубокой выемки в середине.
Наличник ♂ обычно другой формы.
19 (32). Срединное поле промежуточного сегмента голое или с волосками только в средней части.
20 (21). Среднеспинка с очень широкой продольной бороздой в середине. Переднеспинка удлиненная, но ширина ее все же больше длины. …..……………………………………………….….A. induta Kohl
21 (20). Среднеспинка с узкой продольной бороздой или без нее.
22 (25). Среднеспинка с редкими поперечными и слабыми продольными складками. 3-я кубитальная ячейка переднего крыла не стебельчатая. Последний сегмент брюшка с голубым блеском………….. ……………………………………………………..….A. terminata Sm.
23 (24). Бока заднегруди и промежуточного сегмента с густым белым опушением. ………………………………..A. terminata terminata Sm.
24 (23). Бока заднегруди и обычно промежуточного сегмента без густого белого опушения. ………………………..A. terminata terminata Sm.
25 (22). Среднеспинка без складок или с очень слабо развитыми складками. 3-я кубитальная ячейка переднего крыла стебельчатая. Последний сегмент брюшка без голубого блеска.
26 (31). Ноги черные.
27 (28). Отстоящие волоски головы только черные. Срединное поле промежуточного сегмента между косыми складками матовое, с густой микроскульптурой (шагренированное), в задней части почти без складок. Дистальный зубец мандибул ♀ отделен от 2 проксимальных узкой вырезкой. 1-й и 2-й тергумы брюшка ♂ сверху сплошь черные. ……………………………A. pubescens Curt.
28 (27). Отстоящие волоски головы почти все светлые. Срединное поле промежуточного сегмента между косыми складками гладкое и блестящее. Дистальный зубец мандибул ♀ отделен от 2 проксимальных широкой вырезкой. 1-й и 2-й тергумы брюшка ♂ краснорыжие, только 1-й у основания сверху черный. …A. campestris Jur.
29 (30). Весь стебелек брюшка красный. 1-й тергум брюшка ♂ целиком красный. Бока среднегруди с большим каплевидным пятном густого прилегающего опушения. ……..A. campestris separanda F.Mor.
30 (29). По меньшей мере 1-й стернум брюшка ♀ черный. 1-й тергум брюшка ♂ целиком или у основания черный. Бока среднегруди с
менее выраженным пятном густого прилегающего опушения……. ..…………………………………………A. campestris campestris Latr.
31 (26). Ноги отчасти красные. Скульптура груди несколько нежнее, чем у A. campestris Jur. ………………………………....A. cellularis Guss.
32 (19). Срединное поле промежуточного сегмента целиком покрытое волосками.
33 (42). Среднегрудь впереди без бугровидного выступа.
34 (41). Промежуточный сегмент с опушением, не скрывающим скульптуру.
35 (38). Ноги целиком или отчасти красные. Тегулы обычно рыжие. Передняя поверхность головы ♀ с серебристым прилегающим опушением. Среднеспинка и переднеспинка обычно более или менее морщинистые, между морщинами пунктированные……………….
……………………………………………………….A. heydeni Dahlb. 36 (37). Задние ноги целиком черные. 1-й членик стебелька брюшка черный. …………………………….………A. heydeni heydeni Dahlb.
37 (36). Задние голени и нижняя сторона задних бедер частично красные. 1-й членик стебелька брюшка красный…………………...
…………………………….……………………..A. heydeni rubra Rad.
38 (35). Ноги и тегулы черные. Передняя поверхность головы ♀ без серебристого опушения. Среднеспинка в середине гладкая и блестящая, по бокам иногда со слабоо-заметными поперечными морщинами.
39 (40). Последний сегмент брюшка с металлическим синим блеском…………………………………………...…………A. sabulosa L.
40 (39). Последний сегмент брюшка черный, без металлического блеска. Длина 3-го тергума брюшка ♂ не менее, чем в 2 раза больше ширины на конце. …….……………………..………….A. assimilis Kohl
41 (34). Промежуточный сегмент полностью покрыт густым серебристым прилегающим опушением, скрывающим скульптуру. ♀ 15, ♂ 11 мм. Узбекистан, Туркменистан. В Казахстане пока не найден.
….……………………………………………………A. argentina Guss.
42 (34). Среднегрудь впереди с бугровидным выступом. Передние тазики с зубчиком на внутренней стороне.
43 (44). Вершина брюшка заостренная. Наличник ♂ без бугорков. Передние тазики ♀ с острым зубчиком. ……….…....A. dentigera Guss.
44 (43). Вершина брюшка закругленная. Наличник ♂ на переднем крае с 2 тупыми, сжатыми с боков бугорками. Передние тазики ♀ с тупым треугольным отростком. ……………….….A. lativalvis Guss.
Описание: Самка. Наличник выпуклый, в передней половине в середине слегка уплощенно-вдавленный, в середине переднего края с узкой глубокой вырезкой, с передней неправильно продольно складчатой каймой (рис. 76з). Пунктировка наличника редкая, некрупная. Промежутки между точками с очень нежной микроскульптурой. Глаза почти параллельные (рис. 76и). Минимальная ширина лица на наличнике слегка меньше ширины лица перед передним глазком. Промежуток между задними глазками примерно в 1,5 раза уже промежутка между одним из них и ближайшим глазом. Лоб вверху не вдавленный, с ясной срединной бороздой. Опушение лица слабо развитое, очень короткое. Воротничок переднеспинки (рис. 76к) короткий (его ширина в середине длины примерно в 2 раза больше длины), с коротким продольным вдавлением сверху, отделенный от среднеспинки глубокой щелевидной перетяжкой. Переднеспинка с заметными поперечными морщинами (особенно по бокам) , с редкими мелкими точками. Эпистернальный шов внизу не развит. Среднеспинка в середине выпуклая, с глубокими боковыми вдавлениями, с поперечными и косыми морщинами по бокам и во вдавлениях. Щитик с негрубыми продольными морщинами, впереди сглаженный. Заднещитик с неправильной морщинистой скульптурой. Срединное поле промежуточного сегмента голое, с косыми морщинами, идущими от средней линии, у основания поперечно выпуклое. Среднегрудь внизу спереди вдавленная, но без бугров и килей. Передние тазики без угловатых выступов на задне-внутреннем крае. Форма маргинальной и субмаргинальных ячеек как на рис. 76л. 1-й членик стебелька по длине равен 1му членику задних лапок вместе с 1/3 2-го. Голова и грудь с длинными светло-серыми волосками. 1-й членик передних лапок, кроме крепкого шипа и 2 щетинок на вершинном выступе, еще с 4 длинными и 1 более короткой базальной щетинкой. Голова, грудь, усики и ноги черные. Стебелек и вершина брюшка черные, 1-й тергум, 2-й и 3-й сегменты и нижняя часть 4-го красно-рыжие. Передние лапки буровато-рыжие, средние и задние – бурые. Крылья затемненные. Жилки коричневые. Длина тела 18-24 мм.
Самец. Наличник почти плоский, слегка вдавленный в середине, относительно широкий (рис. 76м), внизу слегка отогнутый кпереди. Передний край с треугольной выемкой в середине (рис. 76н). Лицо с густым прилегающим серебристо-белым опушением и длинными отстоящими белыми волосками. Ширина лица на наличнике примерно в 1,5 раза меньше, чем перед передним глазком. Промежуток между задними глазками примерно в 1,4 раза уже, чем между одним из них и ближайшим глазом. Ширина воротничка переднеспинки почти в 2,5 раза больше его длины в середине.. Переднеспинка вверху в середине с коротким продольным вдавлением, с поперечными морщинами, с редкими некрупными точками. Среднеспинка однообразно поперечно морщинистая, с отдельными нечетко ограниченными некрупными точками. Скульптура щитика, заднещитика и срединного поля промежуточного сегмента, как у самки. Длинные волоски груди светло-серые. 1-й членик стебелька брюшка почти равен 1-му членику задних лапок вместе с ½ 2го. Членики лапок относительно толстые, отделены друг от друга узкими перетяжками. Голова, грудь, усики и ноги черные. Лапки бурые. 1-й тергум (кроме основания сверху), 2-й и 3-й сегменты и нижняя часть 4го красно-рыжие. Крылья почти не затемненные, с еоричневыми и бурыми жилками. Коста и субкоста бурые. Лапки буроватые. Шипы лапок светло-рыжевато-коричневые. Черная вершина брюшка с синим оттенком, заметным под определенным углом зрения. Длина тела 17-20 мм.
Ареал: Иран, Средняя Азия, Казахстан, Западный Китай.
Распространение в Казахстане: Пустыня Кызылкум – 5-10 км югозап. Чардары; долина Сырдарьи - окр. Чардары; пески Большие Барсуки; плато Устюрт, п-ов Мангышлак; предгорья Заилийского Алатау – Сюгатинская долина; долина р. Или – 35 км сев.-зап. Капчагая; Капчагайское водохранилище – 10 км сев.-вост. Капчагая; Сев. Прибалхашье – г. Балхаш, 14 км сев. Балхаша; Казахский мелкосопочник – Актогай, 35 км южн. и 5 км сев. Актогая, 60 км вост. Джезказгана, 12 км сев. Жармы, 40 км сев.-вост. пос. Кокпекты, 70 км юго-вост. Георгиевки; Зайсанская котловина – г. Зайсан.
Экология и биология: Обитает в пустынях и полупустынях, по долинам рек, в низкогорьях. Встречается на открытых участках с глинистыми, глинисто-щебнистыми и глинисто-песчаными почвами. Биология неизвестна. Самки, вероятно, выкапывают в земле норки, устраивают в них одноячейковые гнезда и запасают в них гусениц бабочек (Lepidoptera). Отмечено питание имаго на молочае, феруле и других растениях.
Синонимы: retusa Gistel, 1848; ? neoxena F.Smith, 1856; ? striaticollis F.Morawitz, 1888; ? nigrina F.Morawitz, 1888.
Подвид: separanda F.Morawitz, 1890. – Юго-восток европейской части России, Казахстан, Средняя Азия.
Описание: Самка. Наличник слабо выпуклый, в передней половине в середине уплощенный, с густой микропунктировкой и редкими мелкими точками, с узкой гладкой каймой впереди, ограниченной по бокам тупыми углами или маленькими зубцами (рис. 77а). Передний край каймы почти прямой. Глаза слабо сближаются книзу (рис. 77а). Ширина лица перед передним глазком примерно в 1,25 раза больше минимальной ширины на наличнике. Воротничок переднеспинки примерно в 2 раза шире своей длины (рис. 77б). Промежутокмежду задними глазками примерно в 1,6 раза уже промежутка между одним из них и ближайшим глазом. Среднеспинка с густой микропунктировкой, редкой некрупной пунктировкой, по бокам с нежными поперечными морщинами. Щитик с продольными морщинами, редкой пунктировкой и сетчатой микроскульптурой. Заднещитик без морщин, с довольно густой пунктировкой и сетчатой микроскульптурой, полуматовый. Срединное поле промежу- точного сегмента голое, косо-морщинистое, в средней части с сетчатой микроскульптурой, по бокам более или менее сглаженное. Среднегрудь снизу спереди без бугорков, выступов и килей. Передние тазики сзади без выступов. 1-й членик передних лапок на вершине с выступом, проксимальный шип которого короче и толще остальных. 3-я субмаргинальная ячейка стебельчатая (рис. 77в). 1-й членик стебелька брюшка по длине равен 1-му членику задних лапок вместе с 1/3 2-го. Прилегающее серебристое опушение головы и груди слабо развитое, только на боках среднегруди и на вершине боков промежуточного сегмента образыет «войлочные» пятна. Голова, грудь, усики и ноги черные. Брюшко частично красно-рыжее. У типичного подвида 1-й тергум, 2-й сегмент и большая часть 3-го красно-рыжие. Крылья с буроватым затемнением. Отстоящие волоски и щетинки головы и переднегруди снизу чернобурые, на остальной части груди светло-серые. Тазики сзади бурые. Плечевыебугры почти без прилегающего опушения. Длина тела 13-20 мм.
Самец. Передний край наличника почти прямой или слегка выемчатый (рис. 77г). Наличник слабо выпуклый, с относительно слабо развитым прилегающим серым опушением. Глаза сильно сближаются книзу (рис. 77д). Ширина лица перед передним глазком почти в 1,8 раза больше минимальной ширины на наличнике. Промежуток между задними глазками примерно в 3 раза уже промежутка между одним из них и ближайшим глазом. Голова с длинными бурыми отстоящими волосками. Переднеспинка с редкой мелкой пунктировкой, с сетчатой микроскульптурой, в середине слегка продольно вдавленная. Ширина воротничка в средней части примерно в 2 раза больше его длины. Среднеспинка с сетчатой микроскульптурой, нечеткой мелкой пунктировкой и поперечными (сзади косыми) негрубыми морщинами. Срединная борозда среднеспинки хорошо выражена, идет почти до заднего края. Щитик и заднещитик с продольными морщинами. Срединное поле промежуточного сегмента на всей поверхности с густыми поперечными (в середине изломанными) морщинами, с сетчатой микроскульптурой, по крайней мере, в средней части. Грудь без участков густого прилегающего опушения. Передние тазики сзади изнутри бугровидно выпуклые, но без острых выступов. Среднегрудь впереди внизу без килей, зубцов и выступов. 3-я субмаргинальная ячейка стебельчатая. Голова, грудь, ноги и усики черные. 1-й тергум, 2-й и 3-й сегменты брюшка красно-рыжие, часто с темными пятнами. Остальная часть брюшка черная. Крылья слегка буроватые. Жилки бурые. Грудь с длинными буровато-серыми отстоящими волосками. Вершина промежуточного сегмента со сгущенным прилегающим опушением. Длина тела 12-18 мм.
Подвид A. c. separanda отличается от типичного подвида, описанного выше, целиком (у самца только на вершине) красно-рыжим первым члеником стебелька. У него также сильнее развито прилегающее опушение (в частности, на плечевых буграх и на промежуточном сегменте, особенно сзади около срединного поля), волоски на голове светлые и срединное поле между косыми морщинами гладкое и блестящее, по крайней мере, по бокам.
Ареал: Почти вся Европа, Юго-Западная Азия, Средняя Азия, Казахстан, Монголия, Китай, Япония, Сибирь и Дальний Восток России.
Распространение в Казахстане: A. c. campestris Latr. – предгорья и низкогорья Сев. Тянь-Шаня, Саура, Тарбагатая и Алтая; БалхашАлакольская впадина, Зайсанская котловина, Казахский мелкосопочник, Северо-Казахская равнина, Кокчетавская возвышенность, Тургайская ложбина, Южное Предуралье, долина р. Урал, Рын-пески. A. c. separanda F.Mor. – долина р. Или, Прибалхашье, долина р. Сырдарьи, предгорья хр. Каратау, пуст. Кызылкум, пуст. Мойинкум, пуст. Сексеулдала, долина р. Урал.
Биология и экология: Эврибионтный вид; встречается почти повсеместно от лесной зоны до пустынь, от альпийских лугов до предгорных равнин. Самки гнездятся в глинистой или песчаной уплотненной почве. Добыча – гусеницы бабочек (Gejmetridae, Pieridae, Noctuidae) и личинки пилильщиков (Tenthredinidae). В одной ячейке запасается до 6 экземпляров добычи. Отмечено питание взрослых ос на цветках мяты, синеголовника, лука, вероники, тамариска, селитрянки, молочая, саусуреи, кермека, крестовника, клоповника, жузгуна, мирикарии, морковника.
Описание (по В.В.Гуссаковскому – Gussakovskij, 1930): Самка. Голова поперечная (см. спереди); глаза слегка сближаются книзу. Наличник слабо выпуклый, передний край его узко выемчатый в середине, в целом почти равномерно закругленный. Лоб полуматовый, с редкой пунктировкой. Воротничок переднеспинки поперечный, в середине вверху слегка вдавленный, только впереди слабо поперечно морщинистый. Среднеспинка по бокам с поперечными, сзади с косыми морщинами, с некрупной пунктировкой. Щитик со слабыми продольными морщинами. Мезоплевры и бока промежуточного сегмента почти матовые из-за микроскульптуры. Эпистернальный шов хорошо развитый. Среднегрудь снизу впереди без бугров и вдавления. Передние тазики без выступа. Срединное поле промежуточного сегмента с густыми косыми негрубыми морщинами, перед вершиной слегка уплощенно-вдавленное. 1-й членик стебелька брюшка несколько длиннее 1-го тергума. 3-1 членик усиков вдвое длиннее 4-го. 3-я субмаргинальная (кубитальная - ?) ячейка стебельчатая. Толо черное. Лицо с густым серебристым прилегающим опушением. Тегулы и ноги (кроме тазиков, задних голеней и лапок) красно-рыжие, брюшко красно-рыжее, кроме 2 затемненных вершинных сегментов. Крылья затемненные. Длина тела 13 мм. Самец неизвестен.
Ареал не выяснен. Вид описан по одному экземпляру из Северного Казахстана (Павлодар).
Распространение в Казахстане: Прииртышье.
Экология и биология не изучены. Самки, вероятно, устраивают одноячейковые гнезда в земле и охотятся на гусениц бабочек (?).
Описание: Самка. Наличник равномерно выпуклый, с узкой каймой впереди, выемчатой в середине и ограниченной по бокам тупыми углами (рис. 77е), с густым прилегающим серебристым опушением, с небольшим треугольным голым участком в середине у переднего края. Глаза сближаются книзу (рис. 77е). Промежуток между ними у переднего глазка в 1,3 раза шире, чем на наличнике….. Лоб с густым прилегающим серебристо-белым опушением, скрывающим скульптуру. Переднеспинка удлиненная (рис. 77ж), впереди без резкого уступа, без поперечных морщин, с густым прилегающим опушением. Среднеспинка без грубых поперечных морщин, с густым (особенно по бокам) прилегающим серебристым опушением, без срединной борозды, в середине слегка вдавленная. Срединное поле промежуточного сегмента с серебристым прилегающим опушением на всей поверхности. Среднегрудь снизу в середине впереди с бугорком, ограничивающим вогнутую переднюю поверхность. Передние тазики с зубцом на верхне-внутренней поверхности. 1-й членик передних лапок на вершине апикального выступа с 1 крепким темным шипом и длинными крепкими щетинками (рис. 77з). 1й членик стебелька по длине равен 1-му и 2-му членикам задних лапок вместе. Мандибулы, передняя часть наличника, скапус, переднегрудь на большей части, тегулы, промежуточный сегмент на вершине, брюшко и ноги рыжие или красно-рыжие. Длина тела 14-19 мм.
Самец. Наличник выпуклый, в середине до переднего края слегка уплощенно-вогнутый, с узкой гладкой каймой впереди, с почти прямым передним краем (рис. 77и). Глаза сильно сближаются книзу (рис. 77и). Ширина лица у переднего глазка примерно в 1,6 раза больше, чем на наличнике. Промежуток между задними глазками примерно в 1,4 раза уже, чем между одним из них и ближайшим глазом. Лицо с густым серебристым опушением, скрывающим скульптуру, и отдельными светлыми отстоящими волосками. Переднеспинка удлиненная (рис. 77к), без резкого уступа спереди, без грубых поперечных морщин. Среднеспинка с густой сетчатой микроскульптурой, без четких точек и грубых морщин, с прилегающим опушением. Щитик выпуклый, в середине уплощенный, без продольных морщин. Заднещитик бугровидно выпуклый, без морщин. Срединное поле промежуточного сегмента неправильно негрубо морщинистое, с густым прилегающим опушением, без голых участков. Среднегрудь спереди внизу с бугром, ограничивающим переднюю вогнутую поверхность. Средние бедра с угловатым выступом на задневнутреннем крае. 3-я субмаргинальная ячейка впереди слабо суженная, не стебельчатая. 1-й членик стебелька брюшка очень тонкий, в 14-15 раз длиннее своей ширины в середине, слегка длиннее общей длины 1-го и 2-го члеников задних лапок. Мандибулы, передний край наличника, задний край плечевых бугров, тегулы, ноги на большей части и брюшко на большей части красно-рыжие. Скапус спереди буровато-рыжий, сзади бурый. Жгутик усиков черный. 1-й членик стебелька, 1-й тергум сверху, 5-й тергум сверху, 6-й и 7-й сегменты брюшка черные. Задние тазики почти целиком, задние вертлуги и задние бедра отчасти (сверху) черные или черно-бурые. Задние голени изнутри, средние тазики, вертлуги и основание средних бедер иногда бурые. Лапки красно-рыжие. Крылья не затемненные. Жилки рыжевато-коричневые или буровато-коричневые.
Длина тела 14-17 мм.
Ареал: Иран, Средняя Азия, Казахстан.
Распространение в Казахстане: Пустыня Сексеулдала – 50-70 км сев.-вост. Фурмановки; долина р. Или – 65 км зап. Баканаса, 35 км сев. Чилика; Зайсанская котловина – 5-15 км сев.-вост. Каратала, 52 км сев.
Зайсана.
Экология и биология: Вид обитает в песчаных и песчаносолончаковых пустынях. Самки роют норки в земле, причем частицы субстрата относят от гнезда на лету на расстояние до 40-50 см. Гнездо одноячейковое, но около ячейки имеется короткий слепой придаток (отнорок). Его назначение неясно. Возможно, он образуется вследствие того, что оса берет здесь материал для закрывания ячейки (подобное явление отмечено у Prionyx niveatus). Добыча – гусеницы бабочек (Lepidoptera, ? Noctuidae). Отмечено питание имаго на цветках кермека и качима.
Описание: Самец. Передний край наличника широкий, слегка выемчатый (рис. 78а). Наличник почти плоский, впереди слегка отогнутый кпереди, с густым серебоисто-белым опушением, с длинными отстоящими серовато-белыми волосками. Глаза сильно сближаются книзу (рис. 78а). Ширина лица перед передним глазком примерно в 1,75 раза больше, чем на наличнике. Промежуток между задними глазками примерно в 1,2 раза уже, чем между одним из них и ближайшим глазом. Переднеспинка удлиненная, воротничок впереди в середине вдавленный, с резко выступающим косо вверх передним краем, глубоко выемчатым в середине (рис. 78б). Затылок с высоким килем. Поперечные морщины переднеспинки грубые. Среднеспинка с грубыми поперечными (слегка скошенными) морщинами. Щитик м заднещитик с ячеисто- или продольно-морщинистой грубой скульптурой. Срединное поле промежуточного сегмента с грубымит косыми, в средней части неправильно изогнутыми морщинами, по бокам голое, в середине с короткими прилегающими и длинными отстоящими волосками. Форма маргинальной и 2 субмаргинальных ячеек как на рис. 78в. 1-й членик стебелька брюшка слегка длиннее совместной длины 1-го и 2-го члеников задних лапок. Опушение груди хорошо развитое, но сверху не скрывает скульптуру. Голова черная. Мандибулы в середине рыжеватые. Скапус на вершине буроватый. Грудь черная. Тегулы рыжие. Вершина плечевых бугров рыжевато-бурая. Крылья слегка буроватые, с темно-коричневыми (у основания с рыжими) жилками. Ноги черно-бурые. Вершинная часть передних (иногда и средних) бедер и передних голеней более или менее рыжая и рыжевато-бурая. 1-й членик стебелька брюшка черный, на вершине рыжий. 1-й тергум, 2-й сегмент целиком и большая часть 3-го сегмента брюшка красно-рыжие. 4-й тергум рыжий только по бокам и на нижне-передних частях, 4-й стернум (иногда и 5-й) рыжий на переднебоковых частях. Усики черные. Длина тела 15-17 мм.
Самка автору не известна. Возможно, часть самок, определенных автором как A. occipitalis F.Mor., в действительности относятся к этому виду.
Ареал: Юго-восток европейской части России, Туркменистан, Казахстан, Монголия.
Распространение в Казахстане: Рын-пески; сор Кайдак; присырдарьинская равнина – Байгакум, Баламурун, 5 км вост. Чардары; Южное Предуралье – окр. Актюбинска; Тургайская ложбина – 30 км юго-зап. Иргиза, окр. пос. Шиили, 25 км зап. пос. Чиили, 40 км юго-зап. Аркалыка; долина р. Или – 35 км сев.-зап. Капчагая, 60 км вост. пос. Или (бывш.), окр. пос. Или; Прибалхашье – Егеньсу, 15 км юго-зап. Балхаша; Зайсанская котловина – 72 км юго-вос. пос. Кокпекты, 170 км сев.-зап. Зайсана, 52 км сев.-вост. Зайсана, 45 км южн. Каратогая, 5-15 км сев.вост. и 20 км вост. Каратала; 7 км зап. Черняевки; Прииртышье – 10 км сев. Лебяжьего, Белогорье, предгорья Заилийского Алатау – Тескенсу, Бартогай (на р. Чилик), р. Чарын.
Экология и биология: Пустынно-степной вид. Предпочитает участки с более или менее рыхлыми песчаными и супесчаными почвами. Биология неизвестна. Самки, вероятно, устраивают одноячяейковые гнезда в земле и охотятся на гусениц бабочек (Lepidoptera).
Синонимы: longicollis Kohl, 1884; debilis F.Morawitz, 1889; producticollis Morice, 1900 (♂).
Описание: Наличник слегка равномерно выпуклый, с гладкой блестящей каймой впереди, ограниченной по бокам угловатыми выемками. Передний край каймы или слегка, почти равномерно дугообразно выступающий, или почти прямой (рис. 78г), или слегка выемчатый в середине. Срединная лопасть наличника с небольшим голым участком впереди, на остальной поверхности с густым серебристо-белым прилегающим опушением, скрывающим скульптуру, с отдельными белыми или светло-серыми отстоящими волосками. Мандибулы как на рис. 78д. Лицо с густым прилегающим опушением. Лоб с глубокой срединной бороздой (продольно вдавленный). Внутренние края глаз почти параллельные (рис. 78е). Промежуток между задними глазками в 1,4 раза уже, чем между одним из них и ближайшим глазом. Темя в середине бугровидно выпуклое. Воротничок переднеспинки удлиненный, спереди круто обрывающийся, с редкими поперечными морщинами (рис. 78ж,з), с довольно густым прилегающим опушением. Среднеспинка с грубыми поперечными неправильно изогнутыми морщинами, сзади по бокам и в середине сильно продольно вдавленная. Щитик и заднещитик с неправильно изогнутыми грубыми продольными морщинами, в середине с продольным вдавлением. Среднегрудь снизу с 2 косыми килями, оканчивающимися внизу зубцом, ограничивающим вогнутую поверхность. Передние тазики на задне-внутренней поверхности с угловатым выступом. 1-й членик передних лапок на конце вытянут в выступ, несущий одну крепкую шипообразную темно-бурую щетинку и 2 более тонкие уплощенные бурые щетинки (иногда 3 почти одинаковые щетинки). Срединное поле промежуточного сегмента с неправильной грубой морщинистой скульптурой, целиком опушенное. 3-я субмаргинальная ячейка впереди сильно суженная (рис. 78и). Голова и грудь на большей части черные. Мандибулы на большей части, наличник впереди, скапус, плечевые бугры, тегулы, часто вершина промежуточного сегмента, ноги на большей части и брюшко на большей части красно-рыжие. Крылья почти не затемненные. Жилки коричневые, у основания крыльев рыжеватые. Субкоста бурая. 1-й членик стебелька черный, иногда более или менее рыжий. Брюшко с черной вершиной (6-й и отчасти 5-й сегменты). Задние тазики, вертлуги и бедра отчасти черные сверху. Усики черные, отчасти бурые. Длина тела 16-22 мм.
Самец. Наличник вверху слабо выпуклый, в переднкй половине поперечно вогнутый, на переднем крае в середине слегка широко тупоугольно выемчатый. Лицо (включая наличник) с густым серебристым прилегающим опушением, скрывающим скульптуру. Глаза сильно сближаются книзу (рис. 78к). Промежуток между задними глазками примерно в 2,2 раза уже, чем между одним из них и ближайшим глазом. Воротничок переднеспинки удлиненный. Переднеспинка и среднеспинка с очень грубыми поперечными морщинами, не скрытыми под опушением. Бока груди с густым прилегающим опушением. Среднегрудь впереди внизу с каждой стороны близ передних тазиков с косым складкообразным килем, оканчивающимся внизу угловатым выступом. Передние тазики с угловатым выступом сзади изнутри. Срединное поле промежуточного сегмента с неправильной морщинистой скульптурой, с коротким полуприлегающим светлым опушением. Голова и грудь черные. Мандибулы на большей части желтовато-рыжие. Тегулы и отчасти плечевые бугры рыжие. Ноги более или менее рыжие. Лапки, тазики, задние голени, задние бедра, передние вертлуги отчасти черно-бурые. Передние лапки обычно рыжие. Усики черные. 1-й сегмент брюшка (включая 1-й тергум) черный. 1-й тергум по бокам, 2-й стернум, 3-5-й сегменты брюшка более или менее красно-рыжие. Крылья слабо затемненные, передние слегка буроватые. Жилки бурые и темно-коричневые. Вершина брюшка черная, без металлического блеска. Длина тела 14-18 мм.
Ареал: Юго-Восточная Европа, Северная, Центральная и Восточная Африка, Юго-Западная Азия, Средняя Азия, Казахстан, Монголия.
Распространение в Казахстане: Пустыня Кызылкум и Присырдарьинские пустыни – 5-8 км зап., 2-8 км юго-зап., 30 км южн. и 5 км вост. Чардары, Отрар, Коксу, Байгакум (близ Чиили), Джулек (близ Чиили), Камышлыбаш; пуст. Мойинкум – 30 км южн. Чу, Уланбель; пуст. Бетпакдала – 50-60 км сев.-зап. Фурмановки; долина р. Или – 5-6 км южн. Капчагая, 35 км сев.-зап. Капчагая, Или (бывш.), 8 км южн. и 10 км юго-зап. Жаркента (бывш. Панфилов), Аяк-Калкан, 15 км вост. АякКалкана, Басший, 17 км сев.-зап., 35 км юго-вост. и 65 км зап. Баканаса, 8 км сев. пос. Екпинды; Южное и Сев. Прибалхашье – Акколь, Егеньсу, 60 км юго-вост. Копберлика, Балхаш, 5-15 км юго-зап. Балхаша; пуст.
Сарытаукум – Айдарлы; низовья р. Чарын; Южное Предуралье – Саралжин; Прикаспийская низменность – Атырау (Гурьев), Урда; Тургайская ложбина – Иргиз, 5 км южн. Иргиза; Большие Барсуки – 12 км сев.-вост. Челкара; Прииртышье – 40 км сев. Семипалатинска, Лебяжье; СевероКазахстанская равнина – 100 км зап. Павлодара.
Экология и биология: Пустынный вид. Обычен на песках. Биология не изучена. Самки, вероятно, устраивают одноячейковые гнезда в песчаном грунте и запасают в них гусениц бабочек (Lepidoptera). Отмечено питание имаго на цветках молочая, тамариска. Качима, селитрянки, ак-курая, адраспана, кермека, саусуреи, липучки, лука, астрагала, клоповника, солянок и др. растений.
Синонимы: ? attenuata Christ, 1791 (Sphex); ? rubra Radoszkowski, 1876 (? = quadraticollis A.Costa, 1893); rubra Radoszkowski, 1877, nec Radoszkowski, 1876; iberica Ed.Andre, 1886.
Подвиды: rubriventris A.Costa, 1864 (Сицилия, Корсика); sarda Kohl, 1906 (Сардиния, Кипр).
Описание: Самка. Наличник почти равномерно выпуклый. Передний край срединной лопасти наличника почти прямой (рис. 79а) между боковыми углами или зубцами (рис. 79а,б,в). Пунктировка мелкая, довольно редкая. Сетчатая микроскульптура нечеткая. Опушение слабо развитое, не скрывающее скульптуру. Лоб широко клинообразно вдавленный, со срединной бороздкой. Глаза почти параллельные или слегка сближенные внизу (рис. 79б,в). Промежуток между задними глазками почти в 1,5 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка не удлиненная, более чем в 2 раза шире своей длины (рис. 79г), с редкой мелкой пунктировкой и нежной микроскульптурой. Среднеспинка с равномерной поперечно-морщинистой, не очень грубой скульптурой, с редкими точками. Срединная борозда не выражена или слабо выражена. Щитик и заднещитик с продольными густыми морщинами. Бока среднегруди и промежуточного сегмента с пятнами густого прилегающего серебристого опушения. Срединное поле промежуточного сегмента с неправильной (преимущественно поперечной) морщинистой скульптурой, нечетко выраженной пунктировкой, грубой микроскульптурой и негустыми отстоящими волосками, без голых блестящих участков. 1-й членик стебелька брюшка слегка длиннее 1-го и 2го члеников задних лапок. 1-й членик передних лапок с выступом на вершине, несущим крепкий черный шип и 1-2 крепкие щетинки, и 4 крепкими щетинками на наружном крае (рис. 79д). Среднегрудь снизу впереди без бугров, зубцов и килей. Голова, грудь и усики черные. Скапус буроватый у вершины. Передние и средние ноги (кроме тазиков и вертлугов) красно-рыжие, задние черные. Задние лапки часто красновато-бурые. Крылья с явным буроватым затемнением, с бурыми и коричневыми жилками. 1-й членик стебелька и основание 2-го черные. 4-й сегмент брюшка на большей части, 5-й и 6-й сегменты черные. Остальная часть брюшка красно-рыжая. Длина тела 18-24 мм.
Самец. Наличник слегка выпуклый, с густым серебристым прилегающим опушением и несколькими светлыми отстоящими волосками. Его передний край в середине с маленькой выемкой. Глаза сильно сближаются книзу (рис. 79е). Промежуток между задними глазками примерно в 1,1 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка с довольно крупной и довольно густой пунктировкой, с прилегающим опушением и отстоящими светло-серыми водлосками, в середине не вдавленная. Среднеспинка грубо неправильно морщинистопунктированная, с равномерным опушением и отстоящими волосками. Щитик и заднещитик с изогнутыми продольными морщинами. Срединное поле промежуточного сегмента с неправильной ячеистоморщинистой скульптурой, с коротким полуприлегающим опушением (не скрывающим скульптуру) и длинными отстоящими волосками, без голых блестящих боковых участков. Плечевые бугры с густым прилегающим опушением. 1-й членик стебелька по длине примерно равен 1му, 2-му членикам задней лапки и ½ 3-го. Голова и грудь черные. Мандибулы и усики черные. Передние и средние бедра (кроме черного основания), передние и средние голени и базальная часть передних и средних лапок красно-рыжие. Тазики, вертлуги, основание передних и средних бедер, задние ноги целиком черные, иногда отчасти буроватые. Средние лапки на большей части буроватые. Стебелек брюшка черный или бурый. 1-й тергум сверху с продольной черной полосой. Вершина брюшка черная (4-й тергум сверху и 5-7-й сегменты целиком). Длина тела 16-22 мм.
Ареал: Южная и Средняя Европа, Северная Африка, ЮгоЗападная Азия, Средняя Азия, Казахстан, Западный Китай.
Распространение в Казахстане: Почти повсеместно. Обнаружен в пустыне Кызылкум, присырдарьинских пустынях, в долине р. Сырдарьи, в предгорьях и низкогорьях Западного и Северного Тянь-Шаня, в пустыне Мойинкум, Прибалхашье, долине р. Или, Казахском мелкосопочнике, Южном Предураье, Прикаспийской низменности, Тургайской ложбине, на п-ове Мангышлак, в Прииртышье, предгорьях и низкогорьях Алтая, Саура и Тарбагатая. Один из самых обычных видов рода Ammophila.
Вид на территории Казахстана представлен двумя формами, которые различаются окраской ног и брюшка. Типичная форма описана выше. Распространена она на большей части Казахстана. У второй формы, вероятно, соответствующей подвиду A. h. rubriventris, брюшко целиком или почти целиком красно-рыжее и задние ноги тоже частично краснорыжие. Обнаружена на п-ове Мангышлак, в пустынях Южного и ЮгоВосточного Казахстана.
Экология и биология: Эврибионтный пустынно-степной вид. Обитает в пустынях, полупустынях и степях всех типов. Нередок по долинам рек, но густых древесно-кустарниковых зарослей избегает. Самки устраивают одноячейковые гнезда в земле и запасают в них гусениц пядениц (Geometridae) или ночниц (Noctuidae) – по несколько экземпляров в каждой ячейке. Отмечено питание взрослых ос на цветках кермека, молочая, смолевки, тамариска, солнцецвета, вероники, мирикарии, качима, зизифоры, крестовника, клоповника, мяты, синеголовника, морковника и других зонтичных, мордовника, лука, ластовня, саусуреи, селитрянки, адраспана, верблюжьей колючки, подмаренника, сурепки, гораниновии.
Синонимы: turcica Mocsary, 1883; hispanica Mocsary, 1883; fallax Kohl, 1884.
Описание: Самка. Наличник вблизи переднего края с выступающей поперечной складкой (рис. 79ж,з), в целом слабо выпуклый, с редкой пунктировкой и густой микропунктировкой, со слабо развитым коротким прилегающим опушением преимущественно в верхней части. Передний край срединной лопасти наличника почти прямой, с закругленными боковыми углами (рис. 79з). Лицо широкое (рис. 79ж). Глаза сближаются книзу, на внутреннем крае слегка выемчатые. Промежуток между задними глазками в 2 раза уже промежутка между одним из них и ближайшим глазом. Лоб со срединной бороздкой. Нижний участок лба (с усиковыми ямками) сильно вдавленный. Переднеспинка сильно выпуклая, укороченная (рис. 79и). Длина воротничка более чем в 2 раза меньше его ширины в середине. Передняя поверхность воротничка круто спускающаяся к шейке (рис. 79и). Верх переднеспинка без продольного вдавления в середине. Переднеспинка отделена от среднеспинки глубокой щелевидной перетяжкой (рис. 79и). Среднеспинка с сильно выпуклой (особенно в задней половине) средней частью. Переднеспинка и среднеспинка с редкой мелкой пунктировкой, густой микропунктировкой и сетчатой микроскульптурой, полуматовые. Бока переднеспинки сбоку перед плечевыми буграми сильно вдавленные. Среднеспинка на выпуклой средней части, особенно сзади, гладкая и блестящая. Щитик с нежными продольными морщинами в задней части, с редкими мелкими точками. Срединное поле промежуточного сегмента с косыми морщинами, на большей части почти голое (лишь с редкими короткими волосками), в середине с довольно длинными отстоящими волосками. Бока среднегруди внизу без эпистернального шва. Низ среднегруди с продольным срединным низким килем, без бугров и зубцов впереди. Маргинальная и субмаргинальные ячейки как на рис. 80д. 1-й членик стебелька брюшка немного длиннее 1-го членика задних лапок. 1-й членик передних лапок с 1 коротким и толстым и 2 длинными и тонкими шипами-щетинками на конце вершинного выступа. Коготки с зубчиком (рис. 79к). Голова, грудь,ноги и усики черные. Мандибулы черные, отчасти бурые. Лапки отчасти (главным образом на вершине члеников) буроватые. Брюшко частично красное (1-й тергум, 2-й и 3-й сегменты). Основание 1-го тергума обычно черное. Крылья с буроватым, почти равномерным затемнением. Жилки бурые и коричневые. Длина тела 1724 мм.
Самец. Наличник плоский, с почти прямым передним краем между боковыми углами (рис. 79л). Глаза сильно сближаются книзу (рис. 79л). Ширина лица перед передним глазком примерно в 2 раза больше минимальной ширины на наличнике. Промежуток между задними глазками примерно в 1,6 раза уже промежутка между одним из них и ближайшим глазом. Лицо с густым прилегающим серым опушением. Голова с длинными отстоящими черными волосками. Воротничок переднеспинки более чем в 2 раза шире своей длины. Переднеспинка без морщин и четко выраженных точек, с довольно грубой шероховатой микроскульптурой. Среднеспинка с нежными косыми морщинами по бокам, особенно четкими сзади. Щитик равномерно выпуклый, с нежными продольными морщинами. Заднещитик с нежными продольными морщинами и срединным килем. Эпистернальный шов не развит. Срединное поле промежуточного сегмента с косыми нежными морщинами, с сетчатой микроскульптурой, котороая по бокам слабее выражена, с длинными отстоящими серыми волосками, без прилегающего опушения. 1-й членик стебелька короче 1-го и 2-го члеников задней лапки. Голова, усики, грудь, ноги и брюшко на большей части черные. 2-й сегмент брюшка и основание 3-го по бокам и снизу красно-рыжие. Крылья со слабым буроватым затемнением. Жилки бурые и коричневые. Волоски гололвы черные и бурые, волоски груди серые. Длина тела 15-23 мм.
Ареал: Южная и Средняя Европа, Северная Африка, Юго-
Западная Азия, Средняя Азия, Казахстан.
Распространение в Казахстане: Низкогорья и предгорья Северного и Западного Тянь-Шаня – Бурное, Кентау, Подгорное (Киргизский хр.), Ачисай, 20 км вост. Ачисая, 30 км сев.-зап. Ванновки, Подгорное (хр.
Кетмень), Узунагач, Алматы, Жаркент; Казахский мелкосопочник – горы Акчатау.
Экология и биология: Встречается в степных горных и предгорных биотопах и в горном лесном поясе. Самки гнездятся в плотном грунте и устраивают одноячейковые гнезда. Добыча - гусеницы бабочек (Lepidoptera), в частности, волосатые гусеницы.
Ammophila iliensis Kazenas, nom. n.
Синоним: tsunekii Kazenas, 1978 (nec Menke, 1976).
Описание: Самка. Наличник равномерно выпуклый. Передний край каймы срединной лопасти наличника почти прямой между боковыми углами (рис. 80а). Опушение наличника густое, прилегающее, на большей части скрывающее скульптуру, сереблисто-белое. Передняя (голая) часть наличника с густой микропунктировкой на гладком и блестящем фоне. Лоб с густым серебристо-белым опушением, скрывающим скульптуру. Глаза ясно сближаются книзу (рис. 80а). Минимальное расстояние между глазами на наличнике в 1,3 раза меньше ширины темени на уровне переднего края переднего глазка (6,5 : 4,9). Темя с редкой мелкой пунктировкой, нечеткой микроскопической сетчатой скульптурой и микропунктировкой. Отношение расстояния между одним из глазков и ближайшим глазом к расстоянию между задними глазками равно 1,2. 3-й членик усиков почти в 2 раза длиннее 4-го. Последний членик усиков приблизительно в 2 раза длиннее своей ширины, на конце усеченный. Переднеспинка (рис. 80б,в) вверху у заднего края слегка продольно вдавленная, впереди и по бокам более или менее опушенная, вверху голая, с редкой пунктировкой и нечеткой микроскульптурой, с микропунктировкой. Среднеспинка на большей части голая, по краям более или менее опушенная, с довольно редкой крупной пунктировкой (точки в 3,5-4 раза уже переднего глазка; промежутки между ними уже, равны или шире точек), с микропунктировкой и очень нечеткой сетчатой микроскульптурой. Скульптура щитика, как на среднеспинке, только в задней части развиты продольные складки. Бока и низ груди с густым прилегающим серебристо-белым опушением, почти полностью скрывающим скульптуру. Дорсальная поверхность промежуточного сегмента с неправильными складками и пунктировкой, покрытая беспорядочно направленными короткими прилегающими серебристыми волосками и редкими отстоящими длинными волосками. 1-й членик передних лапок как на рис. 80г. Коготки без зубцов. Форма 2-й и 3-й субмаргинальных ячеек как на рис. 80д. Голова и грудь с длинными отстоящими белыми волосками. 1-й членик брюшка (1-й стернум) почти равен 2-му (1-му тергуму), в 1,2 раза короче задней голени и в 1,5 раза длиннее 1-го членика задней лапки. Тергумы брюшка с чрезвычайно нежной сетчатой микроскульптурой. Голова и грудь черные. Мандибулы на большей части, передняя кайма наличника (не всегда), тегулы рыжие. Усики черные или бурые, скапус на передне-внутренней поверхности рыжий. Жилки крыльев бурые, у основания крыльев рыжие. Брюшко на большей части рыжее. 1-й членик стебелька почти целиком и 2-й сверху от основания черные. Вершина 6-го тергума буроватая. Ноги на большей части рыжие. Тазики и задние вертлуги почти целиком черные. Длина тела 15-17 мм.
Самец. Глаза сильно сближаются книзу (рис. 80е). Минимальное расстояние между глазами на наличнике в 1,8 раза меньше ширины темени на уровне переднего края переднего глазка. OOL : POL = 1,25-1,3. 3-й членик усиков в 1,6 раза длиннее 4-го и в 4 раза длиннее своей ширины на конце. Последний членик усиков на конце прямо усеченный, в 2,2 раза длиннее своей ширины. 4-13-й членики усиков с продольной выступающей линией на внутренней поверхности. Опушение и скульптура тела, как у самки, только переднеспинка и среднеспинка с более развитым опушением. 1-й членик брюшка (1-й стернум) в 1,1 раза длиннее 2-го (1-го тергума), почти в 1,3 раза короче задней голени и приблизительно в 1,6 раза длиннее 1-го членика задней лапки. Окраска тела, как у самки, только мандибулы почти целиком бурые, наличник черный, 5-й тергум брюшка сверху с черным пятном, 6-й тергум и 7-й сегмент брюшка черные. Длина тела 16-17 мм.
Ареал: Юго-Восточный Казахстан.
Распространение в Казахстане: Долина р. Или – 15 км вост. АякКалкана, Барохудзир (30 км юго-зап. Коктала).
Экология и биология: Обитатель песчаных и солончаковых пустынь. Встречается по долинам рек. Биология не изучена. Самки, вероятно, устраивают одноячейковые гнезда в земле и охотятся на гусениц бабочек (Lepidoptera).
Описание: Самка. Срединная лопасть наличника с узкой голой и гладкой каймой впереди, имеющей выемку в середине между 2 зубцеобразными тупыми выступами (рис. 80ж), с голым участком впереди, с густым прилегающим серебристым опушением на остальной поверхности, довольно сильно выпуклая. Мандибулы с маленьким зубчиком между апикальным и 2 проксимальными зубцами. Глаза почти параллельные (рис. 80ж). Лоб слабо вдавленный в середине, с густым прилегающим серебристым опушением. Промежуток между задними глазками примерно в 1,4 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка удлиненная, с небольшим бугорком в середине, без уступа впереди (рис. 80з,и), без грубых поперечных морщин, с нежной сетчатой микроскульптурой, отдельными мелкими точками и коротким прилегающим (не очень густым) опушением. Среднеспинка с редкой некрупной пунктировкой, нежной сетчатой микроскульптурой и коротким прилегающим (густым по бокам) опушением. Срединная борозда наиболее глубокая в средней части среднеспинки. Щитик сильно выпуклый, в середине с продольным вдавлением, без морщин. Заднещитик сильно бугровидно выпуклый, в середине слегка продольно вдавленный. Срединное поле промежуточного сегмента с неправильной морщинистой скульптурой, с опушением, только по бокам с голыми, косо морщинистыми участками. Среднегрудь снизу впереди с вогнутым участком, ограниченным сзади бугром. Передние тазики с остроугольным выступом на задне-внутреннем крае. Форма маргинальной и субмаргинальных ячеек как на рис. 80к. Выступ 1-го членика передних лапок с 1 коротким кривым и толстым шипом и 2 длинными крепкими щетинками. 1-й членик стебелька брюшка слегка длиннее 1-го и 2-го члеников задних лапок. Голова и грудь черные. Мандибулы на большей части, наличник впереди, скапус, плечевые бугры, тегулы, ноги на большей части и брюшко красновато-рыжие. 1-й тергум сверху отчасти черно-бурый. Крылья почти не затемненные, с коричневыми (у основания крыльев с рыжеватыми) жилками. Задние лапки целиком, средние отчасти бурые.
Задние вертлуги и задние бедра отчасти бурые. Длина тела 15-18 мм.
Самец. Наличник удлиненный, в базальной половине бугровидно выпуклый. Его передний край в середине без вырезки или с небольшой вырезкой. (рис. 80л). Лицо узкое (рис. 80л). Промежуток между задними глазками примерно равен промежутку между одним из них и ближайшим глазом. Лицо с густым прилегающим серебристо-белым опушением, скрывающим скульптуру, и отдельными отстоящими светло-серыми волосками. Переднеспинка удлиненная, впереди без вертикального уступа. Переднеспинка и среднеспинка без грубых поперечных морщин. Щитик в середине продольнго вдавленный. Заднещитик сильно бугровидно выпуклый, в середине с продольным вдавлением. Среднегрудь снизу впереди вогнутая, с широким бугром, ограничивающим вогнутую поверхность. Срединное поле промежуточного сегмента почти на всей поверхности негрубо поперечно неправильно морщинистое и опушенное, лишь по бокам с более правильной поперечной морщинистой скульптурой и голое. Передние тазики бугровидно выпуклые изнутри, но без зубца. 7-й тергум брюшка широкий (рис. 80м), перед вершиной вогнутый (см. в профиль – рис. 80н). Голова и грудь черные. Мандибулы на большей части, передняя часть наличника, скапус, ноги на большей части, тегулы, вершины плечевых бугров, вершина промежуточного сегмента и брюшко на большей части рыжие или красно-рыжие. Жгутик усиков черный. 1-й тергум брюшка сверху черный. Лапки на большей части или целиком черно-бурые. Длина тела 13-18 мм.
Ареал: Средняя Азия, Южный Казахстан.
Распространение в Казахстане: Бетпакдала (Сексеулдала) – 50-70 км сев.-зап. Фурмановки.
Экология и биология: Пустынный вид. Самки устраивают одноячейковые гнезда в неглубоких норках в уплотненном песчаном или супесчаном грунте. При рытье норки осы выбрасывают частицы грунта на лету в радиусе до 2 м от гнезда. В каждом гнезде запасается до 6 экземпляров гусениц (Lepidoptera).
Описание: Самка. Наличник слабо равномерно выпуклый или почти плоский, впереди с узкой широко дугообразно выступающей каймой, ограниченной по бокам небольшими зубцами (рис. 81а). Лицо с густым серебристым опушением и желтовато-серыми длинными отстоящими волосками. Темя за глазами бугровидно выпуклое. Глаза почти параллельные (рис. 81б). Промежуток между задними глазками примерно в 1,3 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка с 6-7 грубыми поперечными морщинами, слегка удлиненная, но ее ширина в середине лишь в 1,5 раза больше длины. Воротничок переднеспинки (рис. 81в) обычно на передней наклонной поверхности уплощенно-вдавленный в середине. Среднеспинка с глубокой срединной бороздой, сильно выпуклой (особенно сзади)средней частью, с поперечными грубыми морщинами (складками) в боковых широких продольных углублениях, с редкими некрупными точками и редкой микропунктировкой на выпуклых участках. Выпуклые участки среднеспинки между срединной бороздой и боковыми углублениями гладкие и блестящие, остальная часть смреднеспинки с более или менее развитым прилегающим опушением. Вогнутая часть среднегруди впереди ограниченная по бокам косыми килями, оканчивающимися внизу угловатым выступом. Прилегающее опушение груди хорошо развитое, особенно на боковых и нижней поверхностях. Щитик и заднещитик с довольно грубыми продольными морщинами. Срединное поле промежуточного сегмента грубо неправильно морщинистое и густо опушенное, лишь по бокам с голыми участками, имеющими более правильную морщинистую скульптуру. Передние тазики с уплощенным угловатым выступом на внутренне-заднем крае. Выступ на вершине 1-го членика передних лапок с 1 мощным темным шипом и 2 более тонкими и более длинными щетинками на вершине (рис. 81г). Голова и грудь черные. Мандибулы на большей части, передняя часть наличника, скапус, тегулы, плечевые бугры сзади и брюшко на большей части красно-рыжие. 1-й членик сте- белька черный. 5-й и 6-й сегменты брюшка черные или бурые. Ноги на большей части красно-рыжие. Тазики, задние вертлуги, задние бедра и задние голени отчасти черные (или бурые). Крылья со слабым, едва заметным равномерным коричневатым затемнением на всей поверхности. Жилки коричневые, у основания крыльев рыжеватые. Длина тела 19-26 мм.
Самец автору не известен. Не исключено, что описанная выше самка является самкой A. elongata Fisch.-Waldh.
Ареал: Юго-Восточная Европа, Иран, Средняя Азия, Казахстан.
Распространение в Казахстане: Пуст. Кызылкум – 5 км вост. и 30 км южн. Чардары; присырдарьинские пустыни – Джулек (близ Чиили); предгорья хр. Каратау – 30 км сев.-вост. Сузака; пуст. Бетпакдала – 5055 км сев.-зап. Фурмановки; Приилийские пустыни – Капчагай, 17-25 км сев.-зап. Баканаса, 10-20 км сев.-вост. Аяк-Калкана; Казахский мелкосопочник – 54 км сев.-вост. Кайнара, 20 км сев. Караула, 12 км сев. Маинкаина, 40 км юго-зап. Аркалыка, 10 км вост. пос. Амангельды, 10 км зап. пос. Ашутасты; Тургайская ложбина – 10 км юго-зап. и 25 км зап. пос. Шийли, Наурзум, Иргиз; Южное Предуралье – 5 км южн. Актюбинска, Бабатай (близ Уила); Северо-Казахская равнина – 20 км юго-зап. Павлодара; Прикаспийская низменность – Рын-пески; Сев. Приаралье – пески Малые Барсуки; долина р. Урал – Индерборский; долина р. Иртыша – Ямышево, Белогорье; Зайсанская котловина – Рожково, 10 км сев.-вост. и 20 км вост. Каратала, 26 км юго-вост. Приозерного, 45 км южн. Каратогая, 72 км юго-вост. пос. Кокпекты.
Экология и биология: Пустынно-степной вид. В степной зоне встречается на песках. Биология не изучена. Самки, вероятно, устраивают в земле одноячейковые гнезда и запасают в них гусениц бабочек (Lepidoptera). Отмечено питание имаго на цветках кермека, зизифоры, крестовника, качима, липучки, молочая, донника, осота, вероники и тамариска.
Синонимы: arvensis Dahlbom, 1843 (Miscus); susterai Snoflak, 1943; alpicola Beaumont, 1945; adriaansei Wilcke, 1945.
Описание: Самка. Наличник слабо выпуклый, с узкой каймой, с почти прямым (или слегка выемчатым) передним краем (между боковыми тупыми углами – рис. 81д), с редкой некрупной пунктировкой и густой микропунктировкой, почти без прилегающих волосков, с чернобурыми отстоящими волосками. Лицо широкое. Глаза слегка сближенные внизу (рис. 81е). Промежуток между задними глазками в 1,6 раза уже промежутка между одним из них и ближайшим глазом. Воротничок переднеспинки вверху в середине с продольным вдавлением. Переднеспинка (рис. 81ж) без морщин, с редкой некрупной пунктировкой и сетчатой микроскульптурой. Среднеспинка с редкой некрупной пунктировкой, сетчатой микроскульптурой и очень нежными поперечными морщинами (особенно отчетливыми в боковых углублениях). Срединная борозда хорошо развитая, суженная кзади, тянется до середины среднеспинки. Щитик с хорошо развитыми продольными морщинами и отдельными точками, блестящий. Заднещитик с неправильными (преимущественно поперечными) морщинами, нечеткими крупными точками и неправильной микроскульптурой, полуматовый. Бока среднегруди с небольшим пятном прилегающего серебристого опушения. Срединное поле промежуточного сегмента с поперечно-косыми нежными морщинами, почти сглаженное сзади, с сетчатой микроскульптурой, полуматовое. Задняя поверхность промежуточного сегмента со сгущенным серебристым опушением. Среднегрудь снизу впереди без бугра, килей и выступов. Передние тазики без зубца. Выступ 1-го членика передних лапок с коротким проксимальным шипом и 2 более длинными и тонкими щетинками на вершине. 3-я субмаргинальная ячейка стебельчатая (рис. 81з). Среднегрудь снизу впереди без бугров, зубцов и килей. Голова, грудь, мандибулы, усики, ноги, стебелек и вершинная часть брюшка черные. Вершина стебелька снизу, 1-й тергум, 2-й сегмент и основание 3-го сегмента брюшка красно-рыжие. Отстоящие волоски головы и переднегруди снизу черно-бурые, остальной части груди светло-серые. Крылья с буроватым затемнением, с бурыми и коричневыми жилками. 2-й тергум перед вершиной с очень узкой желтой поперечной угловатоизогнутой линией. Вершина брюшка без металлического блеска. Длина тела 13-21 мм.
Самец. Передний край наличника в средней части слегка широко выемчатый (рис. 81и). Глаза сильно сближаются книзу (рис. 81к). Минимальное расстояние между глазами примерно в 2 раза меньше расстояния между ними перед передним глазком. Промежуток между задними глазками примерно в 1,5 раза уже промежутка между одним из них и ближайшим глазом. Среднеспинка с довольно редкими некрупными не очень четко ограниченными точками, с нежными неправильными, слабо выраженными косыми морщинами, неправильной микроскульптурой и слабо заметной микропунктировкой. Щитик с неправильными продольными морщинами. Заднещитик с неправильными продольными и поперечными морщинами. Срединное поле промежуточного сегмента нежно поперечно-косо морщинистое, с густой сетчатой микроскульптурой на всей поверхности, полуматовое, голое. Лицо с серебристым опушением. Голова с длинными бурыми волосками, иногда с примесью светло-серых. Грудь с длинными светло-серыми волосками. Бока среднегруди со сгущенным прилегающим опушением в задней части перед метаплеврами, но без четко оформленного «войлочного» пятна. 1-й членик стебелька по длине примерно равен 1-му членику задних лапок и 2/3 2-го. Голова, грудь, ноги и брюшко на большей части черные. Вершина 1-го членика стебелька, 1-й и 2-й тергумы по бокам, основание 3-го тергума, вершина 1-го стернума, 2-й, 3-й и основание 4-го стернума краснорыжие. 1-й и 2-й тергумы сверху на большей части черные. Крылья слегка затемненные. Длина тела 12-18 мм.
Ареал: Северная и Средняя Европа, Кавказ, Казахстан, Монголия
(?).
Распространение в Казахстане: Казахский мелкосопочник – 8 км юго-зап. Каркаралинска; хр. Тарбагатай – 80 км южн. Аксуата; Кокчетавская возвышенность – окр. Щучинска; низкогорья хр. Саур – р. Кендырлик, окр. с. Пржевальского.
Экология и биология: Вид распространен в лесо-степной и лесной ландшафтных зонах, южнее встречается в горах. В лесах придерживается открытых участков. Самки устраивают одноячейковые гнезда в земле, на солнечных местах, свободных от растений, с не слишком рыхлой песчаной почвой. Добыча – голые гусеницы бабочек (Lepidoptera: Geometridae, Noctuidae), очень редко личинки пилильщиков (Hymenoptera: Tenthredinidae). В каждой ячейке запасается от 3 до 10 экземпляров добычи. Для вида отмечено прогрессивное провиантирование, т.е. снабжение потомства пищей в течение нескольких дней (а не сразу в один день), причем одновременно в нескольких гнездах.
Синонимы: hortensis Poda, 1761 (Sphex); frischii Fourcroy, 1785 (Ichneumon); dimidiata Christ, 1791 (Sphex), nec De Geer, 1773; vulgaris W.Kirby, 1798; pulvillata Sowerby, 1805; mucronata Jurine, 1807 (Sphex); cyanescens Dahlbom, 1845; ? vischu Cameron, 1889; marginalis Perez, 1905; oraniensis Roth, 1928.
Подвиды: vagabunda F.Smith, 1856 (Китай, Тайвань); infesta F.Smith, 1873 (Япония, Корея); tonareg Ed.Andre, 1886 (Алжир); kamtschatica Gussakovskij, 1933 (п-ов Камчатка); nipponica Tsuneki, 1967 (Япония, Китай, Южная и Северная Корея).
Описание: Самка. Наличник равномерно выпуклый, с узкой каймой впереди, ограниченной по бокам зубцевидными углами (рис. 82а). Внутренние края глаз почти параллельные (рис. 82б). Наличник с редкими точками и микропунктировкой. Опушение очень слабо развитое (в верхней части). Промежуток между задними глазками почти в 1,9 раза уже промежутка между одним из них и ближайшим глазом. Воротничок (валик) переднеспинки примерно в 2 раза шире своей длины (рис. 82в), без продольной выемки в середине. Среднеспинка с неравномерной некрупной пунктировкой. Точки разной величины. Поперечные морщины почти отсутствуют, лишь по бокам нечетко обозначенные. Щитик с продольными морщинами, впереди более или менее сглаженный, с несколькими точками. Срединное поле промежуточного сегмента с грубой неправильной морщинистой скульптурой и неправильной микроскульптурой, с длинными светлыми волосками на всей поверхности. Грудь с пятнами густого серебристо-белого прилегающего опушения на плечевых буграх, боках среднегруди и боках промежуточного сегмента у вершины. 1-й членик стебелька брюшка почти равен 1-му членику задних лапок и половине 2-го вместе. 3-я субмаргинальная ячейка не стебельчатая. Среднегрудь снизу впереди без бугров, зубцов и килей. Голова, грудь, усики и ноги черные. 1-3-й тергумы красные. Основание 1-го тергума черное. Крылья с буроватым затемнением, с черно-бурыми и коричневыми жилками. Длина тела 20-28 мм.
Самец. Передний край срединной лопасти наличника почти прямой или слегка выемчатый (рис. 82г). Наличник с густым прилегающим опушением и длинными светлыми волосками. Глаза сильно сближаются книзу (рис. 82д). Промежуток между ними у переднего глазка примерно в 1,5 раза шире минимального на наличнике. Переднеспинка более чем в 2 раза короче своей ширины в середине, с продольным вдавлением в середине (рис. 82е). Среднеспинка с неравномерной довольно крупной (в средней части редкой) пунктировкой и густой микропунктировкой, по крайней мере, на вогнутых боковых участках с поперечными морщинами. Форма маргинальной и субмаргинальных ячеек как на рис. 82ж. Скульптура, опушение, волосяной покров почти как у самки. 1-й членик стебелька по длине равен 1-му членику задних лапок и 2/3 2-го. 3-й тергум на заднем крае черный. 1-й тергум часто с черной продольной полосой сверху. 2-й тергум иногда тоже с черным продольным удлиненным пятном. Вершина брюшка с синеватым металлическим блеском. Голени и лапки более или менее буроватые. Длина тела 17-23 мм.
Ареал: Почти вся Европа, Северная Африка, Юго-Западная Азия, Средняя Азия, Казахстан, Сибирь, Дальний Восток, Монголия, Китай, пов Корея, Япония, Тайвань.
Распространение в Казахстане: Долина р. Сырдарьи и прилегающие пустыни – Джулек (близ Чиили), Байгакум (близ Джулека), КзылОрда, Джетысай; долина р. Чу – Фурмановка; низкогорья и предгорья Западного и Северного Тянь-Шаня – 15 км юго-вост. и 40 км южн. Ленгера, 20-25 км южн. Мерке, р. Чарын (ур. Сортогай), Талгар, 8-10 км южн. Талгара, хр. Чулак (Шолак), Кзыл-Агач, Жаркент (Панфилов), Подгорное (хр. Кетмень), Кегеньский перевал (хр. Кетмень), Бартогай (р. Чилик), 25 км южн. пос. Тургень, Алматы, 5-15 км южн. и 10-20 км зап. Алматы; долина р. Или и прилегающие пустыни – 17 км сев.-зап. и 65 км зап. Баканаса, 35 км сев.-зап. Капчагая, Или (бывш.), устье р. Каскелен, ур. Мын-Булак (близ Аяк-Калкана), 15 км вост. Аяк-Калкана, 5 км юго-вост. и 8-10 км южн. Жаркента (бывш. Панфилов); Вост. Прибалхашье – Карачаган; Казахский мелкосопочник – Жарма, Каркаралинск, Ставрополка, 8 км юго-зап. Каркаралинска, 15-20 км южн. Караула, 5 км сев. Державинска; Тургайская ложбина – Наурзум; Южное Предуралье –
Уил; долина р. Иртыша и прилегающая равнина – Белогорье, Лебяжье, Семипалатинск, 7 км сев. Семипалатинска, Канонерка; предгорья хр. Саур и хр. Тарабагатай – Таскескен, Пржевальское, 35 км вост. Пржевальского, 5 км. сев. Ириновки; предгорья и низкогорья Южного Алтая – Курчум, 20 км юго-зап. Ленинки, 12 и 70 км юго-вост. Георгиевки, Чердояк, 25 км зап. Катон-Карагая, Урыль, Берель, 11-27 км вост. Большенарымского, 15 км сев. Алексеевки, 40 км сев.-вост. пос. Кокпекты, Усть-Каменогорск, 45-60 км южнее, 10 км сев.-зап. и 15 км сев.-вост. Усть-Каменогорска, 8-15 км сев. Самарского, Раздольное, Северное, 15 км юго-зап. Зыряновска, Серебрянск, Горная Ульбинка, Палаты; Кокчетавская возвышенность – Боровое, 40 км юго-вост. Кокчетава, 15 км южнее Степняка.; Северо-Казахская равнина – 10 и 150-180 км вост. Кустаная.
Экология и биология: Эврибионтный вид. Встречается в горах и на равнинах от лесной зоны до пустынной. Избегает песков. В пустынной зоне предпочитает долины рек, оазисы и горные ущелья; в лесной зоне – открытые участки. Самки устраивают одноячейковые гнезда в норках, которые выкапывают сами в уплотненном грунте. Добыча – голые гусеницы бабочек (Lepidoptera: Noctuidae, Geometridae, Limantriidae, Pieridae, Notodontidae и др.). Строительство гнезда предшествует охоте. В ячейке запасается 1-2 (редко больше) экземпляра добычи. Отмечено питание взрослых ос на цветках молочая, липучки, дягиля, ферулы, морковника, лука, синеголовника, кермека, флокса, тамариска,, ластовня, клоповника, барбариса, мирикарии и саусуреи. A. sabulosa – один из немногих видов рода Ammophila, которые встречаются в населенных пунктах и других антропогенных биотопах.
Описание: Самка. Наличник бугровидно выпуклый, на передней поверхности бугра более или менее сглаженный, с легкой пунктировкой и нежными продольными неправильными морщинами, в середине слегка вдавленный, впереди с каймой, узко выемчатой в середине и ограниченной по бокам острыми зубцами (рис. 82з). Верхняя часть наличника с густым золотистым прилегающим опушением. Лицо почти на всей поверхности с густым золотистым прилегающим опушением. . Передний край срединной лопасти наличника. Глаза почти параллельные (рис. 82и), лицо на лбу у переднего глазка слегка шире, чем на наличнике. Промежуток между задними глазками примерно в 1,3 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка не удлиненная (рис. 82к), слегка продольно вдавленная вверху в середине. Ширина воротничка в середине почти в 2 раза больше его длины. Среднеспинка с поперечными и косыми, не очень грубыми морщинами на всей поверхности. Переднеспинка и передняя половина среднеспинки с густым прилегающим золотистым (или латунным) бленстящим опушением. Задняя половина среднеспинки с менее густым опушением; волоски полуприлегающие. Щитик и заднещитик с продольными морщинами. Срединное поле промежуточного сегмента голое, с косыми морщинами, которые в средней части неправильно изломанные. Низ среднегруди со срединной продольной линией, но без бугров, зубцов и килей в передней части. Плечевые бугры, бока среднегруди в средней части, бока промежуточного сегмента на большей части с густым прилегающим латунно-блестящим опушением. 1-й членик стебелька брюшка по длине равен 1-му членику задней лапки и половине 2-го вместе. Голова, грудь, ноги и усики черные. 1-4-й сегменты брюшка (кроме верхней части 4-го тергума) рыжевато-красные (включая стебелек). Вершина брюшка со слабым металлическим блеском. Крылья со слабым коричневым затемнением, с бурыми и коричневыми (отчасти рыжеватыми) жилками. Тегулы красно-рыжие. Лапки буроватые. Длина тела 18-21 мм.
Самец. Наличник в середине сильно бугровидно выпуклый, с выемчатым передним краем (рис. 82л,м). Мандибулы у основания спереди с выступающей складкой (рис. 82н), в средней части сильно расширенные. Опушение лица густое, скрывающее скульптуру. Глаза сильно сближаются книзу. Ширина лица перед передним глазком приблизительно в 1,45 раза больше минимальной ширины его на наличнике. Промежуток между задними глазками почти равен по ширине промежутку между одним из них и ближайшим глазом или слегка уже. Воротничок переднеспинки примерно вдвое шире своей длины, с густым прилегающим опушением. Среднеспинка с четкими поперечными (отчасти косыми) морщинами, с прилегающим опушением. Щитик и заднещитик с грубыми продольными морщинами. Срединное поле промежуточного сегмента с поперечно-косыми морщинами (в средней части более или менее изломанными), с четкой сетчатой микроскульптурой, полуматовое. 3-я субмаргинальная ячейка не стебельчатая. Среднегрудь впереди без килей, зубцов и выступов. Передние тазики без выступа сзади. Бока груди с хорошо развитым прилегающим опушением, особенно густым на плечевых буграх, средней части боков среднегруди, задней части промежуточного сегмента и верхней части задних тазиков. Голова, грудь, усики и ноги черные. Скапус с небольшим рыжим пятном. Тегулы красно-рыжие. Крылья коричневые. Жилки бурые и коричневые. Брюшко почти целиком или на большей части красно-рыжее. У наиболее темных экземпляров 1-й членик стебелька и 2-3 вершинных сегмента брюшка черные. Лапки буроватые. Прилегающее опушение головы и груди серое, с золотистым или латунным блеском. Длина тела 16-22 мм.
Ареал: Юг Восточной Европы, Малая Азия, Казахстан.
Распространение в Казахстане: Предгорья хр. Каратау – 18 км сев.зап. Кантемировки; предгорья и низкогорья Джунгарского Алатау – хр. Архарлы, хр. Малайсары, Кзыл-Агач, 12 км сев. Сары-Озека; предгорья Заилийского Алатау – 10 км сев.-зап. Чемолгана.
Экология и биология: Пустынно-сухостепной вид. Биология не изучена. Самки, вероятно, устраивают одноячейковые гнезда в неглубоких норках, которые выкапывают в уплотненном грунте, и запасают в них гусениц бабочек (Lepidoptera). Отмечено питание имаго на цветках кермека, зизифоры и цельнолистника. Редкий вид.
Описание: Самка. Наличник c узкой гладкой каймой, имеющей по бокам угловатые выступы (рис. 83а). Срединная лопасть наличника выпуклая, в середине уплощенная. Передний край срединной лопасти наличника слегка тупоугольно выемчатый в середине. Наличник на большей части (кроме голой широкотреугольной зоны впереди) и нижняя часть лба с густым прилегающим опушением, скрывающим скульптуру, и длинными отстоящими светло-коричневыми волосками. Налобник вдавленный. Глаза почти параллельные(рис. 83а). Промежуток между задними глазками примерно в 1,7 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка с не очень грубыми поперечными морщинами(рис. 83б) и с редкими точками. Воротничок примерно в 1,5 раза короче своей ширины в середине, спереди без резкого уступа (рис. 83в), в середине слегка уплощенно вдавленный. Переднеспинка отделена от среднеспинки глубокой, относительно широкой перетяжкой. Среднеспинка с глубокой срединной бороздой, сильно выпуклыми блестящими продольными участками по бокам от борозды, с 2 сильно вдавленными продольными участками в задней половине (в середине боковых поверхностей с резкими, но не очень грубыми косыми морщинами, редкими точками и неправильной микроскульптурой почти на всей поверхности). Опушение слабо развитое, очень короткое. Эпистернальный шов вверху в виде поперечной складчатой борозды, ниже уровня скробальной ямки слабо развитый, едва заметный. Низ среднегруди впереди вогнутый, с изогнутыми боковыми килями, оканчивающимися снизу мощным зубцом. Передние тазики с угловатым выступом на задневнутреннем крае. 1-й членик передних лапок с 7 крепкими светлокоричневыми щетинками на наружном крае, на вершине выступа с коротким изогнутым черным шипом и 2 крепкими длинными бурыми щетинками. Щитик с продольной, заднещитик с неправильной морщинистой скульптурой. 3-я субмаргинальная ячейка не стебельчатая (рис. 83г). Срединное поле промежуточного сегмента с четко выраженной срединной линией и косыми морщинами, по бокам голое, в середине с очень нежным коротким полуприлегающим светлым опушением. 1-й членик стебелька брюшка слегка короче 1-го и 2-го члеников задних лапок. Голова, грудь, усики, ноги и мандибулы черные. Тегулы сзади буровато-рыжие. 1-й тергум, 2-й и 3-й сегменты красно-рыжие. 1-й тергум у основания черный. Вершина брюшка черная, со слабым синим блеском. Крылья слегка буроватые. Жилки коричневые, у основания крыльев рыжеватые. Длина тела 17-21мм.
Самец. Наличник слабо вытянутый, почти плоский, в передней части в середине слегка уплощенно-вдавленный, слабо выемчатый на переднем крае между боковыми углами (рис. 83д). Лицо с серебристым прилегающим опушением. Глаза сильно сближаются книзу (рис. 83д). Промежуток между задними глазками примерно в 1,4 раза уже промежутка между одним из них и ближайшим глазом. Верх лба с явственной редкой пунктировкой и неправильной микроскульптурой. Переднеспинка удлиненная, с поперечными морщинами, впереди с уступом (рис. 83е). Среднеспинка с глубокой срединной бороздой, с довольно грубыми поперечными морщинами, с нечетко ограниченными крупными точками. Щитик выпуклый, с неправильно изогнутыми, преимущественно продольными довольно грубыми морщинами. Заднещитик примерно с такой же скульптурой. Срединное поле промежуточного сегмента с морщинистой скульптурой, со срединной килеобразной складкой (морщиной). Морщины в средней части поля неправильно изогнутые, по бокам почти правильные, косые; опушение слабо развитое, не скрывающее скульптуру, не прилегающее, по бокам поля отсутствует. Низ среднегруди впереди вогнутый, с 2 косыми килями, оканчивающимися острым выступом. Передние тазики с остроугольным выступом на задневнутреннем крае. 1-й членик стебелька брюшка плавно изогнутый (см. сбоку), по длине примерно равен 1-му и 2-му членикам задних лапок. Голова, грудь, усики и ноги черные. 1-й тергум, 2-й сегмент целиком и 3-й сегмент почти целиком красно-рыжие. 1-й членик стебелька на вершине красно-рыжий, 2-й у основания с черным пятном. Крылья слабо затемненные. Жилки буровато-коричневые, у основания крыльев рыжеватые. Щупики бурые. Длинные волоски головы и груди светло-серые. Длина тела 14-18 мм.
Ареал: Юго-Восточная Европа (Предкавказье), Казахстан, Южная Сибирь, Китай.
Распространение в Казахстане: Прииртышье – Семипалатинск, Павлодар, Федоровка; Зайсанская котловина – 72 км юго-вост. пос. Кокпекты, 10 км сев.-вост. Каратала; низкогорья Алтая.
Экология и биология: Степной вид. Биология неизвестна. Вероятно, самки устраивают одноячейковые гнезда в норках в земле и запасают в них гусениц бабочек (Lepidoptera).
Подвид: nadigi Roth, 1932 – Сев. Африка.
Описание: Самка. Наличник равномерно выпуклый, впереди уплощенный или слегка вдавленный, с 2 боковыми зубцами на переднем крае, между зубцами слегка дугообразно выступающий или почти прямой (рис. 83ж). Опушение лица очень короткое, слабо развитое, не скрывающее скульптуру. Лицо с мелкой неравномерной, в целом редкой пунктировкой, густой микропунктировкой и очень нежной сетчатой микроскульптурой. Передняя зона наличника голая в середине. Лицо широкое. Глаза почти параллельные (рис. 83з). Лобная борозда глубокая, лоб в целом не вдавленный. Промежуток между задними глазками примерно в 1,4 раза уже промежутка между одним из них и ближайшим глазом. Переднеспинка сверху с коротким продольным вдавлением в середине. Воротничок (рис. 83и) более чем в 2 раза шире своей длины в середине, с довольно грубыми поперечными морщинами, без резкого уступа впереди (рис. 83к). Среднеспинка в середине сильно выпуклая, гладкая и блестящая, с редкими некрупными точками, с глубокой поперечно морщинистой и пунктированной срединной бороздой, с глубокими боковыми вдавлениями (тянущимися вперед от заднего края до 2/3 длины среднеспинки), с грубыми поперечными морщинами по бокам впереди и во вдавлениях. Эпистернальный шов развитый. Щитик с продольными морщинами, в середине слегка продольно вдавленный; заднещитик с неправильной морщинистой скульптурой. Срединное поле промежуточного сегмента с косыми (в середине неправильно изогнутыми) морщинами, по бокам голое, в середине с очень нежным полуприлегающим опушением. Низ среднегруди впереди без бугров, килей и зубцов. 1-й членик передних лапок с 5 крепкими щетинками на наружном крае и 3 щетинками на вершинном выступе (рис. 84а). 3-я субмаргинальная ячейка не стебельчатая (рис. 84б). Голова, грудь, ноги черные. Брюшко с красно-рыжим основанием. Длина тела 22-25 мм.
Самец. Наличник относительно широкий, не вытянутый, со слегка выемчатым передним краем (рис. 84в,г), впереди в середине слегка уплощенно-вдавленный, с длинными отстоящими бурыми волосками и коротким серым прилегающим опушением. Глаза сильно сближаются книзу (рис. 84г). Переднеспинка и среднеспинка с редкими поперечными, щитик и заднещитик с продольными морщинами. Скульптура срединного поля промежуточного сегмента как у самки. 1-й членик стебелька по длине почти равен 1-му и 2-му членикам задней лапки вместе. Голова и грудь черные. 1-й тергум и 2-й сверху черные (или бурые), по бокам красно-рыжие. 3-й тергум черный, только по бокам частично краснорыжий. 3-й стернум красно-рыжий. Опушение тела слабо развитое, грудь без пятен густого прилегающего опушения. Длина тела 19,5-23 мм.
Ареал: Северная Африка, Юго-Западная Азия, Казахстан, Монголия, Восточная Сибирь, Приморье.
Распространение в Казахстане: Казахский мелкосопочник – горы Улытау; 5 км южнее и 40 км западнее Аягуза, 20-25 км сев.-зап. Тансыка; Алтай – 29 км сев.-сев.-вост. Курчума.
Экология и биология: Вид предпочитает открытые степные участки с щебнисто-глинистой почвой. Биология не изучена. Самки, вероятно, устраивают одноячейковые гнезда в земле и охотятся на гусениц бабочек (Lepidoptera). Отмечено питание ос на цветках молочая и кермека.
Описание: Самка. Мандибулы с маленьким зубчиком у основания вершинного зубца на внутреннем крае. Передний край наличника с 2 неглубокими выемками, ограничивающими срединную лопасть (рис. 84д). Наличник довольно сильно выпуклый (равномерно буровидный в средней части). Глаза слегка расходятся книзу. Промежуток между задними глазками примерно в 1,4 раза уже промежутка между одним из них и ближайшим глазом. Темя позади глазков бугровидно выпуклое. Переднеспинка удлиненная, со слабо развитыми поперечными морщинами (рис. 84е). Среднегрудь снизу впереди со слабо заметными боковыми косыми килями, оканчивающимися внизу бугорком. Передние тазики с угловатым выступом на задне-внутреннем крае. Выступ на вершине 1-го членика передних лапок на конце с 3 крепкими длинными, почти одинаковыми щетинками (рис. 84ж). Апикальные выступы члеников передних лапок сильно удлиненные (рис. 84ж). Переднеспинка с уступом впереди. Срединное поле промежуточного сегмента с густым прилегающим опушением на всей поверхности, скрывающим скульптуру, которая состоит преимущественно из неправильно изогнутых коротких продольных морщин и неправильной микроскульптуры. 1-й членик стебелька брюшка по длине примерно равен 1-му и 2-му членикам задней лапки. Среднегрудь снизу впереди без бугров, зубцов и килей. Голова, грудь, тазики и вертлуги ног с густым прилегающим серебристым опушением. Мандибулы на большей части, наличник почти целиком, скапус, педицеллюс, плечевые бугры, тегулы, вершина промежуточного сегмента, брюшко и ноги почти целиком рыжие или красно-рыжие. Жгутик усиков бурый, у основания рыжеватый. Крылья не затемненные, с рыжими и коричневыми жилками. Коста, субкоста, птеростигма и основание главных продольных жилок рыжие. Задние тазики, задние вертлуги и задние бедра сверху с черными или бурыми пятнами. Длина тела 16-20 мм.
Самец. Наличник плоский, с суженной вытянутой вершиной, впереди в середине с закругленной выемкой. Воротничок переднеспинки по длине примерно равен своей ширине, сверху с поперечными морщинами. Скульптура, окраска и опушение тела почти как у самки. Задние бедра сверху и задние лапки на вершине черные. Стебелек брюшка сверху с черной продольной полосой. 5-7-й сегменты брюшка всегда черные. Скапусы впереди с черным пятном. Длина тела 15-18 мм.
Ареал: Иран, Средняя Азия, Южный Казахстан.
Распространение в Казахстане: Пуст. Кызылкум – 5-8 км юго-зап. Чардары; Приилийские пески – 3 км юго-вост. Капчагая, 8 км южн. Жаркента (бывш. Панфилов).
Экология и биология: Обитатель песчаных пустынь. Биология не изучена. Самки, вероятно, устраивают одноячейковые гнезда в земле и охотятся на гусениц бабочек (Lepidoptera).
Синонимы: apicalis Brulle, 1876, nec Guerin-Meneville, 1835; minuta Frivaldsky, 1876; rhaetica Kohl, 1879; julii Fabre, 1879; moksari Marquet, 1881; ? kirgisica F.Morawitz, 1890; turkestana Kohl, 1906, nec Dalla Torre, 1897.
Подвиды: mocsaryi Frivaldsky, 1876 – Южная Европа, Западная Азия; electa Kohl, 1901 – Сев. Африка; turkestana Kohl, 1906 – Средняя Азия. В Казахстане, помимо типичного подвида, обитают еще два. Описания всех трех подвидов приводятся ниже.
Описание: Самка. Наличник с узкой каймой впереди, ограниченной по бокам слабо выступающими углами (рис. 84з). Срединная лопасть наличника слабо равномерно выпуклая, с редкими точками разной величины. Лицо широкое (рис. 84з). Глаза явно сближенные внизу. Промежуток между задними глазками примерно в 1,6 раза уже промежутка между одним из них и ближайшим глазом. Опушение лица слабо развитое, не скрывающее скульптуру. Воротничок переднеспинки короткий (рис. 84и), по длине примерно в 2 раза шире своей длины. Переднеспинка и среднеспинка с резкими поперечными морщинами. Срединная борозда среднеспинки узко-клиновидная, неглубокая. Щитик и заднещитик с густыми продольными морщинами. Заднещитик бугровидно выпуклый. Сединное поле промежуточного сегмента голое, в середине изломанно-морщинистое, по бокам косо-морщинистое и более блестящее, чем в середине. Среднегрудь снизу впереди без бугров, зубцов и килей. 3-я субмаргинальная ячейка не стебельчатая (рис. 84к). Передние тазики без выступов на верхне-внутренней поверхности. Опушение тела слабо развитое. Бока среднегруди сзади, задне-боковая часть промежуточного сегмента и задние тазики сверху с густым серебристым прилегающим опушением.Голова, грудь и усики черные. Нижнегубные щупики бурые. Мандибулы черно-бурые. Тегулы бурые, иногда рыжеватые сзади. Крылья с буроватым затемнением, с бурыми и коричневыми жилками. 1-й членик стебелька черно-бурый; 2-й, 3-й и передняя часть 4-го красно-рыжие. Вершинные членики брюшка с синеватым металлическим оттенком. Ноги черные, отчасти буроватые. Длина тела 12-21 мм.
Самец. Передний край наличника в середине слегка широко тупоугольно выемчатый или прямой (рис. 85а). Глаза сильно сближаются книзу (рис. 86б). Максимальная ширина лица примерно в 2 раза больше минимальной. Лицо на большей части с густым прилегающим серебристым опушением. Промежуток между задними глазками примерно в 1,5 раза уже промежутка между одним из них и ближайшим глазом. Признаки строения груди, окраска, скульптура и опушение тела почти как у самки. 1-й членик стебелька брюшка черный; 1-й, 2-й и 3-й сегменты и основание 4-го тергума красно-рыжие. 3-й тергум обычно с черным поперечным пятном сзади. Ноги черные. Крылья слабее затемненные, чем у самки. Длина тела 10-14 мм.
Ammophila terminata turanica Kazenas, subsp.n.
Голотип: ♀. Долина р. Или, окр. Аяк-Калкана (35 км сев.-вост. Чилика), 14 VI 1971 (Казенас). Хранится в Зоологическом институте РАН (г. Санкт-Петербург).
Описание: Самка. Лицо на большей части с густым прилегающим серебристым опушением, скрывающим скульптуру, и длинными отстоящими белыми волосками. Глаза слегка сближаются книзу (рис. 85в). Переднеспинка в середине слегка вдавленная, со слабо заметными поперечными морщинами. Среднеспинка с менее резко выраженными, чем у типового подвида, поперечными морщинами. Щитик и заднещитик с менее развитыми морщинами, чем у типового подвида. Опушение тела сильнее развитое, на боках груди почти везде скрывающее скульптуру. Срединное поле промежуточного сегмента почти как у типового подвида, только морщины гуще и нежнее. 1-й членик передних лапок как на рис. 85г. 1-й членик стебелька брюшка красн0-рыжий. 4-й тергум почти целиком красно-рыжий, с грязно-бурыми пятнами. Длина тела 20-22 мм.
Самец. Глаза сильнее развиты, чем у типового подвида. Передний край наличника слегка выемчатый (рис. 85д). Переднеспинка (рис. 85е) в середине с продольным вдавлением. Срединное поле промежуточного сегмента с очень нежной сетчатой микроскульптурой (между косыми морщинами). 1-й членик стебелька только в вершинной части краснорыжий. Вершина брюшка с зеленовато-синим металлическим блеском. Ноги с ясным нежным прилегающим опушением. Голова и грудь с длинными белыми волосками. 4-й тергум с красно-рыжим основанием. Длина тела 17-19 мм.
Описание: Самка. Передний край срединной лопасти наличника с 2 боковыми угловатыми выступами, между ними с 2 слабо развитыми дугообразными выступами, с выемкой в середине. Наличник с сетчатой микроскульптурой, с редкими точками разной величины, почти без опушения. Лицо широкое, глаза слегка сближаются книзу. Промежуток между задними глазками примерно в 1,9 раза уже промежутка между одним из них и ближайшим глазом. 1-й членик передних лапок как на рис. 85ж. Переднеспинка (рис. 85з) и среднеспинка с поперечными (на среднеспинке более или менее скошенными, особенно сзади) густыми, не очень грубыми морщинами и редкими, нечетко ограниченными точками. Щитик довольно нежно продольно морщинистый, заднещитик с неправильной ячеисто-морщинистой скульптурой. Срединное поле промежуточного сегмента нежно косо морщинистое, без прилегающего опушения, только с редкими нежными отстоящими волосками. Бока груди без пятен прилегающего светлого опушения. Форма маргинальной ячейки как на рис. 85и. 1-й членик стебелька брюшка короче, чем 1-й и 2-й членики задних лапок, взятые вместе. Среднегрудь снизу впереди без бугров, зубцов и килей. Голова, грудь, усики и ноги черные. Отстоящие волоски головы и груди бурые и черные. Стебелек брюшка, 1-й тергум, 2-й, 3-й и основание 4-го сегмента брюшка красно-рыжие. Вершина брюшка черная, с металлическим синим оттенком. Крылья с буроватым затемнением, с бурыми жилками. Длина тела 17-19 мм.
Самец. Наличник со слегка выемчатым или почти прямым передним краем (рис. 85к), слабо выпуклый, с редкими мелкими точками и сетчатой микроскульптурой. Глаза сильно сближаются книзу (рис. 85к). Лоб вверху и темя почти матовые из-за густой микроскульптуры. Лоб внизу и наличник со слабо развитым прилегающим светлым опушением. Волоски головы черные. Скульптура груди, как у самки. Тегулы сзади бурые. 1-й членик стебелька почти до вершины черный. 3-й тергум у вершинного края черный. 4-й сегмент лишь у основания рыжий. Длина тела 17-18 мм.
Ареал вида: Южная и Средняя Европа, Северная Африка, ЮгоЗападная Азия, Средняя Азия, Казахстан, Южная Сибирь. .
Распространение в Казахстане: A. t. terminata F.Sm. – предгорья и низкогорья Западного и Северного Тянь-Шаня – Кзыл-Орда, Жамбыл, 25 км зап. Жамбыла, Глинково, Чу, Алматы, Талгар, Тескенсу, Бартогай (р. Чилик), Подгорное (хр. Кетмень), Отар, Малое Алматинское ущ., Кзыл-Агач (предгорья Джунгарского Алатау), хр. Алтынемель, Коныролен; Сев. и Вост. Прибалхашье – 15 км юго-зап., 14 км сев. и 170 км сев. Балхаша, Карачаган, Рыбачье, 25 км юго-зап. пос. Лепсы; Казахский мелкосопочник – повсюду; Тургайская ложбина – Семиозерное, АманКарагай, Шийли (р. Караторгай), 20-30 км сев.-вост. пос. Амангельды, 3 км юго-вост. и 30 км юго-зап. Иргиза; Южное Предуралье – Берчогур, Уральск, Уил, Бабатай (близ Уила), Актюбинск, Володарское; Прикаспийская низменность – Елтай, 15 км сев.-зап. пос. Фурманово, Доссор, Рын-пески; плато Устюрт; п-ов Мангышлак; Северо-Казахстанская равнина – 50 км зап. Павлодара; Прииртышье – 7-40 км сев. Семипалатинска, Лебяжье, Белогорье, Щербакты, 8 км юго-вост. Большой Владимировки; Зайсанская котловина – Акжар, 12 км юго-зап. Бурана, 72 км юговост. пос. Кокпекты, 10 км сев.-вост. Каратала; хр. Саур – 40 км юговост. Зайсана; Алтай – Катон-Карагай, 12 км сев.-вост. Курчума, 10-40 км сев.-зап. Усть-Каменогорска, Калбинский хр.
A. t. turkestana Kohl – пуст. Кызылкум (50 км юго-зап. и 30 км зап. Баиркума, 6-8 км юго-зап. Чардары).
A. t. turanica Kaz. – Приилийские пустыни – 15 км вост. АякКалкана, окр. Баканаса, 17-65 км зап. Баканаса, окр. Капчагая.
Экология и биология: Пустынно-степной вид. В лесостепной
зоне встречается на открытых участках с более или менее рыхлыми песчаными и супесчаными
почвами. В степной зоне обычен повсюду. В пустынной зоне в основном встречается
в ущельях невысоких гор и по долинам рек. Самки роют норки в земле, разбрасывая
частицы почвы на лету вокруг входа в радиусе до 3 м. Устройство гнезда предшествует
охоте. Добыча – гусеницы пядениц (Geometridae). Отмечено питание взрослых ос на
цветках кермека, молочая, зизифоры, синеголовника, тысячелистника, лука, сусака,
вероники, селитрянки, тамариска, клоповника и других растений.
4. ЗАКЛЮЧЕНИЕ
В настоящей работе, которая представляет собой 1-й выпуск монографии «Роющие осы» для 30-томной серии «Фауна Казахстана», рассмотрены только 2 подсемейства (из 8 известных для республики): Ampulicinae и Sphecinae. Для Казахстана зарегистрировано 2 вида Dolichurus (триба Dolichurini, подсем. Ampulicinae), 2 вида Chalybion, 4 вида Sceliphron (триба Sceliphronini), 5 видов Sphex, 5 видов Palmodes и 12 видов Prionyx (триба Sphecini), 1 вид Parapsammophila, 2 вида Eremochares, 9 видов Podalonia и 18 видов Ammophila (триба Ammophilini, подсем. Sphecinae) – всего 60 видов из 10 родов и 4 триб. В следующих выпусках предполагается рассмотреть семейства Astatinae и Pemphredoninae (2-й выпуск), Crabroninae (3-й выпуск), Nyssoninae, Entomosericinae (4-й выпуск) и Philanthinae (5-й выпуск). Общий анализ фауны роющих ос Казахстана будет сделан в конце работы.
ЛИТЕРАТУРА
Антропов А.В. К познанию роющих ос рода Trypoxylon (Hymenoptera, Shecidae) палеарктической фауны // Зоол. журн., 1986. Т. 65, вып. 4. С. 624-628.
Антропов А.В. О таксономическом статусе Trypoxylon attenuatum Smith, 1851 и близких видов роющих ос (Hymenoptera, Sphecidae) // Энтомол. обозр., 1991. Т. 70, вып. 3. С. 672-685.
Антропов А.В. Роющие осы рода Belomicroides (Hymenoptera, Sphecidae) азиатской фауны // // Зоол. журн., 1994. Т. 73, вып. 1. С. 8996.
Будрис Э.Р. Новые и малоизвестные виды рода Mimesa Shuck. (Hymenoptera, Sphecidae) из Средней Азии и Казахстана и определительная таблица видов фауны СССР // Тр. Зоолог. ин-та АН СССР, 1985. Т. 132. С. 58-76.
Будрис Э.Р. Новые и малоизвестные виды роющих ос трибы Psenini (Hymenoptera, Sphecidae) // Тр. Всесоюзн. энтомол. о-ва, 1988. Т. 70. С. 101-116.
Будрис Э.Р. Роющие осы рода Mimumesa Malloch (Hymenoptera,
Sphecidae) фауны СССР // Энтомол. обозр., 1990. Т. 69, вып. 4. С. 941948.
Будрис Э.Р., Казенас В.Л. Новые виды роющих ос рода Diodontus (Hymenoptera, Sphecidae) // Зоол. журн., 1992. Т. 71, вып. 8. С. 24-30.
Гуссаковский В.В. Палеарктические виды рода Astatus Latr. (Hymenoptera, Sphecidae) // Ежегодник Зоол. муз. АН СССР, 1927. Т. 28. С. 265-296.
Гуссаковский В.В. Палеарктические виды рода Solierella Spin. (Hymenoptera, Sphecidae) // Рус. энтомол. обозр., 1928а. Т. 22, N 1-2. С. 78-84.
Гуссаковский В.В. Новые виды Sphecidae из Закаспия и Хивы // Изв. Курсов прикл. зоол. фитопатол., 1928б. Вып. 4. С. 3-19.
Гуссаковский В.В. Новые и малоизвестные виды родов Ammophila Kby. и Sphex L. (Hym. Sphecidae) // Рус. энтомол. обозр., 1930а. Т. 24, N 3-4. С. 199-211.
Гуссаковский В.В. Жалящие перепончатокрылые // В кн.: Труды Памирской экспедиции 1928 г. II. Зоология. Л.: Изд-во АН СССР, 1930б. С. 67-78.
Гуссаковский В.В. Восточнопалеарктические виды рода Gastrosericus Spin. (Hymenoptera,Sphecidae) // Ежегодник Зоол. Муз. АН СССР, 1930в. Т. 31, вып. 3-4. С. 449-457.
Гуссаковский В.В. Поправки и дополнения к ревизии рода Solierella Spin. (Hymenoptera) // Рус. Энтомол. Обозр., 1930 г. Т. 24, N 3-4. С. 232-235.
Гуссаковский В.В. (Gussakovskij V.V. Eine neue Sphecidengattung von Transcaspien // Rev. Espan. Entomol., 1930д. N.6. P. 275-286.
Гуссаковский В.В. (Gussakovskij V.V. Revision der Gattung Ammoplanus Giraud und einigen verwandten Sphegidengattungen // Bol. Soc. esp.
Hist. natur., 1931. Vol. 31. P. 437-465.)
Гуссаковский В.В. Sphecidae и Psammocharidae (Hymenoptera), собранные Н.Зарудным в восточной Персии // Тр. Зоол. Ин-та АН СССР, 1933а (1932). Т. 1. С. 269-308.
Гуссаковский В.В. Обзор видов рода Prosopigastra Costa (s.lat.) (Hym., Sphecidae) // Энтомол. Обозр., 1933. Т. 25, вып. 1-2. С. 154-173.
Гуссаковский В.В. К фауне ос ( Hymenoptera, Sphecoidea et Vespoidea) Таджикистана // Тр. Тадж. базы АН СССР, 1935. N 5. С. 405-445.
Гуссаковский В.В. Палеарктические виды рода Trypoxylon Latr. (Hym., Sphecidae) // Тр. Зоол. Ин-та АН СССР, 1936. Т. 3. С. 639-667.
Гуссаковский В.В. Обзор палеарктических видов родов Didineis Wesm., Pison Latr. И Psen Latr. (Hymenoptera? Sphecodea) // Тр. Зоол. инта АН СССР, 1937. Т. 4. С. 599-695.
Гуссаковский В.В. Отр. Hymenoptera – перепончатокрылые // В кн.: Определитель насекомых европейской части СССР. М.-Л., 1948. С. 619-782.
Гуссаковский В.В. Новые и малоизвестные виды Psammocharidae и Sphecidae (Hymenoptera) Западного Таджикистана // Тр. Зоол. Ин-та АН СССР, 1952. Т. 10. С. 199-288.
Джанокмен К.А., Казенас В.Л. Хальцид Habritys brevicornis (Ratzeburg) (Hymenoptera, Pteromalidae) – паразит Ectemnius lapidarius (Panz.) (Hymenoptera, Sphecidae) // Тр. Ин-та зоол. АН КазССР, 1974. Т. 35. С. 104-107.
Исламов Ш.Д. Изучение фауны роющих ос рода Sceliphron Klug. (Hymenoptera, Sphecidae) в Узбекистане // Узб. биол. Журн., 1971а. N 4. С. 55-57.
Исламов Ш.Д. Роющие осы (Hymenoptera, Sphecidae) горных районов Узбекистана // Энтомол. обозр., 1986. Т. 65, вып. 3. С. 513-534.
Исламов Ш.Д., Казенас В.Л. Кормовые связи роющих ос (Hymenoptera, Sphecidae) c энтомофильными растениями в горных районах Узбекистана // Узб. биол. журн., 1979. № 4. С. 64-65.
Исламов Ш.Д., Казенас В.Л. Роющие осы рода Cerceris Latr. (Hymenoptera, Sphecidae) горных районов Узбекистана // Узб. биол. журн., 1983. № 5. С. 44-46.
Казенас В.Л. Оса-воровка // Простор, 1964. N 4. С. 44.
Казенас В.Л. Осы уничтожают саранчу // Природа, 1965. N 11. С. 108.
Казенас В.Л. Материалы к изучению ос рода Ammophila (Hymenoptera, Sphecidae) в южных районах Казахстана // В кн.: Материалы Первой научной конференции молодых ученых Академии наук Казахской СССР. Алма-Ата, 1968а. С. 342-343.
Казенас В.Л. К биологии роющей осы Sphex mocsaryi Kohl (Hymenoptera, Sphecidae) // Энтомол. Обозр., 1968б. Т. 47, вып. 4. С. 806-808.
Казенас В.Л. Оса-истребитель цикадок // Защита раст., 1969а. N 1. C. 56.
Казенас В.Л. Материалы к изучению роющих ос рода Sphex L. (Hymenoptera, Sphecidae) в южных районах Казахстана // В кн.: Материалы I научной конференции молодых специалистов и аспирантов. Алма-Ата, 1969б. С. 21-22.
Казенас В.Л. Новый вид ос (род Philanthus F., Sphecidae) из юговосточного Казахстана // В кн.: Материалы Второй научной конференции молодых ученых АН КазССР. Алма-Ата, 1970а. С. 366-367.
Казенас В.Л. К биологии роющей осы Ammophila (Eremochares) dives Brulle (Hymenoptera, Sphecidae) // Энтомол. обозр., 1970б. Т. 49, вып. 2. С. 292-302.
Казенас В.Л. Питание имаго роющих ос // В кн.: Шестой съезд Всесоюзного энтомологического общества. Аннотации докладов. Воронеж, 1970в. С. 76-77.
Казенас В.Л. Роющие осы (Hymenoptera, Sphecidae) юговосточного Казахстана (автореферат канд. дис.). Алма-Ата, 1970г. 20 с.
Казенас В.Л. Роющие осы (Hymenoptera, Sphecidae) Заилийского Алатау // Тр. Ин-та зоол. АН КазССР, 1971. Т. 32. С. 153-162.
. Новыйвид рода Cerceris Latreille (Hymenoptera, Sphe-
cidae) из юго-восточного Казахстана // Зоол. журн., 1972а. Т. 51, вып. 3. С. 454-455.
Казенас В.Л. Роющие осы (Hymenoptera, Sphecidae) ЮгоВосточного Казахстана // В кн.: Насекомые аридных областей СССР и сопредельных стран. Л., 1972б. С. 93-186.
Казенас В.Л. Новый вид из рода Dinetus Panz. (Hymenoptera, Sphecidae) из Алма-Атинского заповедника «Поющая гора» // Тр. заповедн. Казахстана, 1972в. Т. 3. С. 16-18.
Казенас В.Л. Новый вид Lindenius (Hymenoptera, Sphecidae) из юго-восточного Казахстана //Зоол. Журн., 1973. Т. 52, вып. 2. С. 289-290.
Казенас В.Л. Новые виды роющих ос рода Cerceris Latr. (Hymenoptera, Sphecidae) из юго-восточного Казахстана // Тр. Ин-та зоол. АН КазССР, 1974а. Т. 35. С. 97-103.
Казенас В.Л. Питание роющих ос (Hym., Sphecidae) // Тр. Ин-та зоол. АН КазССР, 1974б. Т. 35. С. 108-113.
Казенас В.Л. Роющие осы (Hymenoptera, Sphecidae) – истребители прямокрылых в Казахстане // В кн.: Материалы Седьмого съезда Всесоюзного энтомологического общества. Часть вторая. Л., 1974в. С. 72.
Казенас В.Л. Два новых вида Eremiasphecium (Hymenoptera, Sphecidae) из юго-восточного Казахстана // Зоол. Журн., 1974г. Т. 53, вып. 11. С. 1733-1736.
Казенас В.Л. (Kazenas V.L. A new species of Diploplectron Fox (Hymenoptera, Sphecidae) from Kazakhstan // Pol. pis. entomol., 1975a. T. 45. P. 635-638.)
Казенас В.Л. Роющие осы рода Diodontus Curtis (Hymenoptera, Sphecidae) юго-восточного Казахстана // Энтомол. Обозр., 1975б. Т. 54, вып. 2. С. 421-428.
Казенас В.Л. Роющие осы (Hymenoptera, Sphecidae) юговосточного Казахстана. Алма-Ата: Ин-т зоол. АН КазССР, 1975в. 41 с. (Рукопись деп. в ВИНИТИ 14 апр. 1975 г., N 1061-75 Деп.)
Казенас В.Л. Искусственные гнездовья ос // Защита раст., 1976а. N
11. C. 26.
Казенас В.Л. Роющие осы (Hymenoptera, Sphecidae) АлмаАтинского заповедника «Поющая гора». Алма-Ата: Ин-т зоол. АН КазССР, 1976б. 9 с. (Рукопись деп. в ВИНИТИ 13 мая 1976 г., N 1657-76
Деп.)
. Новый вид рода Dolichurus (Hymenoptera, Sphecidae)
из юго-восточного Казахстана // Зоол. журн., 1976в. Т. 55, вып. 11. С. 1769-1770.
Казенас В.Л. Гнезда ос (Hymenoptera, Sphecidae, Vespidae) в стеблях шиповника и тростниковых трубочках в Северном Прибалхашье. Алма-Ата: Ин-т зоол. АН КазССР, 1976г. 8 с. (Рукопись деп. В ВИНИТИ 3 мая 1976 г., N 1658-76 Деп.)
Казенас В.Л. Новые виды роющих ос рода Cerceris (Hymenoptera,
Sphecidae) из Казахстана // Энтомол. обозр., 1977а. Т. 56, вып. 2. С. 431434.
Казенас В.Л. (Kazenas V.L. A new species of Dinetus Panzer (Hymenoptera, Sphecidae) from Transcaspia, with biological observations // Pol. pis. еntomol., 1977б. T. 47. P. 363-369.)
Казенас В.Л. Роющие осы Казахстана и Средней Азии (Hymenoptera, Sphecidae). Определитель. Алма-Ата: Наука, 1978а. 172 с.
Казенас В.Л. Новые виды роющих ос (Hymenoptera, Sphecidae) из Юго-Восточного Казахстана // Энтомол. обозр., 1978б. Т. 57, вып. 3. С. 661-665.
Казенас В.Л. Роющие осы (Hymenoptera, Sphecidae) Северного Казахстана. Алма-Ата: Ин-т зоол. АН КазССР, 1978в. 34 с. (Рукопись деп. В ВИНИТИ 31 мая 1978 г., N 1807-78 Деп.)
Казенас В.Л. Новый вид рода Larra F. (Hymenoptera, Sphecidae) из юго-восточного Казахстана // Зоол. журн., 1979а. Т. 58, вып. 1. С. 130132.
Казенас В.Л. Новый вид рода Belomicroides Kohl (Hymenoptera, Sphecidae) из Юго-Восточного Казахстана // Тр. Всесоюзн. энтомол. ова, 1979б. Т. 61. С. 172-174.
Казенас В.Л. Новые виды роющих ос рода Cerceris Latr. (Hymenoptera, Sphecidae) из Южного Казахстана // Энтомол. обозр., 1979в. Т. 58, вып. 4. С. 857-859.
Казенас В.Л. Новые виды рода Bembix F. (Hymenoptera, Sphecidae) из Казахстана // В кн.: Насекомые Казахстана. Алма-Ата: Наука, 1980а. С. 137-143.
Казенас В.Л. Новый туранский вид рода Cerceris Latr. (Hymenoptera, Sphecidae) // В кн.: Насекомые Казахстана. Алма-Ата: Наука, 1980б. С. 144-146.
. Новыевиды роющих ос рода Cerceris Latr. (Hymenop-
tera, Sphecidae) из Средней Азии // Зоол. Журн., 1980в. Т. 59, вып. 5. С. 793-794.
Казенас В.Л. Новый вид рода Gastrosericus (Hymenoptera, Sphecidae) из южного Таджикистана // Зоол. Журн., 1980г. Т. 59, вып. 7. С. 1103-1105.
Казенас В.Л. Новые виды роющих ос (Hymenoptera, Sphecidae) из Таджикистана // Изв. АН ТаджССР, отд. Биол. Наук, 1980д. N 4 (81). С.49-55.
Казенас В.Л. Материалы к фауне роющих ос рода Cerceris Latr. (Hymenoptera, Sphecidae) Казахстана и Средней Азии. Алма-Ата: Ин-т зоол. АН КазССР, 1983. 40 с. (Рукопись деп. В ВИНИТИ 29 июня 1983 г., N 3527-83 Деп.)
Казенас В.Л. Роющие осы-церцерисы Средней Азии и Казахстана. Алма-Ата: Изд-во «Наука» КазССР, 1984а. 232 с.
Казенас В.Л. Эволюция типов добычи у роющих ос (Hymenoptera, Sphecidae) // IX съезд Всесоюзн. энтомол. о-ва, Киев; окт. 1984. Тез. докл. Ч. I. Киев, 1984б. С. 206.
Казенас В.Л. Новый вид рода Crabro (Hymenoptera, Sphecidae) из Казахстана // Зоол. журн., 1984в. Т. 63, вып. 8. С. 1271-1273.
Казенас В.Л. (Kazenas V.L. The prey of Laphyragogus turanicus Gussakovskij (Hymenoptera, Sphecidae) // Sphecos, 1985. N 10. P. 17.
Казенас В.Л. Новый вид рода Gorytes Latr. (Hymenoptera, Sphecidae) из Юго-Восточного Казахстана // Зоол. журн., 1987. Т. 66, вып. 12. С. 1899-1900.
Казенас В.Л. Новый вид рода Harpactus (Hymenoptera, Sphecidae) из Южного Казахстана // Зоол. журн., 1988. Т. 67. вып. 1. С. 148-150.
Казенас В.Л. Новые виды роющих ос рода Harpactus (Hymenoptera, Sphecidae) из Казахстана // Зоол. журн., 1989а. Т. 68, вып. 9. С. 148-152.
Казенас В.Л. Два малоизвестных и один новый вид роющих ос рода Harpactus Schuckard (Hymenoptera, Sphecidae) из Средней Азии // Энтомол. обозр., 1989б. Т. 68 , вып. 4. С. 775-777.
Казенас В.Л. Типы гнезд у роющих ос Казахстана и их эволюционные отношения // В кн.: Тез. докл. X съезда ВЭО, Л., 1990а. С. 57-59.
Казенас В.Л. Новые виды роющих ос рода Oxybelus Latreille (Hymenoptera, Sphecidae) из Казахстана // Тр. Ин-та зоол. АН КазССР, 1990б. Т. 45. С. 59-64.
. Taukumia – новый род роющих ос (Hymenoptera,
Sphecidae) из Казахстана // Зоол. журн., 1991а. Т. 70, вып. 5. С. 156-158.
Казенас В.Л. Два новых вида роющих ос рода Belomicrus A.Costa (Hymenoptera, Sphecidae) из Юго-Восточного Казахстана // Зоол. журн., 1991б. Т. 70, вып. 11. С. 145-147.
Казенас В.Л. Сцелифрон Шестакова. Сфекс желтокрылый. Прионикс Хаберхауэра. Прионикс траурный. Лестифорус горолюбивый // Красная книга Казахской ССР. Т. I. Животные. Алма-Ата: Гылым,
1991в. С. 418-426.
Казенас В.Л. Новые виды роющих ос рода Harpactus Shuckard (Hymenoptera, Sphecidae) из Казахстана и Узбекистана // Энтомол. обозр., 1992а. Т. 71, вып. 3. С. 640-645.
Казенас В.Л. Новые виды роющих ос рода Miscophus Jurine (Hymenoptera, Sphecidae) из Казахстана // Зоол. журн., 1992б. Т. 71 , вып. 10. С. 97-107.
Казенас В.Л. Роющие осы (Hymenoptera, Sphecidae) Репетекского гос. заповедника // Изв. АН Туркменистана, 1992в. № 6. С. 24-30.
Казенас В.Л. Новые виды ос рода Belomicrus A.Costa
(Hymenoptera, Sphecidae) из Казахстана // Энтомол. обозр., 1993а. Т. 72, № 1. С. 186-189.
Казенас В.Л. Новые виды роющих ос рода Miscophus (Hymenoptera, Sphecidae) из Туркменистана // Вестник зоол. , 1993б. № 2. С. 28-34.
Казенас В.Л. Роющие осы рода Dinetus Panzer (Hymenoptera, Sphecidae) фауны СССР // Зоологические исследования в Казахстане. Алматы, 1993в. С. 58-65.
Казенас В.Л. Новый вид рода Pseudoscolia Radoszkowski (Hymenoptera, Sphecidae) из Южного Казахстана // Selevinia, 1993г. № 1. С. 5355.
Казенас В.Л. (Kazenas V.L.) New species of the genus Pseudoscolia Radoszkowski from Central Asia (Hymenoptera, Sphecidae) // Russian Entomol. J., 1994a (1993). Vol. 2. № 5-6. Р. 47-57.
Казенас В.Л. Новые виды рода Pseudoscolia Radoszkowski (Hymenoptera, Sphecidae) из Казахстана // Изв. НАН Респ. Каз-н. Сер. биол., 1994б (1993). № 4. С. 29-36.
Казенас В.Л. Новые виды рода Pseudoscolia (Hymenoptera, Sphecidae) из пустыни Каракум (Туркменистан) // Зоол. журн., 1994в. Т. 73 , вып. 4. С. 72-78.
. Новые виды роющих ос рода Pseudoscolia Radosz-
kowski (Hymenoptera, Sphecidae) из Южного Казахстана // Изв. НАН Респ. Каз-н. Сер. биол., 1994г. № 3. С. 36-42.
Казенас В.Л. Новые виды ос рода Oxybelus Latreille (Hymenoptera,
Sphecidae) из Казахстана // Энтомол. обозр., 1994д. Т. 73, № 4. С. 906912.
Казенас В.Л. Необычная добыча роющих ос рода Palarus Latr. (Hym., Sphecidae) // Бюлл. Моск. Общ. Испыт. прир. Биол. 1994е. Т. 99, № 1. С. 115.
Казенас В.Л. Малоизвестные виды роющих ос рода Mimesa из группы shestakovi (Hymenoptera, Sphecidae) из Казахстана // Selevinia, 1995а. № 2. С. 26-29.
Казенас В.Л. Новые виды роющих ос рода Belomicrus A.Costa
(Hymenoptera, Sphecidae) из Казахстана // Энтомол. обозр., 1995б. Т. 74, № 3. С. 701-705.
Казенас В.Л. Род Diploplectron (Hymenoptera, Sphecidae) в Казахстане // Зоол. журн., 1996. Т. 75, вып. 6. С. 936-939.
Казенас В.Л., Александер Б. (Kazenas V.L., Alexander B.) The nest, prey, and larva of Entomosericus kaufmanni Radoszkowski (Hymenoptera, Sphecidae) // Intern. J. Hymenopterology, 1993. Vol. 2, № 1. P. 221-225.
Казенас В.Л., Антропов А.В. Новые роющие осы рода Belomicrus (Hymenoptera, Sphecidae) азиатской части Палеарктики. I. Виды групп italicus и radoszkowskii // Зоол. журн., 1994а. Т. 73, вып. 1. С. 68-77.
Казенас В.Л., Антропов А.В. Новые роющие осы рода Belomicrus (Hymenoptera, Sphecidae) азиатской части Палеарктики. 2. Виды группы kohlii // Зоол. журн., 1994б. Т. 73, вып. 1. С. 78-88.
Казенас В.Л., Джанокмен К.А. К биологии роющей осы Ammophila (Eremochares) dives Br. (Hymenoptera, Sphecidae) // В кн.: Материалы Научной конференции молодых биологов гю Алма-Аты. Алма-Ата,
1970. С. 101-102.
Казенас В.Л., Джанокмен К.А. Хальцид Habritys brevicornis (Ratz.) (Hymenoptera, Pteromalidae) – паразит Ectemnius lapidarius (Panz.) (Hymenoptera, Sphecidae) // Тр. Ин-та зоол. АН КазССР, 1974. Т. 35. С. 104107.
Казенас В.Л., Есенбекова П.А. Цикадовые (Homoptera, Auchenorrhyncha) – добыча роющих ос (Hymenoptera, Sphecidae) в Казахстане // Selevinia, 1995. № 3. С. 79-81.
., Зонштейн С.Л., Милько Д.А. Надсем. Sphecoidea -
сфекоидные осы // В кн.: Кадастр генетического фонда Кыргызстана. Том III. Бишкекб 1996. С. 381-385.
Казенас В.Л., Насырова С.Р. Роющие осы (Hymenoptera, Sphecidae) – враги прямокрылых (Orthoptera) в пустынной зоне Казахстана // Изв. АН КазССР. Сер. биол., 1991. № 6. С. 37-40.
Казенас В.Л., Тобиас В.И. Ночевочные скопления роющих ос (Hymenoptera, Sphecidae) в Юго-Восточном Казахстане // Энтомол. обозр., 1992. Т. 71, вып. 1. С. 28-31.
Колесников В.А. Роющие осы (Hymenoptera, Sphecidae) Брянской области и их значение как энтомофагов // Энтомол. Обозр., 1977. Т. 56, вып. 2. С. 315-325.
Мальковский М.П. Роль роющей осы – Tachysphex pompiliformis (Panz.) в снижении численности атбасарской кобылки Dociostaurus kraussi Ingen // Тр. Каз. НИИ защиты раст., 1962. Т. 7. С. 342-346.
Малышев С.И. Перепончатокрылые, их происхождение и эволюция. М.: Советская наука, 1959. 297 с.
Малышев С.И. Становление перепончатокрылых и фазы их эволюции. М.-Л.: Наука, 1966. 330 с.
Мариковский П.И. Этюды по биологии насекомых // Изв. АН
Кирг.ССР, сер. биол. наук, 1961. Т. 3, вып. 1. С. 219-221.
Мариковский П.И. Целебный огонь. М.: Детгиз, 1963. 126 с.
Мариковский П.И. Загадочное излучение. Алма-Ата: изд-во «Наука» КазССР, 1972. 168 с.
Маршаков В.Г. Новые виды роющих ос рода Lindenius (Hymenoptera, Sphecidae) фауны СССР // Зоол. журн., 1973а. Т. 52, вып. 7. С. 10921097.
Маршаков В.Г. Новые виды роющих ос рода Lindenius (Hymenoptera, Sphecidae) фауны СССР. Сообщение 2 // Зоол. журн., 1973б. Т. 52, вып. 9. С. 1420-1422.
Маршаков В.Г. Обзор роющих ос трибы Crabronini (Hymenoptera, Sphecidae) фауны CCCР. Род Crabro Fabricius, 1775. // Энтомол. Обозр., 1975а. Т. 54, вып. 4. С. 854-872.
Маршаков В.Г. Роющие осы трибы Crabronini (Hymenoptera, Sphecidae) фауны Монголии // В кн.: Насекомые Монголии. Л., 1975б. С. 326336.
Маршаков В.Г. Обзор родов трибы Crabronini (Hymenoptera, Sphecidae) фауны СССР. Род Rhopalum Stephens, 1829. // Тр. Зоол. Ин-та АН СССР, 1976а. Т. 67. С. 100-112.
Маршаков В.Г. Новые виды роющих ос (Hymenoptera, Sphecidae) // Зоол. журн., 1976б. Т. 55, вып. 6. С. 938-941.
Маршаков В.Г. Роющие осы родов Eremiasphecium Kohl, Ammoplanus Guss. и Anomiopteryx Guss. (Hymenoptera, Sphecidae) фауны СССР и Монголии // Энтомол. Обозр., 1976в. Т. 55, вып. 3. С. 668-683.
Маршаков В.Г. Обзор родов трибы Crabronini (Hymenoptera, Sphecidae) фауны СССР. 3. Род Entomognathus Dahlbom // Зоол. журн., 1976г. Т. 55, вып. 4. С. 614-618.
Маршаков В.Г. Роющие осы подсемейства Crabroninae (Hymenoptera: Sphecidae) фауны СССР. Дис. соиск. учен. степ. канд. биол. наук. Ленинград: Зоол. ин-т АН СССР, 1976д. 24 с.
Маршаков В.Г. Роющие осы родов Protostigmus Turner и Ammoplanus Giraud (Hymenoptera, Sphecidae) фауны Монголии и Средней Азии // В кн.: Насекомые Монголии. Вып. 6. Л., 1979а. С. 362-374.
Маршаков В.Г. Роющие осы родов Crossocerus Lepeletier et Brulle, 1834, Ectemnius Dahlbom, 1845 и Towada Tsuneki, 1970 (Hymenoptera, Sphecidae) Дальнего Востока СССР // В кн.: Наземные членистоногие Дальнего Востока. Владивосток, 1979б. С. 90-107.
Маршаков В.Г. Роющие осы рода Crossocerus Lepeletier et Brulle (Hymenoptera, Sphecidae) фауны Монголии, Казахстана и Средней Азии // В кн.: Насекомые Монголии. Вып. 7. Л., 1980а. С. 336-365.
Маршаков В.Г. Роющие осы подрода Othyreus Marshakov рода Crabro Fabricius (Hymenoptera, Sphecidae) // В кн.: Насекомые Монголии. Вып. 7. Л., 1980б. С. 327-335.
Митяев И.Д., Матесова Г.Я., Куленова К.З., Казенас В.Л., Жданко А.Б. Список насекомых (Auchenorrhyncha, Coccoidea, Chrysomelidae, Sphecidae, Lepidoptera) поймы Алея. Алма-Ата: Ин-т зоол. АН КазССР. 1979. 36 с. (Рукопись деп. в ВИНИТИ 4 июня 1979 г. № 1936-79Деп.).
Мярцева С.Н. Экологическое распределение роющих ос (Hymenoptera, Sphecidae) в низовьях Мургаба // Изв. АН ТССР, сер. биол. наук, 1963а. N 4. С. 56-63.
Мярцева С.Н. К биологии и гнездовому поведению ос – Ammophila ebenina Spin. (Hymenoptera, Sphecidae, Sphecinae) // Изв. АН ТССР, сер. биол. наук, 1963б. N 2. С. 62-71.
Мярцева С.Н. К экологии некоторых роющих ос рода Sphex L. (Hymenoptera, Sphecidae) низовий Мургаба // Изв. АН ТССР, сер. биол. наук, 1964а. № 2. С. 73-77.
Мярцева С.Н. Развитие Gomodontochrysis chloe Sem. в гнездах роющих ос (Hymenoptera, Sphecidae) // Изв. АН ТССР, сер. биол. наук, 1964б , N 1 . С.
Мярцева С.Н. Возможности использования сфецид для снижения численности вредных насекомых // Изв. АН ТССР, сер. биол. наук, 1966. № 6. С. 48-53.
Мярцева С.Н. Эколого-фаунистичекая характеристика сфецид трибы Bembicini (отряд перепончатокрылые, семейство роющие осы) равнинной части Туркмении // В кн.: Тезисы докладов Всесоюзной научной конференции по изучению и освоению пустынных территорий Средней Азии и Казахстана. Секция 2. Ашхабад, 1968а. С. 32-33.
Мярцева С.Н. Роющие осы рода Sceliphron Klug. (Hymenoptera, Sphecidae) Туркмении // Изв. АН ТССР, сер. биол. наук, 1968б , № 2 . С. 61-66.
Мярцева С.Н. Особенности биологии и этологии сфецид трибы Ammophilini (Hymenoptera, Sphecidae) в Туркмении // Изв. АН ТССР, сер. биол. наук, 1969. № 3. С. 48-56.
Мярцева С.Н. Зоогеографическая характеристика фауны сфецид (Hymenoptera, Sphecidae) Туркмении // Тр. XIII Междунар. энтомол. конгр., 1971. Т. 1. С. 179-180.
Мярцева С.Н. Фауна сфецид южной Туркмении // В кн.: Насекомые южной Туркмении. Ашхабад, 1972а. С. 75-100.
Мярцева С.Н. Паразиты и хищники роющих ос Туркмении // В кн.: Насекомые южной Туркмении. Ашхабад, 1972б. С. 111-116.
Мярцева С.Н. Заметки по биологии малоизученных сфецид Туркмении (Hymenoptera, Sphecidae) // В кн.: Экология и хозяйственное значение насекомых Туркмении. Ашхабад, 1976. С. 72-82.
Мярцева С.Н., Чарыкулиев Д.М. К экологии мух подсем. Miltogrammatinae (Diptera, Sarcophagidae) // Изв. АН ТССР, сер. биол. наук, 1964а. N 2 . С. 84-88.
Мярцева С.Н., Чарыкулиев Д.М. Новые данные об инквилинах роющих ос // В кн.: Тезисы докладов Первой республиканской конференции молодых зоологов Туркменистана. Ашхабад, 1964а. С. 74-75.
Мярцева С.Н., Чарыкулиев Д.М. Естественные враги пчелиного волка // Колх.-совх. производство Туркменистана, 1964б. № 5. С. 36-37.
Немков П.Г. К фауне роющих ос (Hymenoptera, Sphecidae) Прибайкалья // В сб.: Перепончатокрылые Восточной Сибири и Дальнего Востока. Владивосток, 1986. С. 92-110.
Немков П.Г. Роющие осы трибы Gorytini (Hymenoptera, Sphecidae) фауны СССР. Роды Gorytes Latreille, Pseudoplisus Ashmead, Kohlia Handlirsch // Энтомол. обозр., 1990. Т. 69, вып. 3. С. 675-690.
Немков П.Г. Роющие осы трибы Gorytini (Hymenoptera, Sphecidae) фауны России и сопредельных стран. Роды Lestiphorus Lepeletier, Oryttus Spinola и Olgia Radoszkowski // Энтомол. обозр., 1992. Т. 71. С. 935-949.
Немков П.Г. Роющие осы трибы Gorytini (Hymenoptera, Sphecidae) фауны России и сопредельных стран. Роды Sphecius Dahlbom и Ammatomus A.Costa // Энтомол. обозр., 1995. Т. 74, вып. 1. С. 177-185.
Немков П.Г., Казенас В.Л., Будрис Э.Р., Антропов А.В. Надсем. Sphecoidea. 67. Сем. Sphecidae – Роющие осы // В кн.: Определитель насекомых Дальнего Востока России. Т. IV. Часть I. С.-Пб.: Наука, 1995. С. 368-480.
Нестеров М.А. Два новых вида роющих ос рода Gorytes (Hymenoptera, Sphecidae) из Казахстана // Журн. Укр. ентомол. тов-ва, 1993. Т. 1, № 1. С. 51-54.
Пулавский В.В. Надсем. Sphecoidea. 1. Сем. Sphecidae – Роющие осы // В кн.: Определитель насекомых европейской части СССР. Т. 3. Перепончатокрылые. Первая часть. Л.: Наука, Ленинградское отделение, 1978. С. 173-279.
Радошковский О.И. Sphegidae // В кн.: Путешествие в Туркестан
А.П.Федченко. Т. 2. Зоогеографические исследования. Ч. 5. 1877. С. 1-
87.
Расницын А.П. Происхождение и эволюция перепончатокрылых насекомых. М.: Наука, 1980. 192 с.
Смирнов Д.О. О нравах Ammophila (Eremochares) dives Brulle // Рус. энтомол. обозр., 1915. Т. 15. С. 153-155.
Тобиас В.И. 27. Отряд Hymenoptera – Перепончатокрылые. Введение // В кн.: Определитель насекомых европейской части СССР. Том III. Перепончатокрылые. Первая часть. Л.: Наука, 1978. С. 7-42.
Шестаков А.В. Новые палеарктические виды рода Cerceris Latr. (Hymenoptera, Crabronidae) // Рус. энтомол. обозр., 1912. Т. 12, вып. 3. С. 507-510.
Шестаков А.В. Новые палеарктические виды рода Cerceris Latr. (Hymenoptera, Crabronidae) // Рус. энтомол. обозр., 1914а. Т. 14, вып. 1. С. 90-99.
Шестаков А.В. Род Cerceris Latr. (Hymenoptera, Crabronidae) в коллекции Зоологического музея Московского университета // Рус. энтомол. обозр., 1914б. Т. 14, вып. 4. С. 408-411.
Шестаков А.В. О новых видах рода Сerceris Latr. (Hymenoptera, Crabronidae) // Рус. энтомол. обозр., 1915. Т.15, вып. 1. С. 8-15.
Шестаков А.В. Материалы для фауны ос рода Cerceris Latr. (Hymenoptera, Crabronidae) Туркестана // Ежегодник Зоол. муз. Росс. Акад. наук, 1918 (1917). Т. 22. С. 118-166.
Шестаков А.В. Новые виды рода Cerceris Latr. (Hymenoptera, Crabronidae) в коллекциях Зоологического музея Российской академии наук // Ежегодник Зоол. муз. Росс. Акад. наук, 1922 (1918). Т. 23. С. 1-31.
Шестаков А.В. Обзор палеарктических видов ос нового подрода Apiraptrix (Hymenoptera, Crabronidae). Ярославль: изд-во Яросл. Губ. С.х. И куст.-промысл. Союз. Кооперативов, 1923. 15 с.
Шестаков А.В. К синонимии видов Cerceris Latr. (Hymenoptera, Crabronidae) // Рус. Энтомол. Обозр., 1925. Т. 19. С. 239-240.
Шестаков А.В. Очерк географического распространения ос подсем. Philanthinae (Hymenoptera, Crabronidae) в пределах Европейской части СССР. Ярославль: Яросл. Пед. Ин-т, 1927а. 20 с.
Шестаков А.В. (Shestakov A. Neue Cerceris Arten aus der Gruppe Cerceris tuberculata Vill. (Hymenoptera) // Soc. entomol., 1927б. Vol. 42. P. 30-31.)
Abrahamsen S.E. Danske gravehvespe // Natur og mus. (Dan.), 1972. Bd 15, N 3. S. 3-22.
Alexander B.A. An exploratory analysis of cladistic relationships within the superfamily Apoidea with special reference to sphecid wasps (Hymenoptera) // J. Hym. Res., 1992. Vol. 1, N 1. P. 25-61.
Alfieri A. Les especes egyptiennes du genre Ammophila Kirby (Hymenoptera – Aculeata: Sphecoidea – Sphecidae // Bull. Soc. Fouad 1er Entomol., 1946. Vol. 30. P. 105-142.
Bajari E.N. Kaparodarazs Alkatuak. I. (Sphecoidea I). Budapest: Akad. kiado, 1957. 117 l.
Balthasar V. Fauna CSSR. Bd 20. Grabwespenn – Sphecoidea. Praha:
Verl. Tschechosl. Acad. Wissensch., 1972. 471 S.
Beaumont J. de. Le genre Olgia Radoszk. (Hym. Sphecid.) // Rev. suisse zool., 1953a. T. 60, fasc. 2, N 3-4. P. 205-223.
Beaumont J. de. Synonymie de quatre genres de Sphecidae decrits par
Gussakovskij (Hym.) // Mitt. Schweiz. entomol. Ges., 1955. Bd 28, H. 2. S. 222-223.
Beaumont J. de. Sphecidae du nord de l’Iran (Hym.) // Mitt. Schweiz. Entomol. Ges., 1957. Bd 30, H. 2. S. 127-139.
Beaumont J. de. Contribution a l’etude de la faune d’Afghanistan 39. Sphecidae // Mitt. Schweiz. Entomol. Ges., 1960. Bd 33, H. 3. S. 169-172.
Beaumont J. de. Ergebnisse der Deutschen Afghanistan-Expedition 1956 der Landessammlungen fur Naturkunde Karlsruhe (Sphecidae, Hymenoptera) // Beitr. Naturk. Forsch. SW-Deutschl., 1961. Bd 19, H. 3. S. 271276.
Beaumont J. de. Notes sur les Sphex palearctiques du Sousgenre Chlorion Latr. (Hym. Sphecidae) // Boll. Ist. entomol. Univ. studi Bologna, 1962. Vol. 26. P. 29-41.
Beaumont J. de. Some Sphecidae collected by Klapperich in Afghanistan // Ann. Hist.-Natur. Mus. Nat. Hung., 1963. T. 55, ps. Zoologica. P. 417419.
Beaumont J. de. Insecta Helvetica. Fauna 3. Hymenoptera: Sphecidae. Lausanne, 1964. 170 p.
Beaumont J. de. Beitrage zur Kenntnis der Fauna Afghanistans. Sphecidae, Hym. // Cas. Morav. Mus. 1969. Roc. 54. S. 385-406.
Beaumont J. de. Sphecidae de l’Iran (Hym.) // Stuttgart. Beitr. Naturk., 1970. № 220. P. 1-18.
Becker A. Reise in die Kirgisensteppe, nach Astrachan und an das Caspische Meer // Bull. Soc. imp. natur. Mosc., 1866. Vol. 39, N 2. P. 163-207.
Berland L., Berland F. Les Sphex de France (Hym., Sphec.) // Ann. Soc. entomol. Fr., 1947. Vol. 116. P. 1-16.
Bitsch J. Captures de Chalybion femoratum dans le Midi de la France et observation d’un nid // Bull. Soc. entomol. Fr., 1988. Vol. 93. P. 43-51.
Bischoff H. Die Biologie der Hymenopteren. Berlin: Biol. Studienbucher, 1927. 598 S.
Bischoff H. Hymenoptera I. Entomologische Ergebnisse der DeutschRussischen Alai-Pamir-Expedition 1928 (1). // Mitt. Zool. Mus. Berlin, 1930.
Bd 16, H. 2. S. 215-225.
Bohart R.M. and Menke A.S. A Reclassification of the Sphecinae with a revision of the nearctic species of the tribes Sceliphronini and Sphecini (Hymenoptera, Sphecidae). Berkeley; Los Angeles: University of California Press, 1963. 182 p.
Bohart R.M. and Menke A.S. Sphecid wasps of the World. A generic revision. Berkeley; Los Angeles; London: University of California Press, 1976. 696 p.
Brothers D.J. Phylogeny and classification of the aculeate Hymenoptera, with special reference to Mutillidae // Univ. Kansas Sci. Bull., 1975. Vol. 50, N 11. P. 483-648.
Ceballos G. Catalogo de los Himenopteros de Espana. Madrid, 1956. 554 p.
Dalla Torre C.G. Catalogus Hymenopterorum. Vol. VII. Fossores (Sphecidae). Lipsiae, 1897. p.
Dollfuss H. Fam. Sphecidae // In: Catalogus Faunae Austriae. Teil XVII. Wien: Verl. Osterrl. Akad. Wiss., 1983. 32 S.
Dollfuss H. Eine Revision der Gattung Spilomena Shuckard der westlichen und zentralen palaarktischen Region (Hymenoptera, Sphecidae) // Ann. Naturhist. Mus. Wien, 1986. Bd. 88/89. S. 481-510.
Dollfuss H. Bestimmungsschussel der Grabwespen Nord- und Zentraleuropas (Hymenoptera, Sphecidae) mit speziellen Angaben zur Grabwespenfauna Osterreichs // Stapfia, 1991. Vol. 24. S. 1-247.
Dollfuss H. A worldwide revision of Pemphredon Latreille 1796 (Hymenoptera, Sphecidae) // Linzer biol. Beitr., 1995. Bd 27, N 2. S. 905-1019.
Evans H.E. Ethological studies on digger wasps of the genus Astata (Hymenoptera, Sphecidae) // J. N. Y. Entomol. Soc., 1957f. Vol. 65, N 3-4. P. 159-185.
Evans H.E. Studies on the comparative ethology of digger wasps of the genus Bembix. Ythaca, New Jork: Comstock Publ. Assoc., 1957b. 248 p.
Eversmann E. Fauna hymenopterologica Volgo-Uralensis. Fam. III. Sphegidae Latr. // Bull. Soc. imp. natur. Mosc., 1849. T. 22, N IV. P. 359436.
Friese H. Die Insekten Mitteleuropas insbesonders Deutschlands. Bd I. Hymenopteren. Erster Teil. Die Bienen, Wespen, Grab- und Goldwespen. Stuttgart, 1926. 192 S.
Grandi G. Studi di un entomologo sugli Imenotteri superiori // Boll. Ist.
entomol. Univ. studi Bologna, 1961. Vol. 25. P. 167-180.
Handlirsch A. Monographie der mit Nysson und Bembex verwandten Grabwespen // S. B. Akad. Wiss., Math. Naturw. Cl., 1987a. Bd 95. S. 246421; 1887b. Bd 96. S. 219-311; 1888. Bd 97. S. 316-565; 1889a. Bd 98. S. 441-517; 1890. Bd 99. S. 77-201; 1892. Bd 100. S. 25-204; 1893. Bd 102. S. 657-942.
Hensen R.V. Revision of the subgenus Prosceliphron van der Vecht (Hymenoptera, Sphecidae) // Tijjdschr. Entomol., 1987. Vol. 129. P. 217-261.
Hensen R.V. Revision of the nominate subgenus Chalybion Dahlbom (Hymenoptera, Sphecidae) // Tijjdschr. Entomol., 1988. Vol. 131. P. 13-64.
Iwata K. Evolution of Instinct. Comparative Ethology of Hymenoptera. New Delhi: Amerind Publ. Co. Pvt Ltd, 1976. IX+539 p.
Kohl F.F. Die Gattungen der Sphecinen und die palaearktischen SphexArten // Termesz. Fuz., 1885. Kot. 9, fuz. 2. L. 154-207.
Kohl F.F. Die Hymenopterengruppe der Sphecinen. I-II. Monographie der naturlichen Gattung Sphex L. (sens lat.) // Ann. Naturhist. Hofmus. Wien, 1890. Bd 5. S. 77-194, 319-462.
Kohl F.F. Zur Kenntniss der Hymenopteren-Gattung Philanthus Fabr. (sens. lat.) // Ann. Naturhist. Hofmus. Wien, 1891. Bd 6. S. 345-370.
Kohl F.F. Zur Monographie der naturlichen Gattung Sphex Linne // Ann. Naturhist. Hofmus. Wien, 1895. Bd 10. S. 42-74.
Kohl F.F. Die Gattungen der Sphegiden // Ann. Naturhist. Hofmus. Wien, 1896. Bd 11. S. 233-516.
Kohl F.F. Zur Kenntnis der palaarktischen Diodontus-Arten // Verh. Zool.-bot. Ges. Wien, 1901a. Bd 51, S. 120-134.
Kohl F.F. Ueber neue Arten der Hymenopteren-Gattung Tachysphex Kohl // Verh. Zool.-bot. Ges. Wien, 1901b. Bd 51. S. 777-784.
Kohl F.F. Die Hymenopterengruppe der Sphecinen. III. Monographie der Gattung Ammophila (sens lat.) // Ann. Naturhist. Hofmus. Wien, 1906. Bd 21. S. 228-382.
Kohl F.F. Die Crabronen der Palearktischen Region // Ann. Naturhist. Hofmus. Wien, 1915. Bd 29. S. 1-453.
Kohl F.F. Beitrag zur Kenntnis der Gattung Cerceris auf Grundlage der
O.Radoszkowskischen Sammlung // Arch. Naturgesch., 1916 (1915). Bd 88,
H. 7. S. 107-125.
Kohl F.F. Die Hymenopterengruppe der Sphecinen. VI. Die naturliche Gattung Sceliphron Klug (=Pelopoeus Latr.) // Ann. Naturhist. Hofmus.
Wien, 1918. Bd 32. S. 1-171.
Kohl F.F. Gie Hymenopteren-Gattung Belomicrus A.Costa // Konovia, 1923. Bd 2. S. 98-122, 180-202, 258-278.
Kohl F.F., Handlirsch A. Transcaspische Hymenopteren // Verh. Zool.bot. Ges. Wien, 1889. Bd 39. S. 267-286.
Lomholdt O. The Sphecidae (Hymenoptera) of Fennoscandia and
Denmark // In: Fauna Entomol. Scand., vol. 4. Klampenborg, 1975-1976. 450
S.
Lomholdt O. On the origin of the bees (Hymenoptera: Apidae, Sphecidae) // Entomol. Scand., 1982. Bd 13. S. 185-190.
Lomholdt O. A reclassification of the larrine tribes with a revision of the Miscophini of southern Africa and Madagascar (Hymenoptera; Sphecidae) // Entomol. Scand., Suppl., 1985. P. 1-183.
Menke A.S. The identity of Parapsammophila lutea Taschenberg, 1869 (Hymenoptera, Sphecidae) // Pol. pis. entomol., 1966. T. 36, N 4. P. 57-61.
Merisuo A.K. Die im Holz und im Boden nistenden finnischen Raubwespen (Hym., Sphecidae), ihre Nesttypen und Provianttiere // Ann. Entomol. Fenn., 1943. Bd 9, N 4. S. 219-260.
Merisuo A.K. Zur Kenntnis der Pemphredon Latreille-Arten (Hym., Sphecoidea) // Ann. entomol. fenn., 1972. V. 38, N 4. S. 190-192.
Merisuo A.K. Zur Bilogie von Passaloecus clypealis Faester (Hym., Sphecoidea) // Ann. entomol. fenn., 1973. V. 39, N 2. S. 103-108.
Merisuo A.K. Drei neue asiatische Arten zur Gattung Passaloecus Shuckard (Hym., Sphecoidea) // Ann. entomol. fenn., 1976. V. 42, N 2. S. 171-177.
Morawitz F. Hymenoptewra aculeata nova // Hor. Soc. entomol. ross., 1888. T. 22. P. 224-302.
Morawitz F. Die russischen Thyreus Arten // Hor. Soc. entomol. ross., 1889. T. 23. P. 540-554.
Morawitz F. Hymenopteras fossoria transcaspica nova // Hor. Soc. entomol. ross., 1890. T. 24. P. 570-645.
Morawitz F. Ueber astrachan'sche Fossoria // Hor. Soc. entomol. ross., 1891. T. 25. P. 174-233.
Morawitz F. Hymenoptera aculeata rossica nova // Hor. Soc. entomol. ross., 1892. T. 26. P. 132-181.
Morawitz F. Catalog der von D.Glasunov in Turkestan gesammelten Hymenoptera Fossoria // Hor. Soc. entomol. ross., 1893a. T. 27. P. 361-428.
Morawitz F. Neue Hymenopteren vom Pamir // Hor. Soc. entomol. ross., 1893b. T. 27. P. 429-433.
Morawitz F. Beitrag zur Raubenwespenfauna Turkmeniens // Hor. Soc. entomol. ross., 1894. T. 28. P. 327-365.
Morawitz F. Ueber einige transcaspische Raubenwespen // Hor. Soc. entomol. ross., 1896. T. 30. P. 144-160.
Noskiewicz J., Pulawski W. Klucze do oznaczania owadow Polski. Cz. 24. Blonkowki – Hymenoptera. Zeszyt 67. Grzebaczowate – Sphecidae. Warszawa, 1960. 185 s.
Oehlke J. Beitrage zur Insektenfauna der DDR. Hymenoptera – Sphecidae // Beitr. Entomol., 1970. Bd 20, H. 7/8. S. 615-812.
Olberg G. Das Verhalten der solitaren Wespen Mitteleuropas (Vespidae, Pompilidae, Sphecidae). Berlin: Dtsch. Verl. Wiss., 1959. XIII+401 S.
Olberg G. Brutfursorge und Koloniales Verhalten der Heuschreckennnnnsandwespe Sphex maxillosus // Natur und Museum, 1966. Bd 96. S. 1-
8.
Pulawski W. Grzebacze (Hymenoptera, Sphecidae) zebrane w czasie podrozy do Bulgarii // Pol. Pismo Entomol., 1958 (1957). N. 27, N 20. P. 161-192.
Pulawski W.J. Les Tachytes Panz. De la region palearctique occidentale et centrale (Hym., Sphecidae) // Pol. Pis. Entomol., 1962. T. 32, N 22. P. 311-475.
Pulawski W. Badania nad egipskimi Sphecidae (Hym.) // Pol. Pis. Entomol., 1964. T. 34, N 6. P. 63-155.
Pulawski W.J. O synonimice niektorych palearktycznych Sphecidae (Hym.) // Pol. Pis. Entomol., 1965a. T. 35, N 19. P. 563-578.
Pulawski W.J. Diploplectron asiaticum sp. n. (Hym., Sphecidae) // Pol. Pis. Entomol., 1965b. T. 35, N 4. P. 221-224.
Pulawski W. Les Tachysphex Kohl (Hym., Sphecidae) de la region palearctique occidentale et centrale. Wroclaw: Panstw. Wyd. Nauk, 1971. 464 p.
Pulawski W.J. Les Ammatomus A.Costa (Hym., Sphecidae) de la region palearctique occidentale et centrale // Pol. Pis. Entomol., 1973. T. 43, N 2. P. 273-287.
Pulawski W.J. Synonymical notes on Larrinae and Astatinae (Hymenoptera: Sphecidae) // J. Wash. Acad. Sci., 1974. Vol. 64, N 4. P. 308-323.
Pulawski W.J. A revision of the Old World Parapiagetia Kohl (Hym., Sphecidae) // Pol. Pis. Entomol., 1977. T. 47. P. 601-669.
Pulawski W.J. Two new synonyms in transcaspian Sphecidae (Hymenoptera) // Pol. Pis. Entomol., 1979a. T. 49. P. 303-304.
Pulawski W.J. A revision of the World Prosopigastra Costa (Hymenoptera, Sphecidae) // Pol. Pis. Entomol., 1979b. T. 49. P. 3-134.
Pulawski W.J. New synonyms in Old World Sphecidae (Hymenoptera) // Mitt. Schweiz. entomol. Ges., 1981. Bd 54. S. 363-366.
Pulawski W.J. Identification and synonymies of two western palearctic Cerceris: maculata Radoszkowski and hathor n. Sp. (Hymenoptera: Sphecidae) // Pan-Pacif. Entomol., 1983. Vol. 59. N 104. P. 240-245.
Pulawski W.J. The status of Trypoxylon figulus (Linnaeus, 1758), medium de Beaumont, 1945, and minus de Beaumont, 1945 (Hymenoptera: Sphecidae) // Proc. Calif. Acad. Sci., 1984. Vol. 43, N 10. P. 123-140.
Pulawski W.J. The wasp genus Gastrosericus Spinola, 1839 (Hymenoptera: Sphecidae) // Mem. California Acad. Sci, 1995. N 18. P. 1-173.
Radoszkowski O. Materiaux pour servir a une faune hymenopterologique de la Russie (Suite) // Hor. Soc. entomol. ross., 1876. T. 12. P. 82-110.
Radoszkowski O. Faune hymenopterologique Transcaspienne // Hor. Soc. entomol. ross., 1886. T. 20. P. 3-56.
Radoszkowski O. Faune hymenopterologique Transcaspienne (suite) / Hor. Soc. entomol. ross., 1887. T. 21. P. 88-101.
Radoszkowski O. Etudes hymenopterologique // Hor. Soc. entomol. ross., 1888a. T. 22. P. 315-331.
Radoszkowski O. Faune hymenopterologique Transcaspienne // Hor. Soc. entomol. ross., 1888b. T. 22. P. 338-349.
Radoszkowski O. Faune hymenopterologique Transcaspienne // Hor. Soc. entomol. ross., 1893a. T. 27. P. 38-81.
Radoszkowski O. Faune hymenopterologique Transcaspienne. Supplement // Hor. Soc. entomol. ross., 1893b. T. 27. P. 490-493.
Roth P. Les Sphex de l'Afrique du Nord // Ann. Soc. entomol. Fr., 1925. T. 94, N 4. P. 365-404.
Roth P. Les Ammophiles de l'Afrique du Nord // Ann. Soc. entomol.
Fr., 1928. T. 97. P. 153-169, 229-232.
Roth P. Les Ammophiles de l'Espagne (Hymenoptera-Sphegidae) // Eos, 1929. T. 5. P. 161-190.
Roth P. Les Sphecius palearctiques // Ann. Soc. entomol. Fr., 1948. T. 118. P. 79-94.
Roth P. Les Sphecius palearctiques // Bull. Soc. entomol. Fr., 1959. Vol. 64. P. 68-79.
Roth P. Les Sphex palearctiques du sous-genre Palmodes // Mem. Mus. Nat. Hist. Natur., nouv. Ser., 1963. T. 18, fasc. 2. P. 139-186.
Schletterer A. Die Hymenopteren-Gattung Cerceris Latr. Mit vorzugsweiser Berucksichtigung der palaarktischen Arten // Zool. Jahrber., 1887. Bd 2. S. 349-510.
Schletterer A. Beitrag zur Kenntniss der Hymenopteren-Gattung Cerceris Latr. // Zool. Jahrber., Abt. Syst., 1889a. Bd 4. S. 1124-1131.
Schletterer A. Nachtragliches uber die Hymenopteren-Gattung Cerceris Latr. // Zool. Jahrber., Abt. Syst., 1889b. Bd 4. S. 879-904.
Schmiedeknecht O. Die Hymenopteren Nord- und Mitteleuropas. Jena: Verl. G.Fischer, 1930. X+1062 S.
Tsuneki K. Ethological studies on the Japanese species of Pemphredon, with notes on their parasites, Ellampus spp. // J. Fac. Sci. Hokkaido Univ., Ser. VI, Zool., 1952. Vol. 11, N 1. P. 57-75.
Tsuneki K. Comparative studies on the nesting biology of the genus Sphex (s.l.) in East Asia (Hymenoptera, Sphecidae) // Mem. Fac. Lib. Arts. Fukui Univ., Ser. II, Natur. sci., 1963. N 13, pt. 2. P. 13-78.
Tsuneki K. The biology of Ammophila in east Asia (Hym., Sphecidae) // Etizenia, 1968. N 33. P. 1-64.
Tsuneki K. Spheciden aus inneren Mongolei und dem nordlichen China (Hymenoptera) // Etizenia, 1971a. N 58. P. 1-38.
Tsuneki K. Ergebnisse der Zoologischen Forschungen von Dr. Z.Kaszab in der Mongolei. 239. Sphecidae (Hymenoptera). I-II // Acta Zool. Acad. sci. Hung., 1971b. T. 17, N 1-2. P. 139-217.
Tsuneki K. Ergebnisse der Zoologischen Forschungen von Dr. Z.Kaszab in der Mongolei. 259. Sphecidae (Hymenoptera). III // Acta Zool. Acad. sci. Hung., 1971c. T. 17, N 3-4. P. 409-453.
Tsuneki K. Ergebnisse der Zoologischen Forschungen von Dr. Z.Kaszab in der Mongolei. 280. Sphecidae (Hymenoptera). IV-V // Acta Zool. Acad. Sci. Hung., 1972a. T. 18, N 1-2. P. 147-232.
Tsuneki K. Ergebnisse der Zoologischen Forschungen von Dr. Z.Kaszab in der Mongolei. 284. Sphecidae (Hymenoptera). VI // Acta Zool. Acad. Sci. Hung., 1972b. T. 18, N 3-4. P. 381-416.
Vecht J. van der. Uber Taxonomie und Evolution der Grabwespengattung Sceliphron Klug (Hymenoptera, Sphecidae) // Verhandl. XI. Internat. Kongress Entomol. (Wien, 1960), 1961. Bd 1. S. 251-256.
Vecht J. van der, Breugel F.M.A. van. Revision of the nominate subgenus Sceliphron Latreille (Hym., Sphecidae) // Tijdshr. entomol., 1968. D. III, N 6. S. 185-255.
White E. Nest-building and provisioning in relation to sex in Sceliphron spirifex L. (Sphecidae) // J. Anim. Ecol., 1962. Vol. 31, N 2. P. 317-329.
Yasumatsu K. A revision of the genus Sphex Linne of the Japanese empire (Hymenoptera, Sphecidae) // Tenthredo, 1938. Vol. 2. N 1. P. 43-135.
Zavadil V., Snoflak J. Kutilky (Sphecidae) Ceskoslovenske Republiky.
Vyskov, 1948. 179
s.
adelpha Kohl, Ammophila …………………………………158 afer Lep., Sphex ……………………………………………. 89 affinis (K.), Podalonia……………………………………….134 alpina (Kohl), Podalonia…………………………………….136 atrocyanea (Ev.), Podalonia…………………………………137 campestris Latr. , Ammophila ………………………………160 cellularis Guss. , Ammophila ..……………………………...162 deforme (Sm.), Sceliphron (Prosceliphron) …………………..82 dentigera Guss. , Ammophila………………………………..163
destillatorium (Ill.), Sceliphron (Sceliphron)………………….80
dives (Br.), Eremochares …………………………………….151
ebenina (Spin.), Podalonia…………………………………...139
elongata Fisch.-Waldh. , Ammophila………………………..164
femoratum (F.), Chalybion (Hemichalybion) ………………...75
fera (Lep.), Podalonia………………………………………...140
flavipennis F., Sphex ………………………………………….91
funerarius Guss., Sphex …….…………………………………93
gracillima Tasch. , Ammophila ..…………………………….166
haberhaueri (Rad.), Prionyx..…………………………………109
heydeni Dahlb. , Ammophila………………………………....168
hirsuta (Scop.), Podalonia…………………………………….142
hungarica Mocs. , Ammophila…………………………..……170
iliensis Kaz. , Ammophila………………………………..…..172
kirbyi (Lind.), Prionyx…………………………………….….110
lativalvis Guss. , Ammophila……………………………...….173
lividocinctus (Costa), Prionyx………………………………..113
luffi (Saund.), Podalonia……………………………………...144
macula (F.), Prionyx………………………………………….114 madraspatanum (F.),
Sceliphron (Sceliphron)…………………84 major Kaz., Dolichurus ……………………………………..…70
melanarius (Mocs.), Palmodes …………………………………97 minor (F.Mor.),
Palmodes……………………………………...99 mirabilis (Guss.), Eremochares…………………………..……153
nigropectinatus (Tasch.), Prionyx………………………….….116
niveatus (Duf.), Prionyx…………………………………….…117
nudatus (Kohl), Prionyx…………………………………….…119
occipitalis F.Mor. , Ammophila…………………………….…175
occitanicus (Lep. et Serv.) , Palmodes ………………………..101
orientalis (Mocs.), Palmodes………………………………..…103
pruinosus Germ., Sphex ..……..…………………………….…93
pubescens Curt. , Ammophila………………………………….176
pungens (Kohl), Podalonia……………………………………..146 radoszkowskyi
(Kohl), Prionyx …………………………..……121 rufocinctus Br., Sphex………….………………………….……94
sabulosa (L.) , Ammophila ……………………………………..178 sareptana Kohl , Ammophila
…………………………………...180
shestakovi Guss., Sceliphron (Prosceliphron) …………………..86
sinensis Sickm. , Ammophila..……………………………….…182
sirdariensis (Rad.), Prionyx……………………………………..123
songaricus (Ev.), Prionyx………………………………….…….123
strigulosus (Costa), Palmodes…………………………………...104
striata Mocs. , Ammophila………………………………………184
subfuscatus (Dahlb.), Prionyx ..…………………………………125
tekkensis Guss. , Ammophila……………………………………185
terminara F.Smith , Ammophila ..…………………………….…186
terminata terminata F.Smith, Ammophila………………………187 terminata
turanica Kazenas , Ammophila ……………………..188 terminata turkestana Kohl,
Ammophila.………………………..188
turanica F.Mor., Parapsammophila ……………………………131 turanicum
(Guss.), Chalybion (Chalybion)……………………..76 turanicus Guss. , Dolichurus
.…………………………………..71 tydei (Guill.), Podalonia…………………………….…………148
viduatus (Christ.), Prionyx………………………………….…127
СОДЕРЖАНИЕ
ОТ АВТОРА……………………………………………………………2
1. ВВЕДЕНИЕ………………………………………………………….3
2. ОБЩАЯ ХАРАКТЕРИСТИКА СЕМЕЙСТВА РОЮЩИХ ОС…9
2.1. Морфология имаго и преимагинальных стадий………………9
2.2. Общие особенности биологии…………………………………27
2.3. Систематика и филогения………………………………………47
3. ТАКСОНОМИЧЕСКИЙ СОСТАВ ФАУНЫ РОЮЩИХ ОС
КАЗАХСТАНА…………………………………………………….52
3.1. Определительная таблица подсемейств, триб и родов……….52
3.2. Подсем. Ampulicinae…………………………………………….67
3.3. Подсем. Sphecinae……………………………………………….72
4. ЗАКЛЮЧЕНИЕ…………………………………………………….191
ЛИТЕРАТУРА…………………………………………………………192
ИНДЕКС ЛАТИНСКИХ НАЗВАНИЙ ВИДОВ РОЮЩИХ ОС
ПОДСЕМЕЙСТВ AMPULICINAE И SPHECINAE КАЗАХСТАНА
………………………………………………………………….……….213
ИЛЛЮСТРАЦИИ……………………………………………………...217
ИЛЛЮСТРАЦИИ

Рис. 1. Dolichurus turanicus Guss., ♂
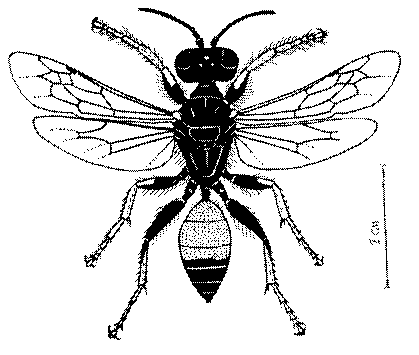
Рис. 2. Sphex rufocinctus Br., ♀ (по Lomholdt, 1975)

Рис. 3. Ammophila sabulosa L., ♀ (по Lomholdt, 1975)

Рис. 4. Astata sp., ♀
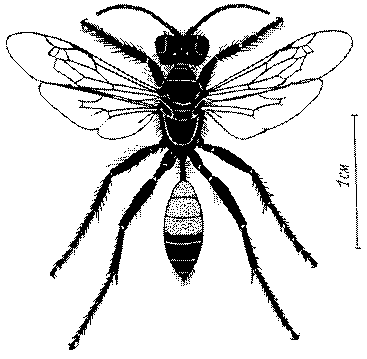
Рис. 5. Podalonia hirsuta (Scop.), ♀ (по Lomholdt, 1975)
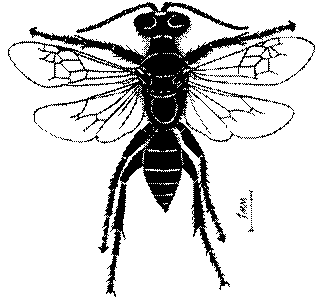
Рис. 6. Diploplectron pulawskii Kazenas, ♂
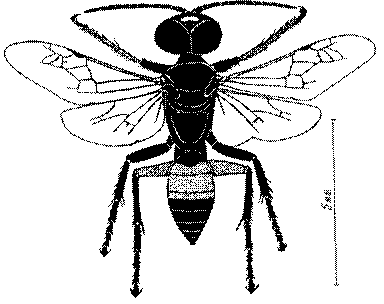
Рис. 7. Dryudella tricolor (Lind.), ♂

Рис. 8. Dinetus pictus (F.), ♀ (по Lomholdt, 1975)
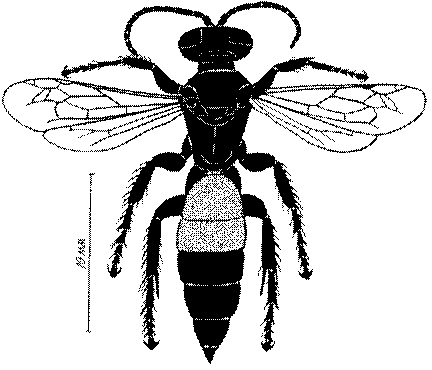
Рис. 9. Larra anathema (Rossi), ♀

Рис. 10. Parapiagetia tridentata Ts., ♀

Рис. 11. Pison sogdianum Guss., ♀

Рис. 12. Представители подсем. Crabroninae: a) Palarus variegatus
(F.), ♀; б) Tachysphex panzeri (Lind.), ♀; в) Trypoxylon attenuatum Sm.,♀

Рис. 13. Miscophus sp., ♀

Рис. 14. Nitela borealis Valk., ♀ (по Lomholdt, 1975)

Рис. 15. Belomicrus borealis , ♀ (по Lomholdt, 1975)

Рис. 16. Oxybelus argentatus (по Grandi, 1961)
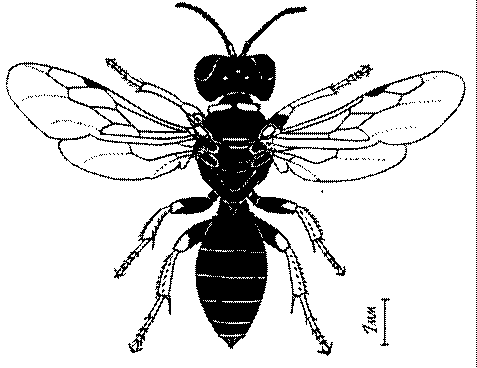
Рис. 17. Lindenius albilabris (F.), ♀

Рис. 18. Entomognathus brevis (Lind.), ♀

Рис. 19. Crossocerus elongatulus Lind., ♀

Рис. 20. Rhopalum coarctatum Scop., ♀

Рис. 21. Crabro cribrarius (L.), ♀

Рис. 22. Ectemnius cavifrons (Thoms.), ♀
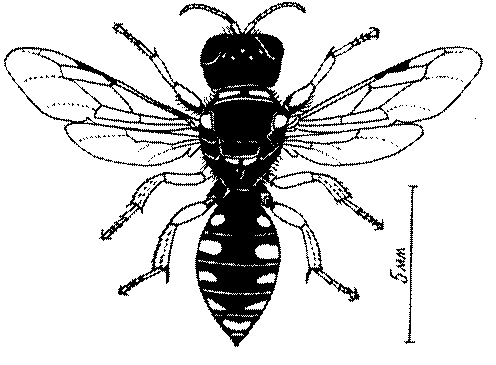
Рис. 23. Lestica subterranea (F.), ♀

Рис. 24. Entomosericus kaufmanni Rad., ♀
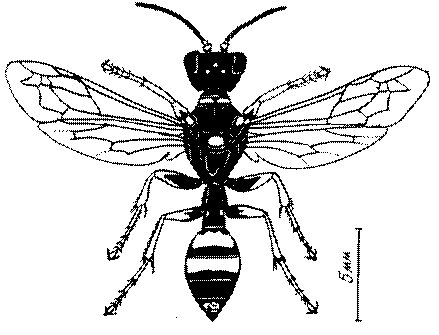
Рис. 25. Mellinus arvensis (L.), ♀

Рис. 26. Nysson spinosus (J.Forst.), ♀ (по Lomholdt, 1975)

Рис. 27. Olgia modesta Rad., ♀

Рис. 28. Argogorytes mystaceus L., ♀ (по Lomholdt, 1975)
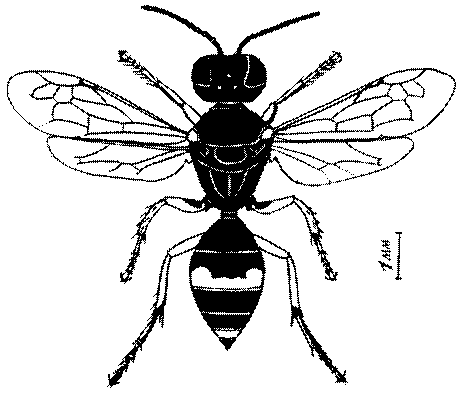
Рис. 29. Harpactus tumidus (Panz.), ♀

Рис. 30. Gorytes quadrifasciatus (F.), ♀

Рис. 31. Ammatomus coarctatus Spin., ♂
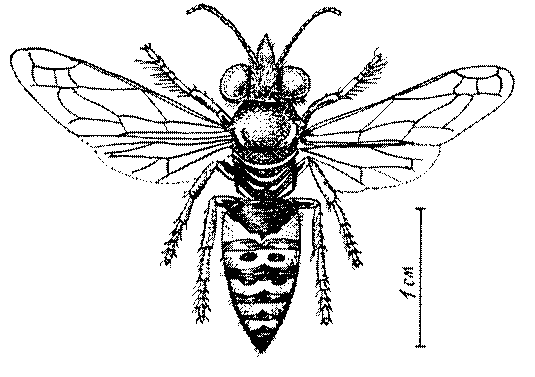
Рис. 32. Bembix oculata Panz., ♀ ( по Grandi, 1961)

Рис. 33. Eremiasphecium bicolor Guss., ♀
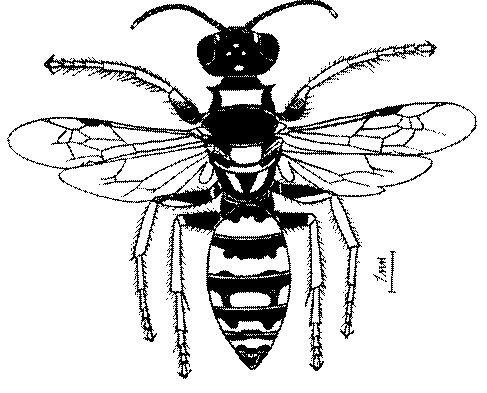
Рис. 34. Pseudoscolia simplicicornis F.Mor., ♀

Рис. 35. Philanthus triangulum (F.), ♀
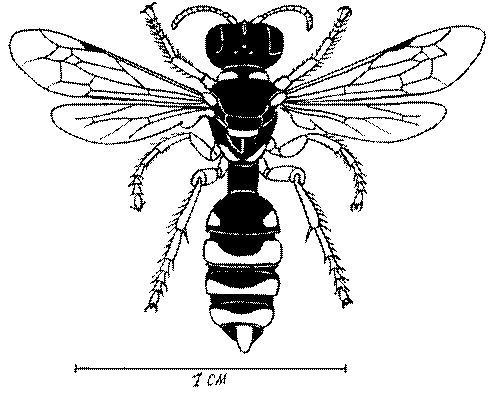
Рис. 36. Cerceris dorsalis Ev., ♀
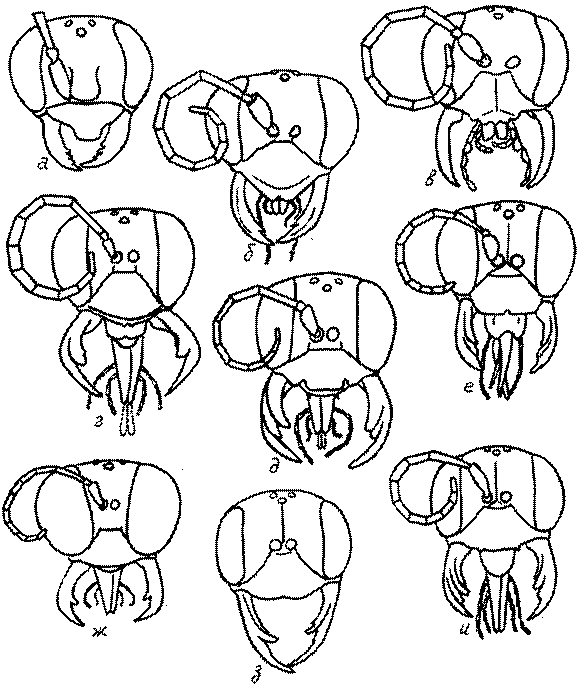
Рис. 37. Голова представителей Ampulicinae и Sphecinae: а – Dolichurus sp., б – Sceliphron sp., в – Chalybion sp., г – Sphex sp., д – Palmodes sp., e – Prionyx sp., ж – Eremochares sp., з – Podalonia sp., и – Ammophila sp.; а-и - ♀♀
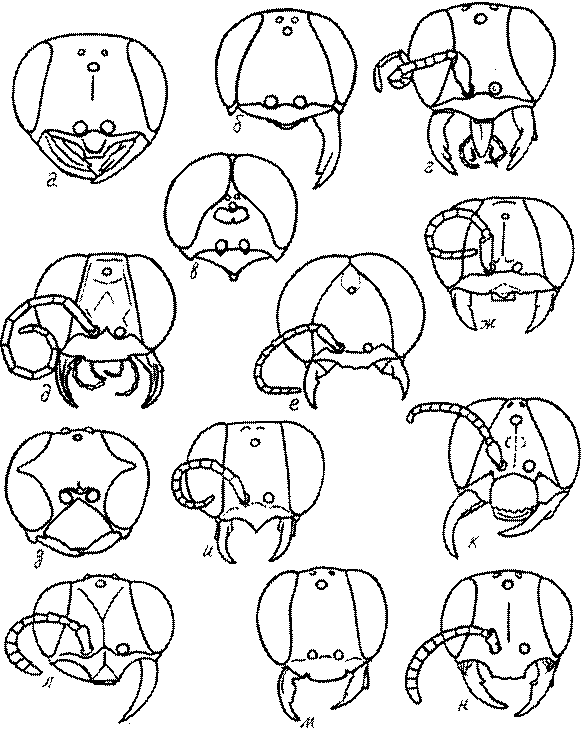
Рис. 38. Голова Astatinae и Crabroninae: a – Astata boops; б,в – Dryudella stigma; г – Dinetus sp., д – Liris sp., е - Prosopigastra sp., ж - Parapiagetia sp., з - Trypoxylon sp., и - Gastrosericus waltlii, к - Palarus sp., л - Solierella sp., м - Miscophus sp., н - Plenoculus sp., а,б,г,д,ж,з,к,л,м - ♀♀; в,е,и,н - ♂♂
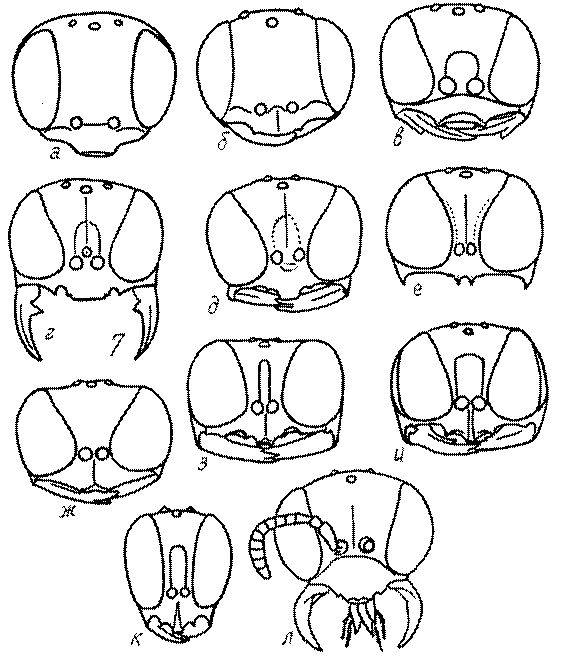
Рис. 39. Голова Crabroninae, Entomosericus: a – Belomicrus antennatus, б – Oxybelus bipunctatus, в – Entomognathus brevis, г – Lindenius panzeri, д – Rhopalum gracile, е – Crossocerus annulipes, ж – Crabro cribrarius, з – Ectemnius cavifrons, и – Lestica alata, к – L. clypeata, л – Entomosericus sp. ; а-и, л - ♀♀, к - ♂
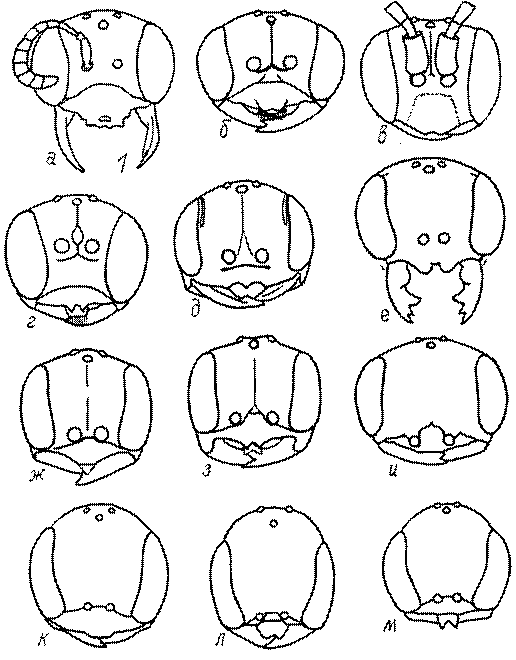
Рис. 40. Голова Pemphredoninae: а – Mimesa sp., б – Mimumesa dahlbomi, в – Psen ater, г – Psenulus sp., д – Diodontus minutus, е – Pemphredon lethifer, ж – Passaloecus gracilis, з – Stigmus solskyi, и – Spilomena sp., к – Ammoplanops carinatus, л – Ammoplanus transcaspicus, м – Ammoplanellus sp.;
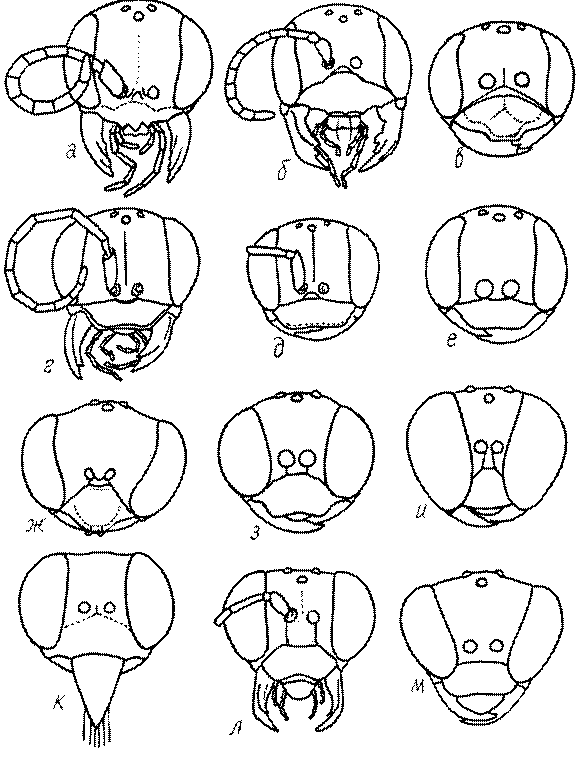
Рис. 41. Голова Nyssoninae: а – Alysson sp., , - Mellinus sp., в – Argogorytes sp., г – Lestiphorus sp., д – Oryttus sp., е – Harpactus tumidus, ж – Nysson spinosus, з – Gorytes quinquefasciatus, и – Ammatomus coarctatus, к – Bembix rostrata, л – Stizus ruficornis, м – Bembecinus tridens; а-м - ♀♀
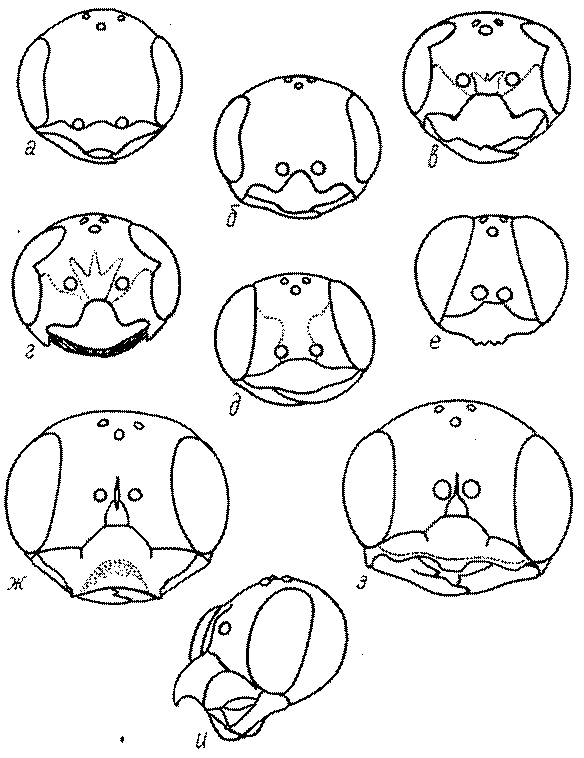
Рис. 42. Голова Philanthinae: а – Eremiasphecium crassicorne, б – Philanthinus albiceps, в,г – Philanthus triangulum, д,е – Pseudoscolia spp., ж – Cerceris edolata, з – C. tinnula, и – C. ruficornis; а,в,д-и - ♀♀, б,г - ♂♂

Рис. 43. Голова Sphecidae сверху (а-г), сбоку (д) и глазки (е-м). а – Pemphredon lugubris, б – Passaloecus gracilis, в – Crossocerus capitosus, г –
Lestica clypeata, д – Pemphredon inornatus; а-д - ♀♀; е – Larra anathema, ж – Gastrosericus funereus, з – Tachysphex panzeri, и – Tachytes obsoletus, к – Palarus variegatus, л – Parapiagetia tridentata, м – Bembix sp.
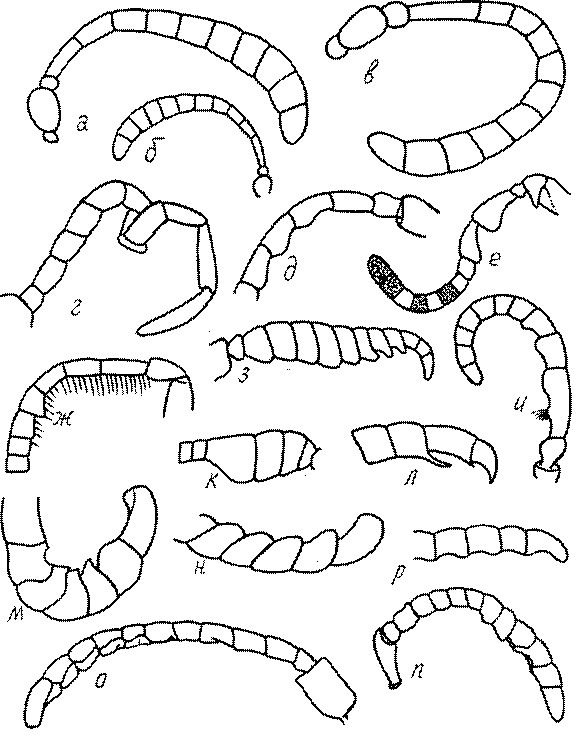
Рис. 44. Усики Sphecidae: а – Mimesa pulawskii, б - Ammatomus coarcttus, в - Philanthus marikovskii, г - Dinetus pictus, д - Prosopigastra rufiventris, е - Rhopalum coarctatum, ж - Crossocerus denticoxa, з - Crabro cribrarius, и - Ectemnius sexcinctus, к - E. crassicornis, л - Bembecinus tridens, м - Bembix turca, н - Pseudoscolia diversicornis, о - Psen ater, п - Hoplisoides sp., р - Harpactus affinis; a- ♀; б-р - ♂♂.
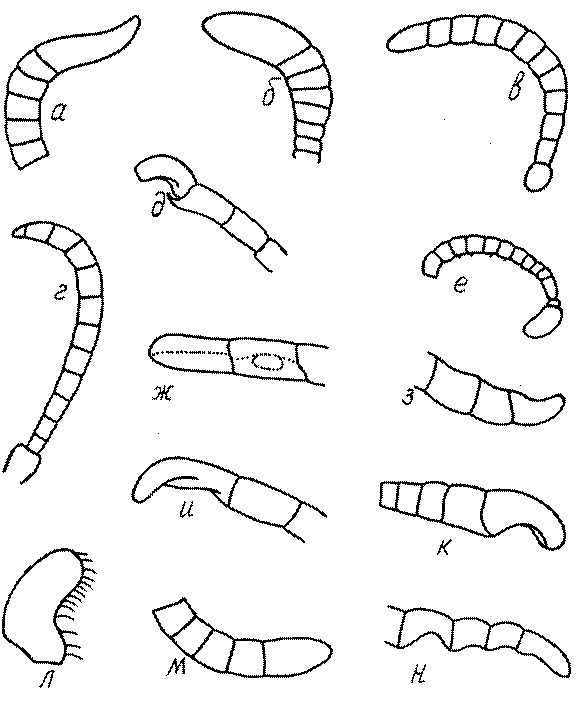
Рис. 45. Усики и вершинная часть усиков Sphecidae: а – Trypoxylon attenuatum, б – T. kolazyi, в – Plenoculus murgabensis, г – Solierella compedita, д – Didineis sp., е – Entomosericus sp., ж – Oryttus cribratus, з – Psammaecius luxuriosus, и – Sphecius antennatus, к – Nysson maculatus, л – Cerceris antilope, м – Pseudoscolia simplicicornis, н – Harpactus elegans; а-н -
♂♂
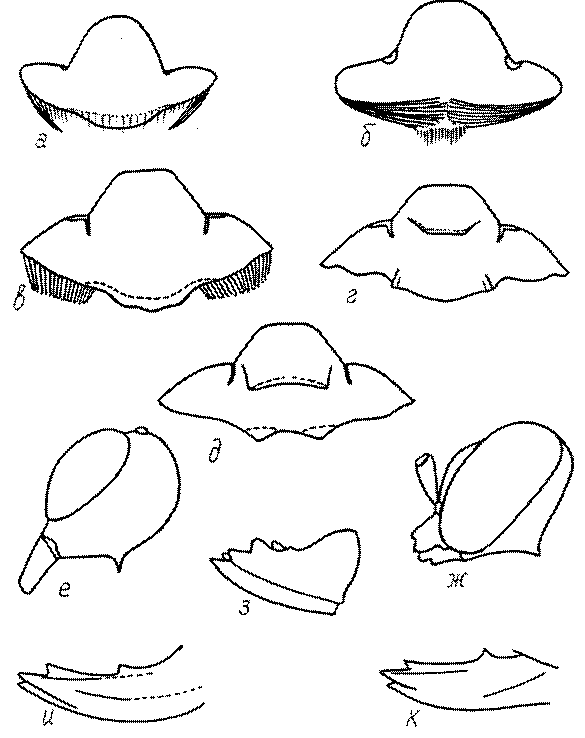
Рис. 46. Детали строения головы Sphecidae: а – Philanthus coronatus, б – Ph. venustus, в – Cerceris turkestanica, г – C. bracteata, д – C. lavilabris, е – Lindenius panzeri, ж – Gastrosericus sp., з – Pemphredon inornatus, и – Crossocerus dimidiatus, к – Ectemnius continuus; ф-д – наличник, е,ж – голова сбоку, з-к – мандибула; а-в,е - ♂♂; г,д,ж-к - ♀♀
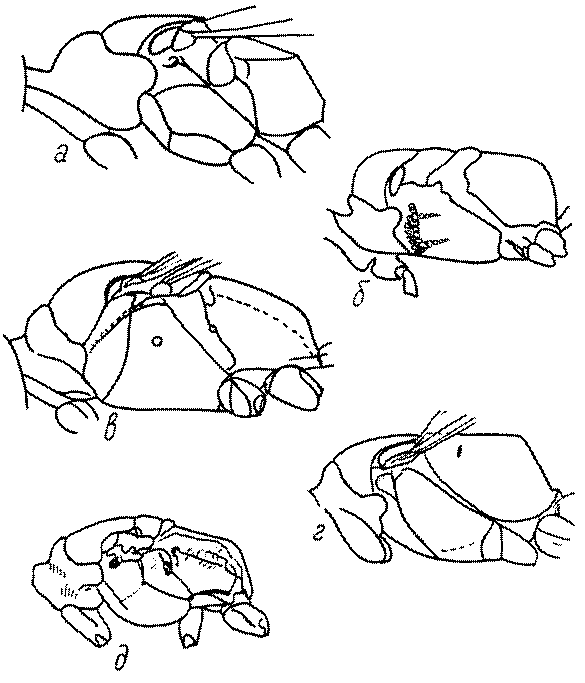
Рис. 47. Грудь Sphecidae сбоку: а – Dolichurus turanicus, б – Passaloecus gracilis, в – Trypoxylon sp., г – Liris nigra, д – Alysson sp.
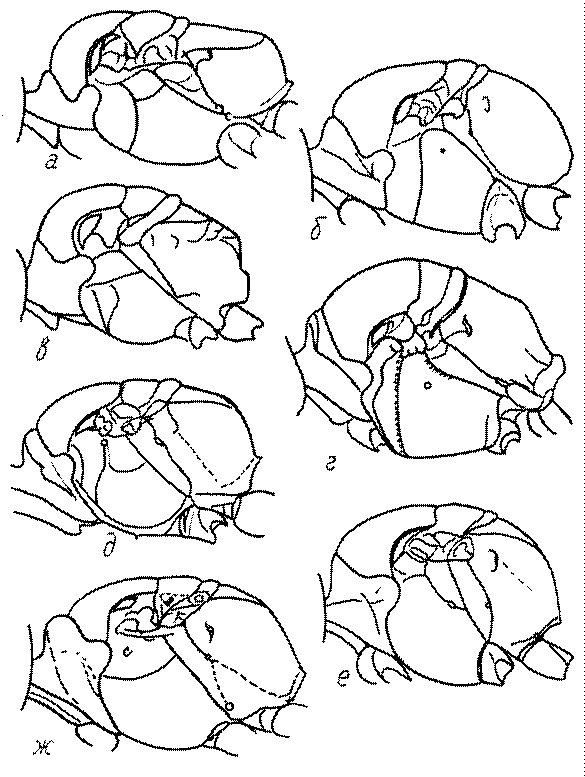
Рис. 48. Грудь Sphecidae сбоку: a – Astatsa boops, б – Tachytes obsoletus, и – Diodontus minutus, г – Lindenius sp., д – Gorytes sp., е – Stizus ruficornis, ж – Cerceris sp.
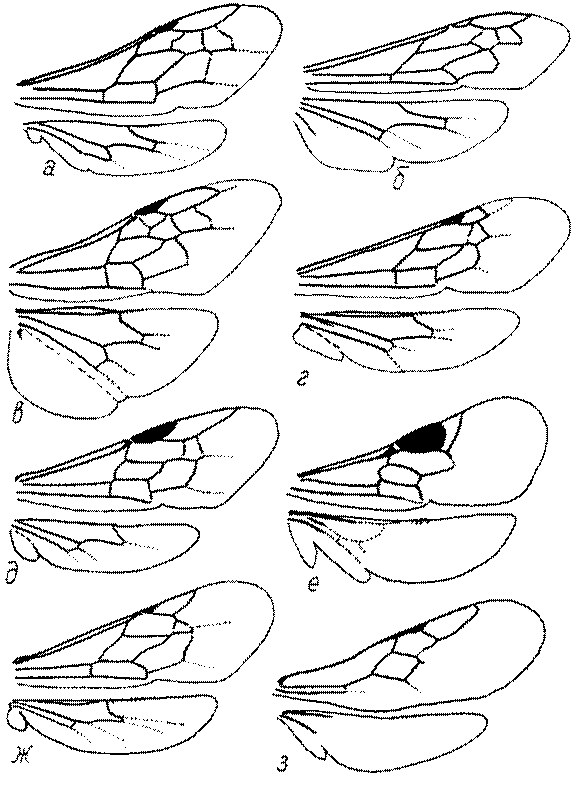
Рис. 49. Жилкование крыльев Sphecidae: а – Dolichurus sp., б – Chalybion sp., в – Astata sp., г – Dinetus pictus, д – Diodontus tristis, е – Ammoplanus sp., ж – Miscophus sp., з – Nitela sp.
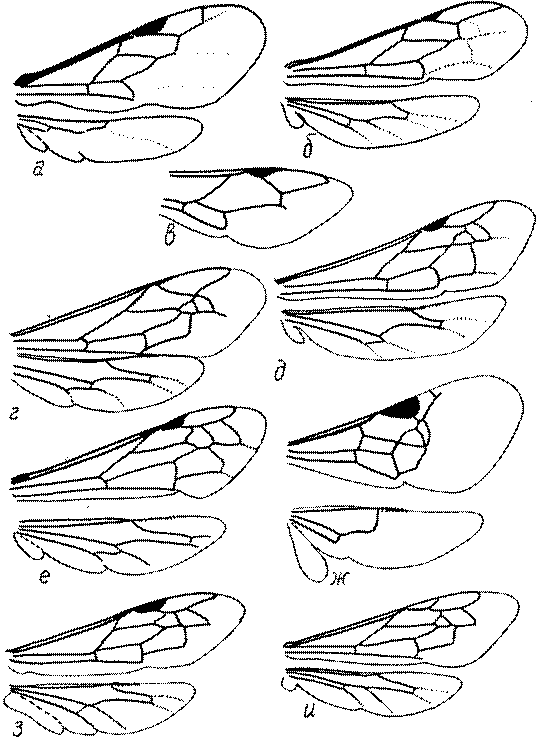
Рис. 50. Жилкование крыльев Sphecidae: а – Entomognathus brevis, б – Oxybelus sp., в – Trypoxylon sp., г – Nysson sp., д – Didineis sp., е – Gorytes sp., ж – Eremiasphecium sp., з – Philanthus coronatus, и – Cerceris rybyensis.
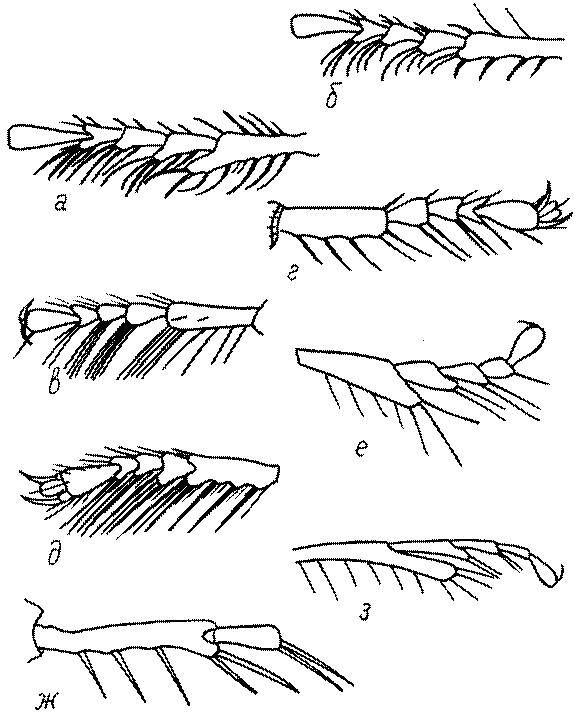
Рис. 51. Передние лапки самок Sphecidae: а – Parapsammophila sp., б – Eremochares dives, в – Tachysphex panzeri, г – Gastrosericus funereus, д – Bembix turca, е – Eremiasphecium crassicornis, ж – Philanthus marikovskii, з – Eremiasphecium digitatum.
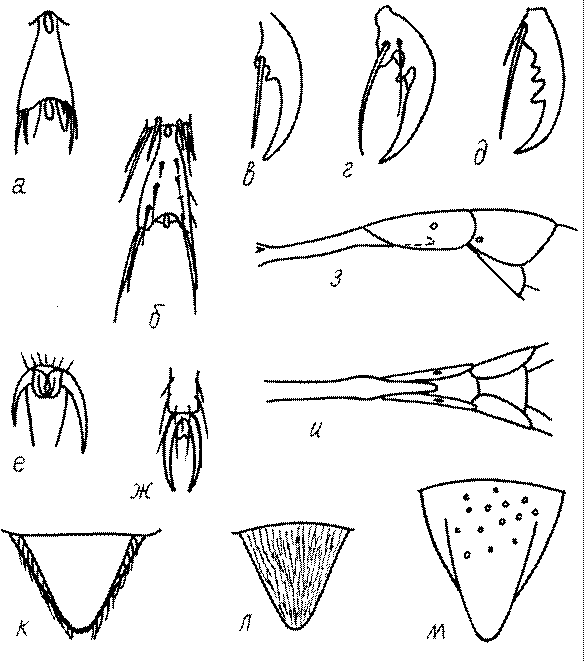
Рис. 52. Детали строения лапок (а-ж) и брюшка (з-м) Sphecidae: а – Sceliphron sp., б – Parapsammophila turanica, в – Eremochares dives, г – Sphex flavipennis, д – Prionyx viduatus, е – Bembix sp., ж – Podalonia affinis, з,и – Ammophila sp., к – Astata rufipes, л – Tachytes bidens, м – Gastrosericus funereus; а,б – 3-й членик снизу; в-д – коготок лапок; е,ж – вершина лапок; з – базальная часть брюшка сбоку; и – то же снизу; к-м – пигидиальное поле самок.
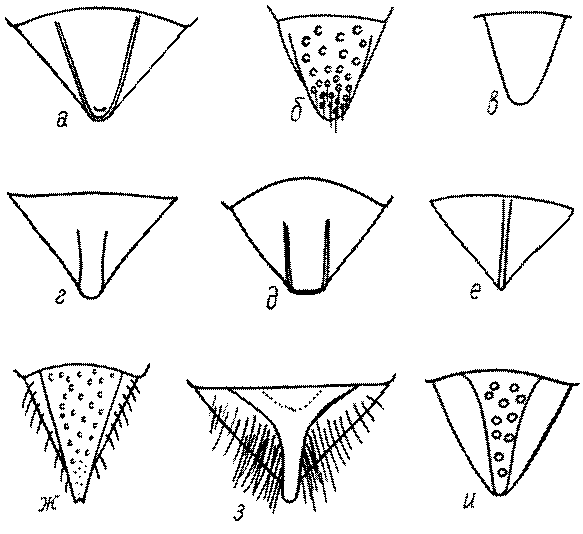
Рис. 53. Пигидий ♀♀ Sphecidae: а – Psenulus laevis, б – Psen ater, в – Diodontus minutus, г – Pemphredon sp., д – P. inornatus, е – P. lugens, ж – Tachysphex panzeri, з – Lestica clypeata, и – Ectemnius sp.
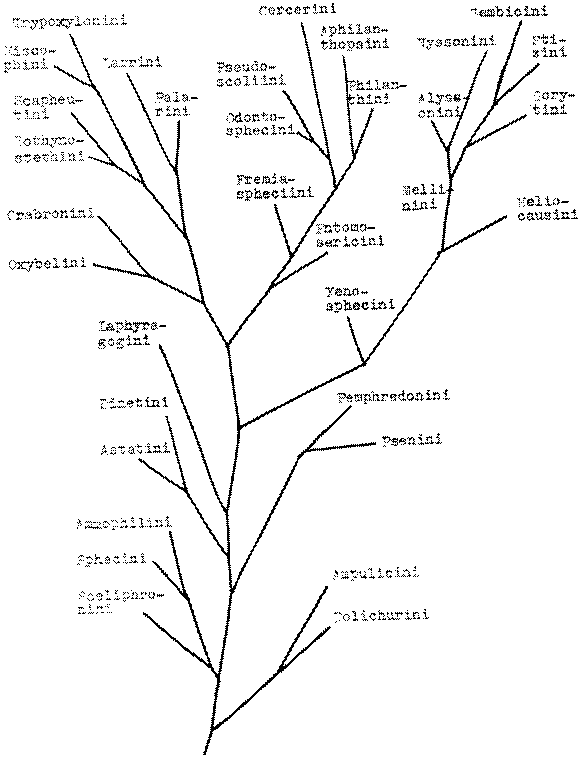
Рис. 54. Схема филогенетических отношений триб роющих ос
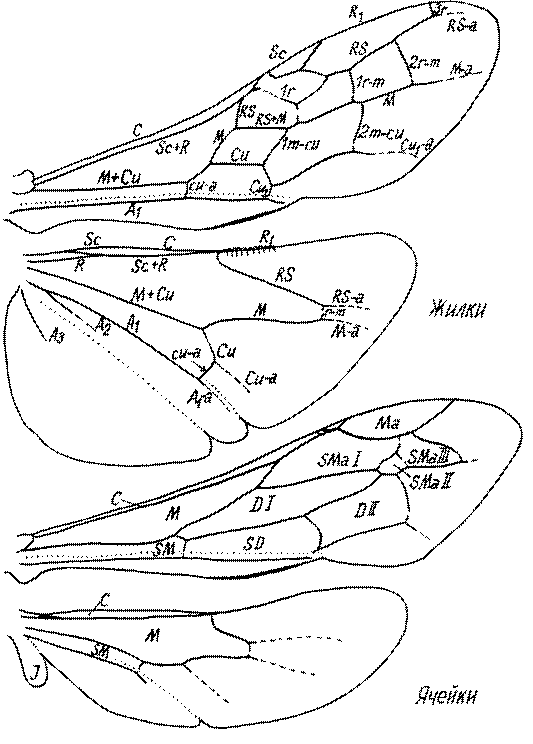
Рис. 55.Жилкование крыльев Sphecidae. Условные обозначения: ячейки (по Bohart and Menke, 1976): С – костальная; DI, DII – 1-я и 2-я дискоидальные; J – югальная; М – медиальная; Ма – маргинальная; SM – субмедиальная; SmaI, SmaII, SMaIII – 1-я, 2-я и 3-я субмаргинальные; SD – субдискоидальная; жилки (по Тобиасу, 1978): A – анальная; А-а, Cu-a, M-a, RS-a – апикальный отрезок жилок A, Cu, M, RS; С – костальная; Cu – кубитальная; cu-a – нервулюс (на переднем крыле) или нервеллюс (на заднем крыле); М – медиальная; M+C – базальная (медио-кубитальная продольная); m-cu – возвратная, или медио-кубитальная поперечная; R – радиальная; RI – метакарп; r – поперечная радиальная; r-m – радио-медиальная; RS – радиальный сектор; Sc – интеркостальная; Sc+R – субкостальная
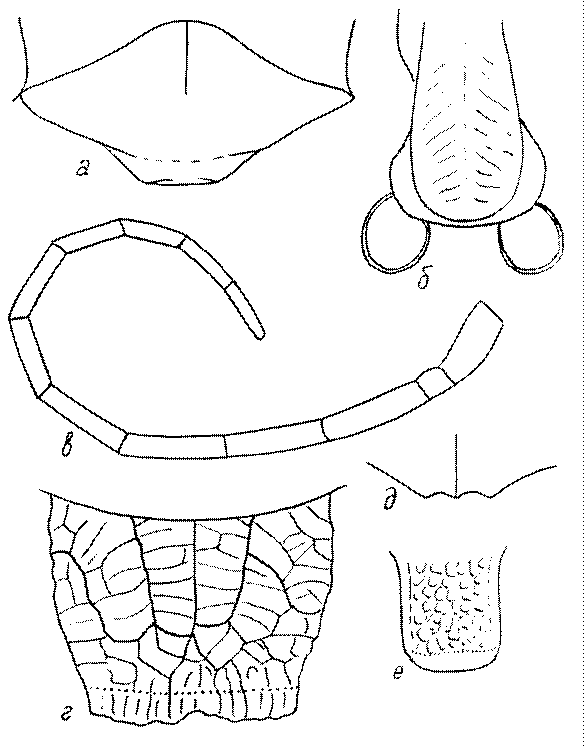
Рис. 56. Dolichurus major, самка (а-г) и D. turanicus, самец (д,е): а – наличник, б,е – лобный выступ, в – усик, г – промежуточный сегмент сверху, д – передний край наличника
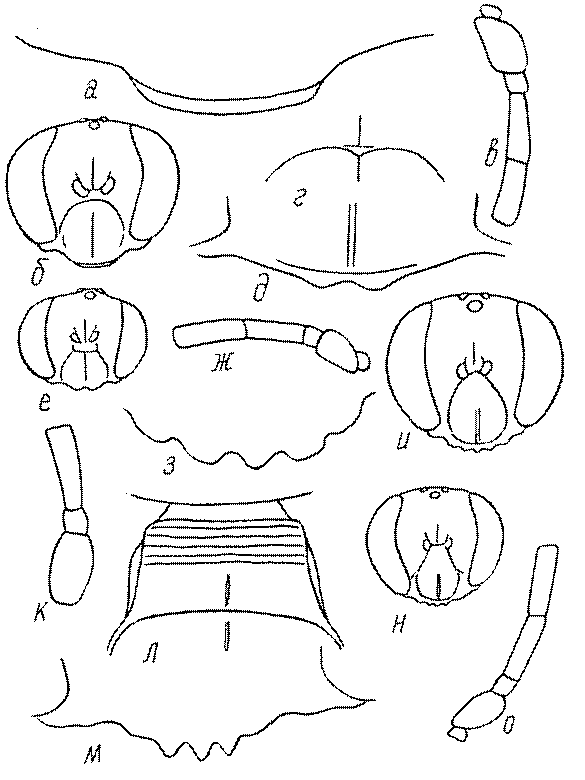
Рис. 57. Chalybion femoratum (а-ж) и Ch. turanicum (з-о): ф,д,з,м – передний край наличника; б,е,и,н – голова спереди; в,ж,к,о – базальные членики усика; г – переднеспинка (вид спереди); л – переднеспинка сверху; а-г, з-л - ♀♀; д-ж, м-о - ♂♂.
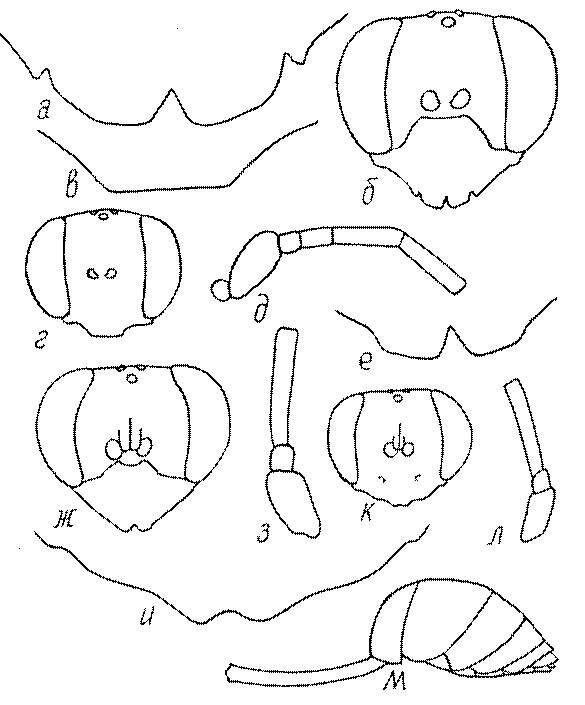
Рис. 58. Sceliphron destillatorium (а-д) и S. deforme (е-м): a,в,е,и – передний край наличника; б,г,ж,к – голова спереди; д,з,л – базальные членики усика; м – брюшко сбоку; а-в, е-з - ♀♀; г,д,и-м - ♂♂
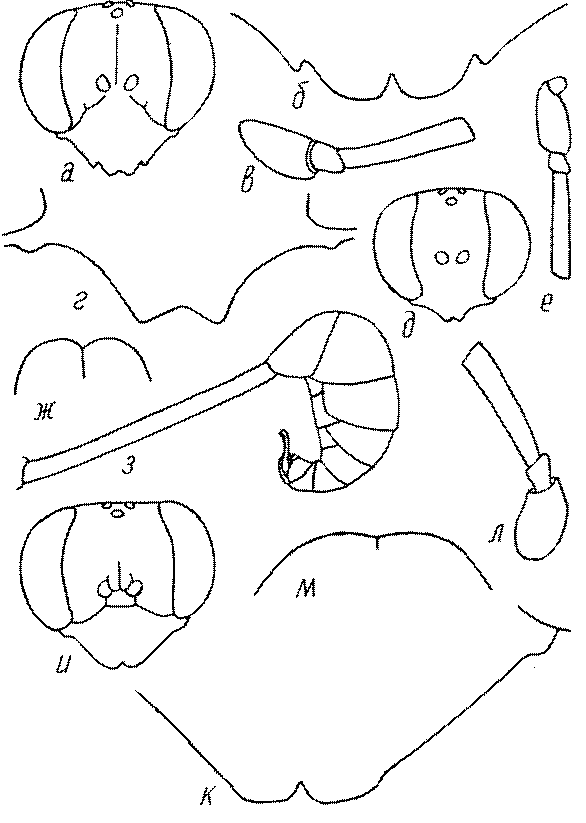
Рис. 59. Sceliphron madraspatanum (а-з) и S. shestakovi (и-м): а,д,и – голова спереди; б,г,к – передний край наличника; в,е,л – базальные членики усиков; ж,м – переднеспинка спереди; з – брюшко; а-в - ♀♀; г-з - ♂♂
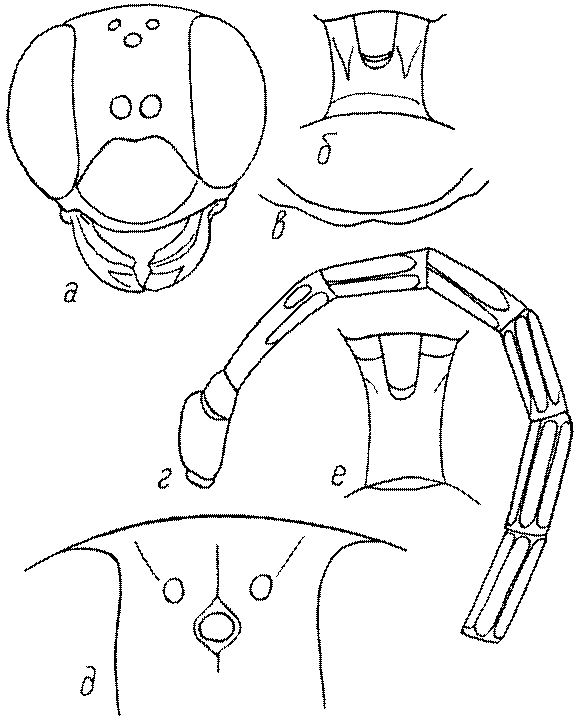
Рис. 60. Sphex afer: а – голова спереди, б,е – стебелек брюшка, в – передний край наличника, г – усик, д – темя; а,б - ♀; в-е - ♂.
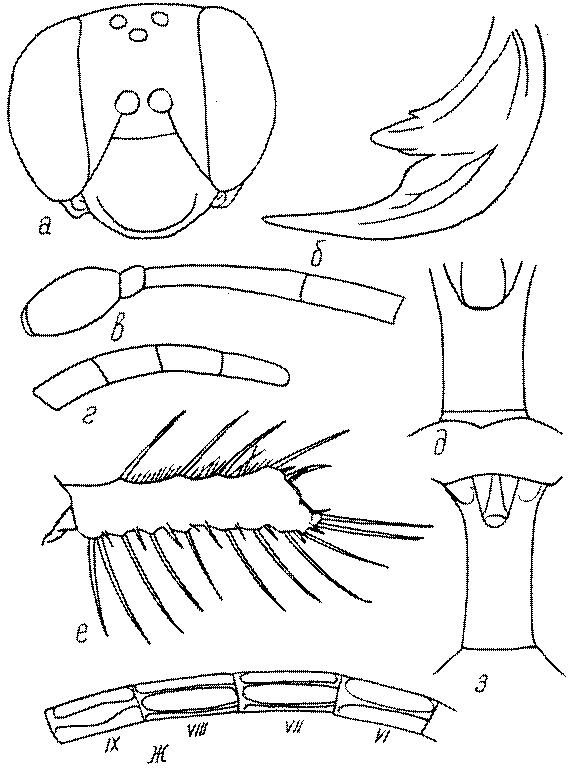
Рис. 61. Sphex flavipennis: а – голова спереди, б – мандибула, в – базальная часть усика, г – вершинная часть усика, д,з – стебелек брюшка сверху, е – 1-й членик передних лапок, ж – средняя часть усика; а-е - ♀; ж,з - ♂
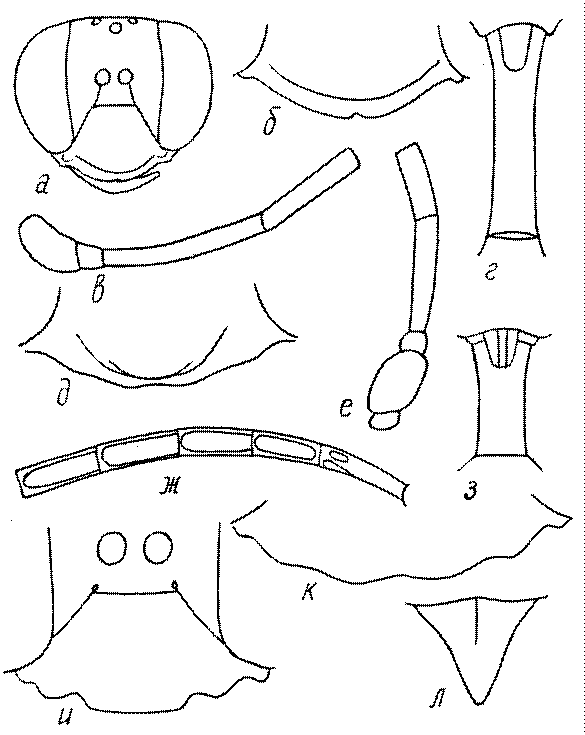
Рис. 62. Sphex pruinosus (а-г), S. rufocinctus (д-з) и Palmodes melanarius (и-л): ф – голова спереди; б,к – передний край наличника; в,е – базальная часть усика; г,з – стебелек брюшка сверху; ж – базальная часть жгутика усика; и – наличник и нижняя часть лба; л – 8-й стернум; а-е,и - ♀; ж,з,л - ♂.
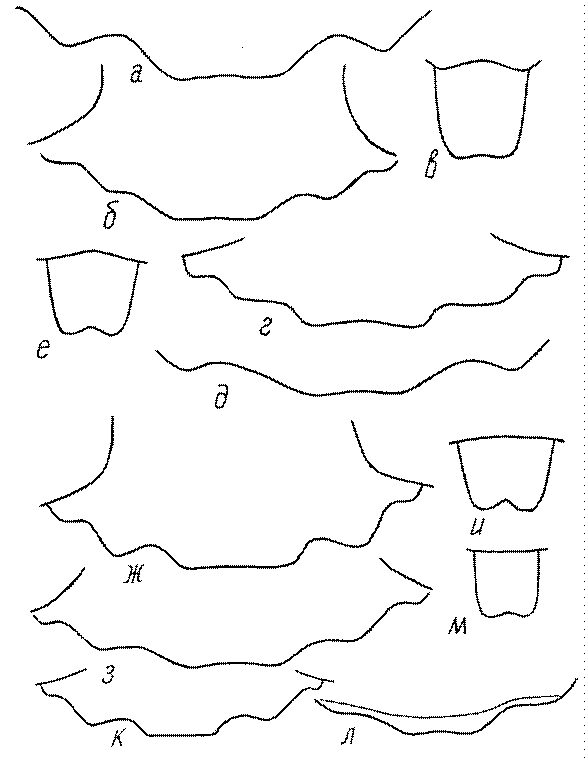
Рис. 63. Palmodes minor (а-в), P. occitanicus (г-е), P. orientalis (ж-и), P. strigulosus (к-м): а,б,г,д,ж,з,к,л – передний край наличника; в,е,и,м – 8й стернум; а,г,ж,к - ♀; б,в,д,е,з,и,л,м - ♂.
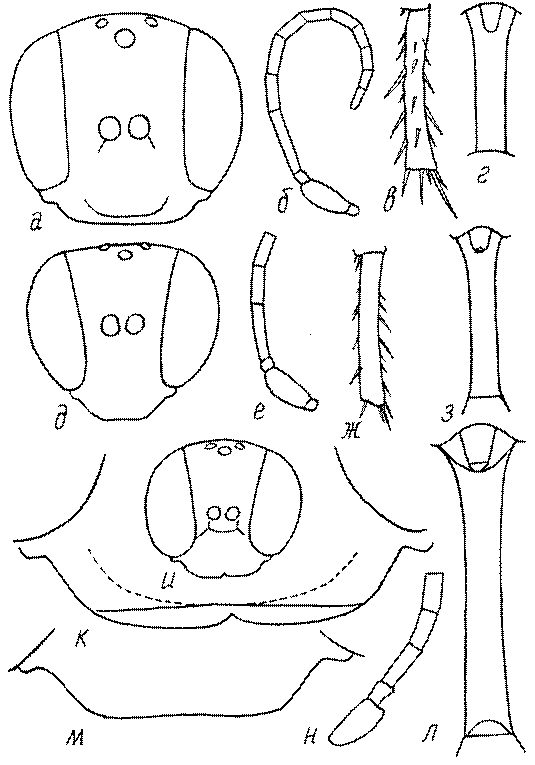
Рис. 64. Prionyx haberhaueri (а-з) и P. kirbyi (и-н): а,д,и – голова спереди; б – усик; в,ж – 1-й членик задней лапки; г,з,л – стебелек брюшка; е,н – базальные членики усика; к,м – передний край наличника; а-г, и-л - ♀; д-з, м, н - ♂.
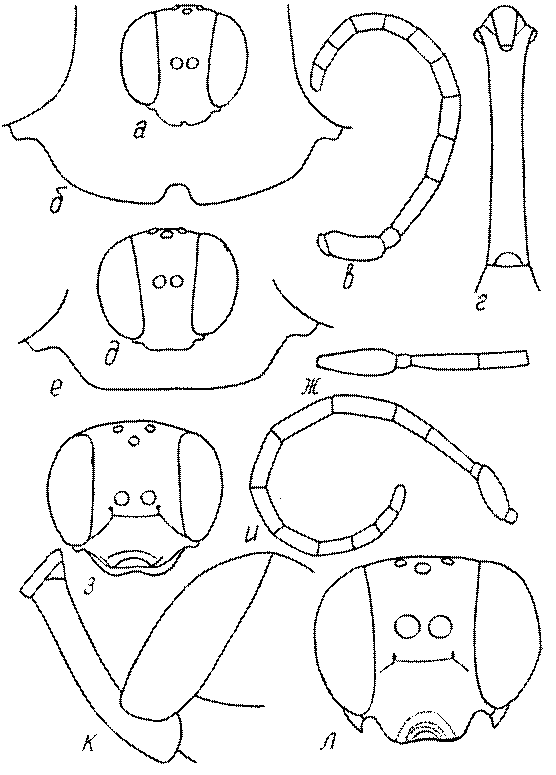
Рис. 65. Prionyx lividocinctus (а-ж) и P. macula (з-л): а,д,з,л – голова спереди; б,е – передний край наличника; в,и – усик; г – стебедек брюшка сверху; ж – базальные членики усика; к – основание брюшка сбоку; а-г, и-к - ♀; д-ж, л - ♂.
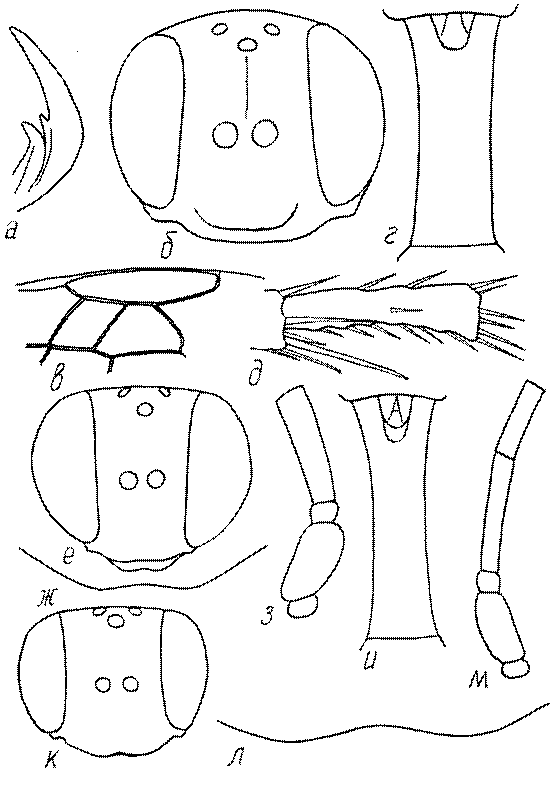
Рис. 66. Prionyx nigropectinatus (а-и) и P. niveatus (к-м): а – мандибула; б,е,к – голова спереди; в – часть переднего крыла; г,и – стебелек брюшка; д – 1-й членик задней лапки; ж,л – передний край наличника; зм – базальная часть усика; а-д, к-м - ♀; е-и - ♂.
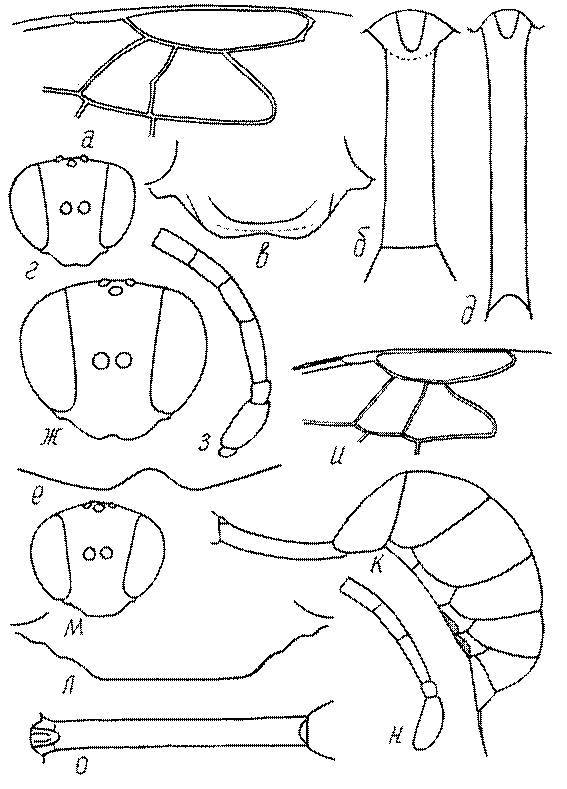
Рис. 67. Prionyx niveatus (а-д), P. nudatus (е-о): а,и – часть переднего крыла; б,д,о – стебелек брюшка; в – наличник; г,ж,м – голова спереди; е,л – передний край наличника; з,н – базальная часть усика; к – брюшко сбоку.
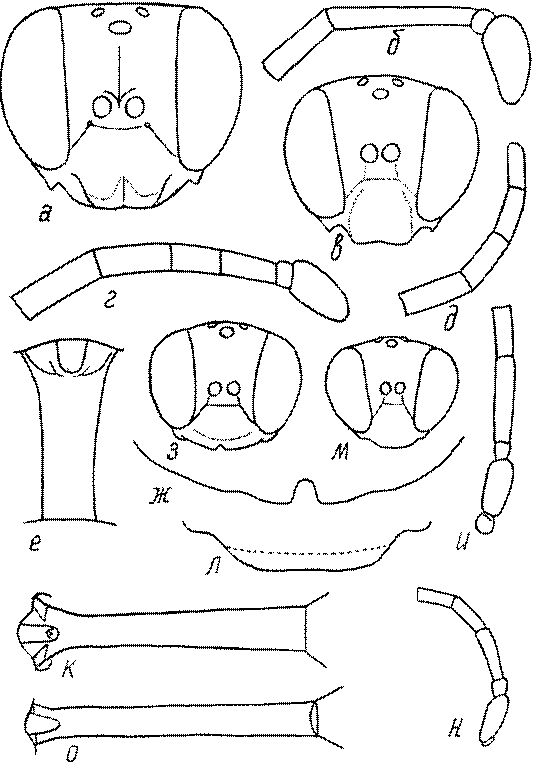
Рис. 68. Prionyx radoszkowskyi (а-е) и P. songaricus (ж-о): а,в,з,м – голова спереди; б,г,и,н – базальная часть усика; д – вершинная часть усика; е,к,о – стебелек брюшка сверху; ж,л – передний край наличника; а,б,ж-к - ♀; в-е, л-п - ♂.
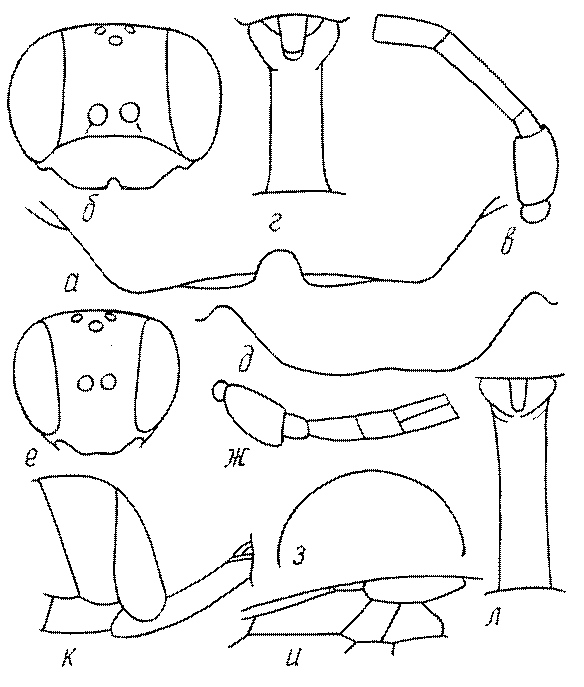
Рис. 69. Prionyx subfuscatus: а,д – передний край наличника; б,е – голова спереди; в,ж – базальная часть усика; г,л – стебелек брюшка сверху; з – переднеспинка спереди; и – часть переднего крыла; к – базальная часть брюшка сбоку; а-г - ♀; д-л - ♂.
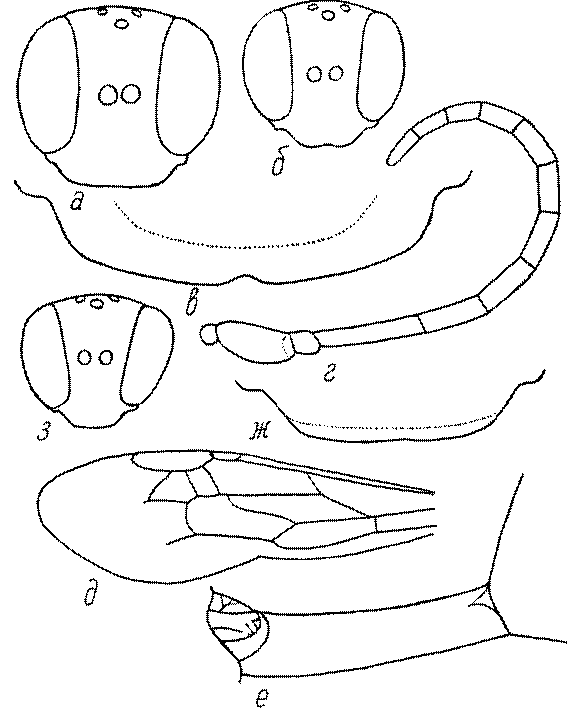
Рис. 70. Prionyx viduatus: а,б,з – голова спереди; в,ж – передний край наличника; г – усик; д – переднее крыло; е – стебелек брюшка (косо сверху); а-е - ♀; ж,з - ♂.
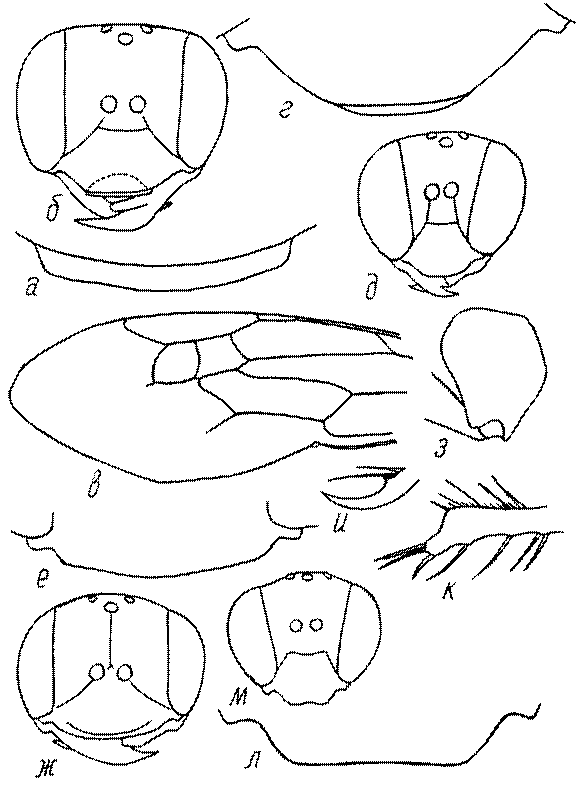
Рис. 71. Parapsammophila turanica (а-д), Podalonia affinis (е-м): а,г,е,л – передний край наличника; б,д,ж,м – голова спереди; в – переднее крыло; з – передний тазик; и – коготок лапок; к – 1-й членик передней лапки.
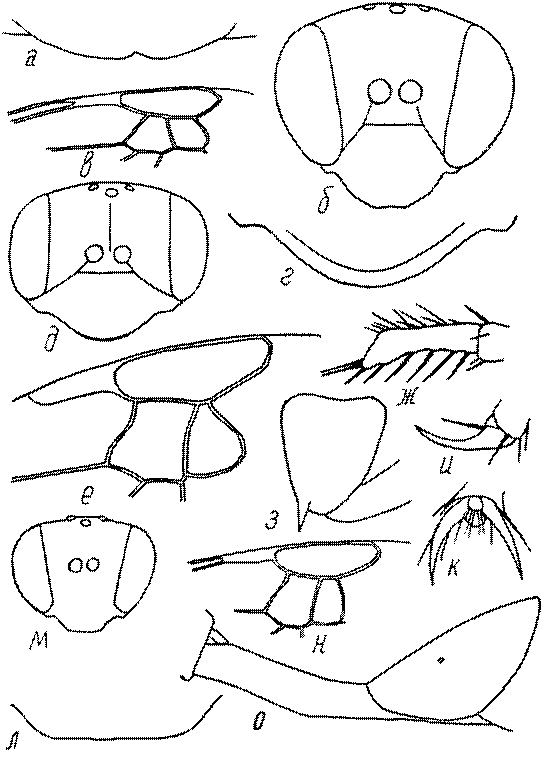
Рис. 72. Podalonia alpina (а-в) и P. atrocyanea (г-о): а,г,л – передний край наличника; б,д,м – голова спереди; в,е,н – часть переднего крыла; ж – 1-й членик передних лапок; з – передний тазик; и – коготок лапок; к – вершина лапок; о – базальная часть брюшка сбоку; а,г-к - ♀; б,в,л-о - ♂.
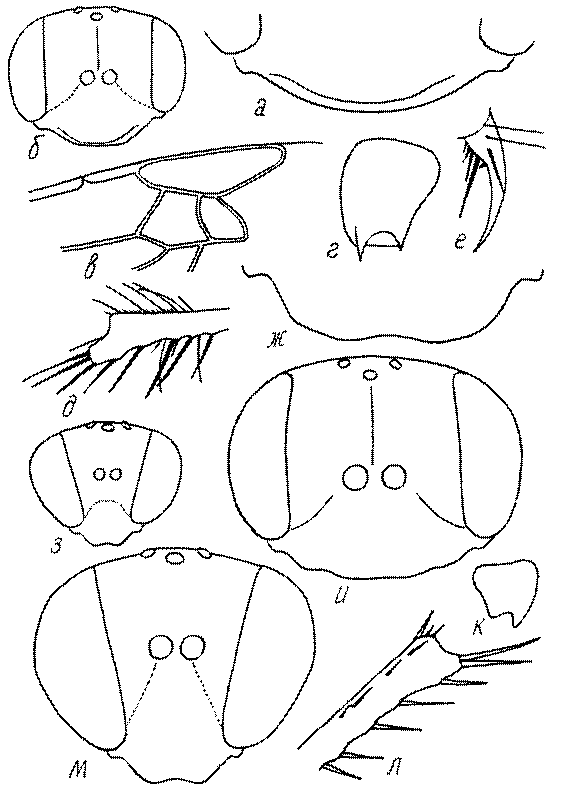
Рис. 73. Podalonia ebenina (а-з), P. fera (и-м): а,ж – передний край наличника; б,з,и,м – голова спереди; в – часть переднего крыла; г,к – передний тазик; д,л – 1-й членик передних лапок; е – коготок лапок; а-д,ил - ♀; ж,з,м - ♂.
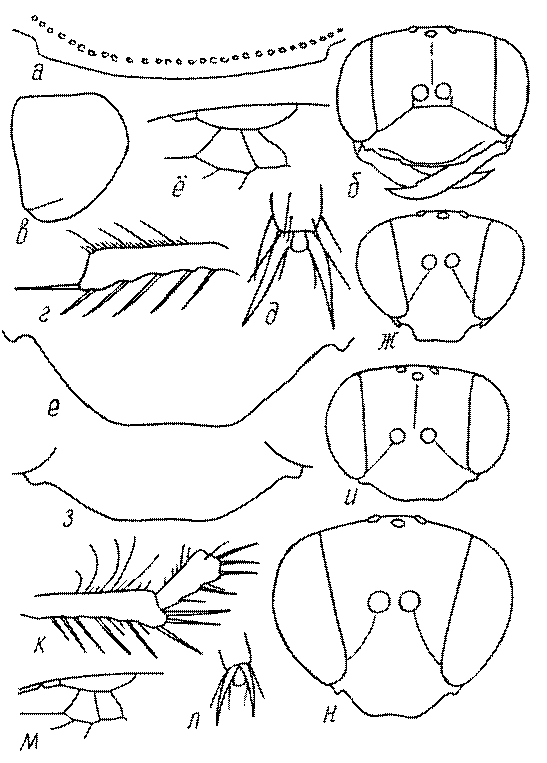
Рис. 74. Podalonia hirsuta (а-ж), P. luffi (и-н): а,е,з – передний край наличника; б,ж,и,н – голова спереди; в – передний тазик снизу; г – 1-й членик передних лапок; д,л – вершинный членик передних лапок; ё,м – часть переднего крыла; к – 1-й и 2-й членики передних лапок; а-е,з-м - ♀; е,ж,н - ♂.
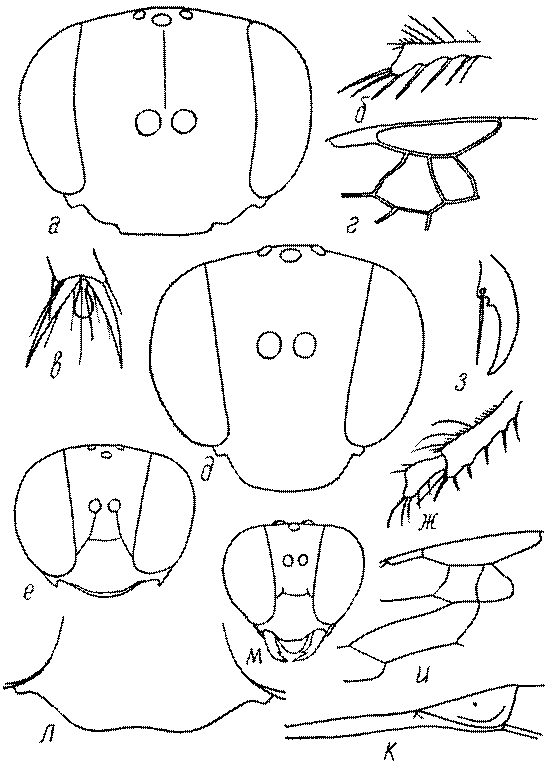
Рис. 75. Podalonia tydei (а-д) и Eremochares dives (е-м): а,д,е,м – голова спереди; б – 1-й членик передних лапок; в – вершина лапок; г,и – часть переднего крыла; ж – 1-й и 2-й членики передних лапок; з – коготок лапок; к – базальная часть брюшка сбоку; л – передний край наличника; а-г, е-к - ♀; д,л,м - ♂.
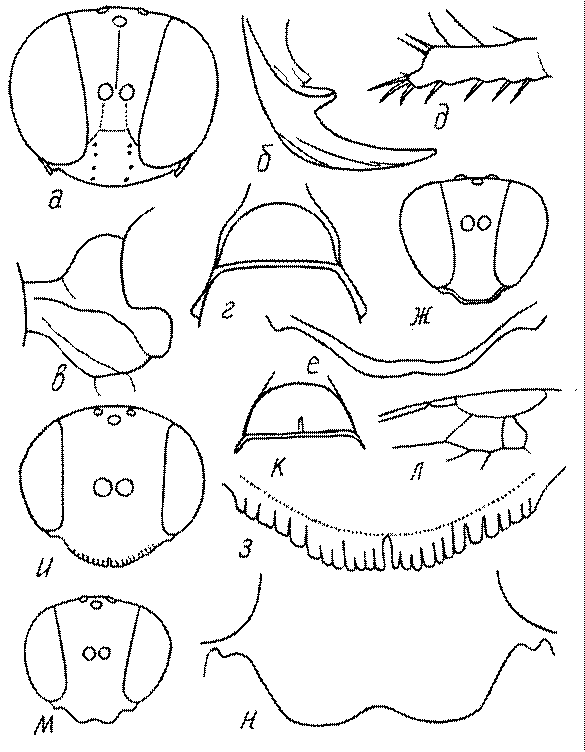
Рис. 76. Eremochares mirabilis (а-ж) и Ammophila adelpha (з-н): а,ж,и,м – голова спереди; б – мандибула; в,к – переднегрудь сбоку; г – переднеспинка сверху; д – 1-й членик передних лапок; е,з,н – передний край наличника; л – часть переднего крыла; а-д,з-л - ♀; е,ж,м,н - ♂.
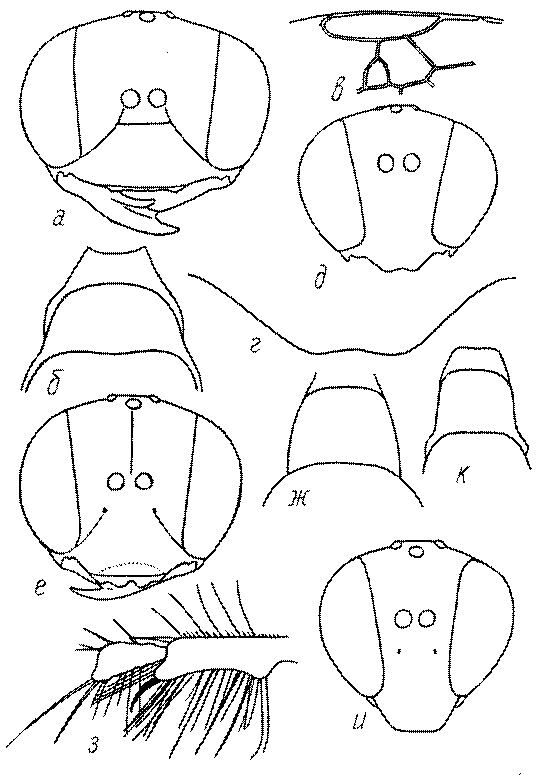
Рис. 77. Ammophila campestris (а-д) и A. dentigera (е-к); а,д,е,и – голова спереди; б,ж,к – переднеспинка сверху; в – часть переднего крыла; г – передний край наличника; з – 1-й и 2-й членики передних лапок; а-в, е-з - ♀; г,д,и,к - ♂.
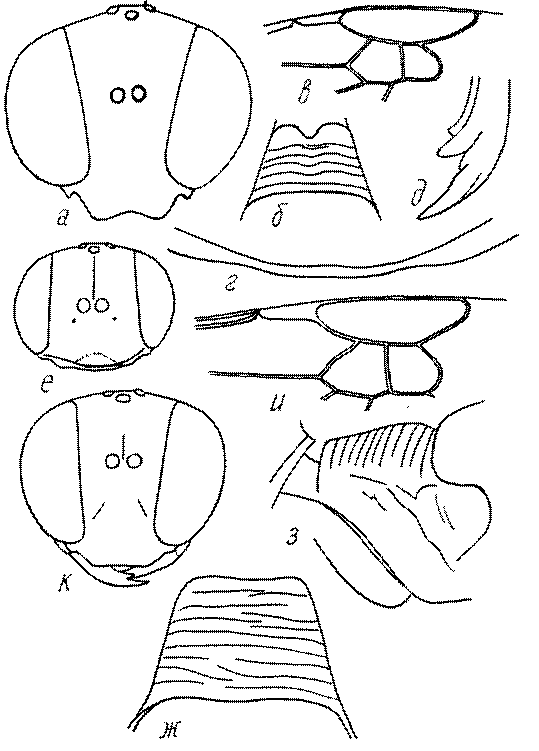
Рис. 78. Ammophila elongata (а-в) и A. gracillima (г-к): а,е,к – голова спереди; б,ж – воротничок переднеспинки сперху; в,и – часть переднего крыла; г – передний край наличник; д – мандибула; з – переднегрудь сбоку.
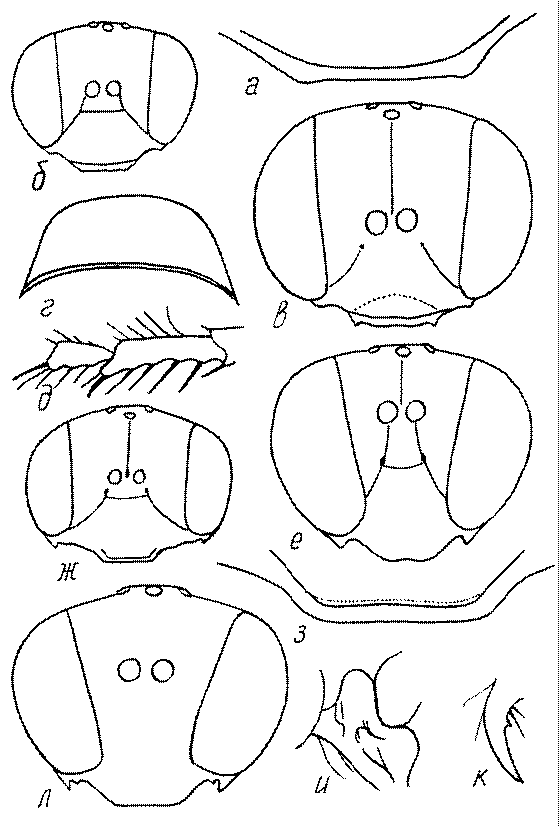
Рис. 79. Ammophila heydeni (а-е) и A. hungarica (ж-л): а-з – передний край наличника; б,в,е,ж,л – голова спереди; г - валик переднеспинки сверху; д – базальные членики передней лапки; и – переднегрудь сбоку; к – коготок лапок; а-д,ж-к - ♀; е,л - ♂.
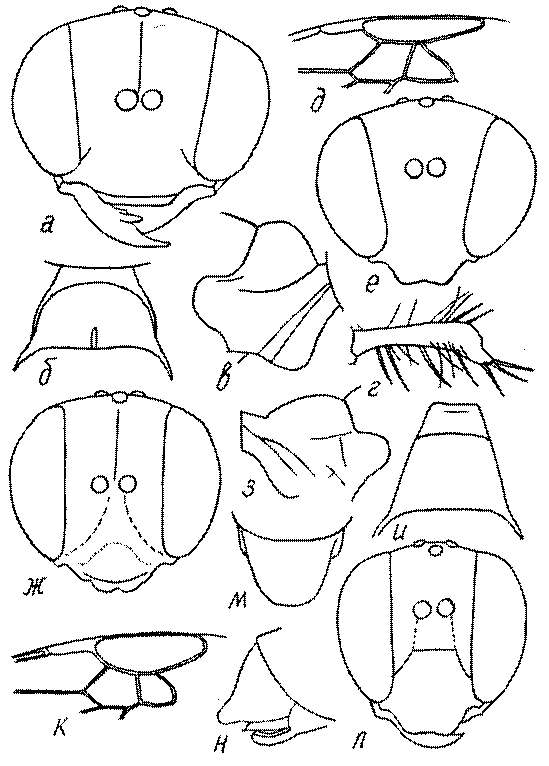
Рис. 80. Ammophila iliensis (а-е) и A. lativalvis (ж-н): а,е,ж,л – голова спереди; б,и – переднеспинка сверху; в,з – переднегрудь сбоку; г – 1-й членик передних лапок; д,к – часть переднего крыла; м – 7-й тергум сверху; н – вершина брюшка сбоку; а-д,ж-к - ♀; е,м,н - ♂.
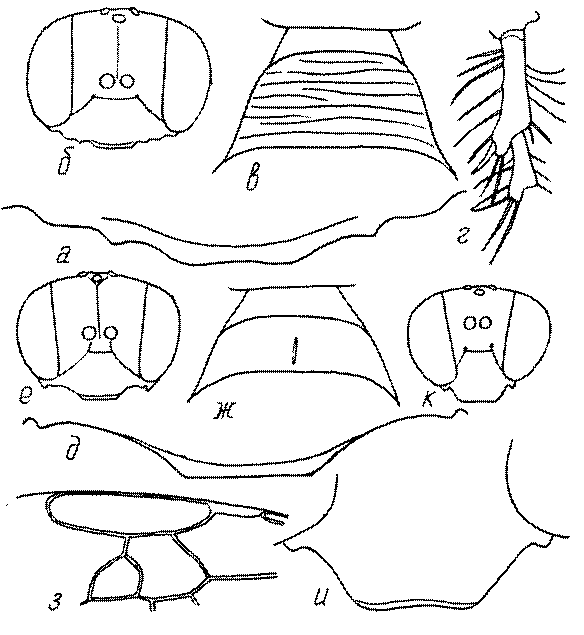
Рис. 81. Ammophila occipitalis (а-г) и A. pubescens (д-к): а,д,и – передний край наличника; б,е,к – голова спереди; в,ж – переднеспинка сверху; г – 1-й и 2-й членики передних лапок; з – часть переднего крыла; а-з - ♀ и; ,к - ♂.
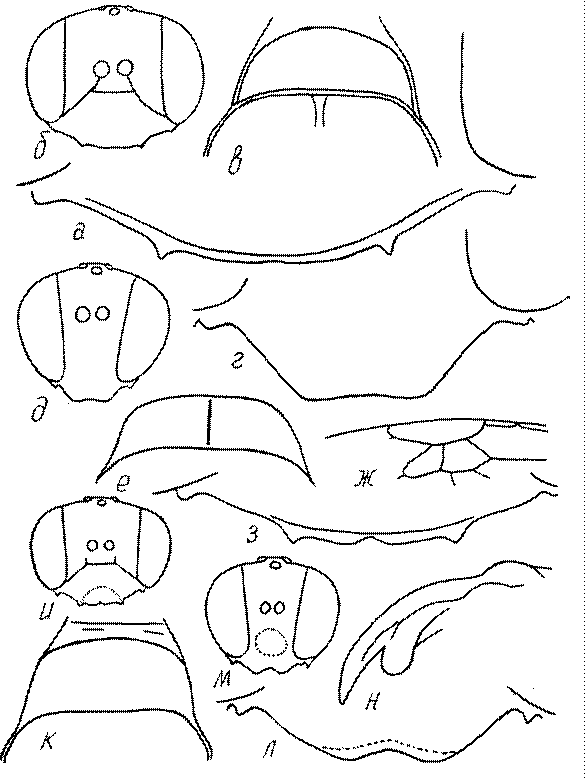
Рис. 82. Ammophila sabulosa (а-ж), A. sareptana (з-н): а,г,з,л – передний край наличника; б,д,и,м – голова спереди; в,е,к – переднеспинка сверху; ж – часть переднего крыла; н – мандибула; а-в,з-к - ♀; г-ж,л-н - ♂.
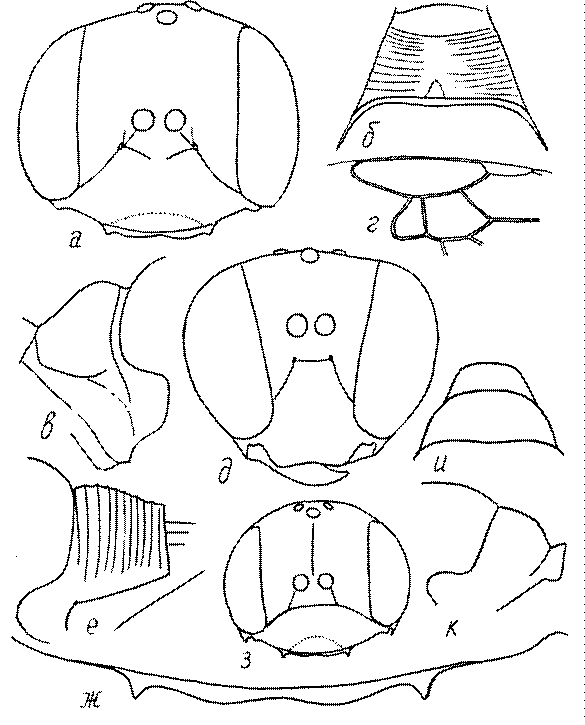
Рис. 83. Ammophila sinensis (а-е), A. striata (ж-к): а,д,з – голова спереди; б,и – переднеспинка сверху; в,к – переднеспинка сбоку; г – часть переднего крыла; ж – передний край наличника; а-г,ж-к - ♀; д,е -♂.
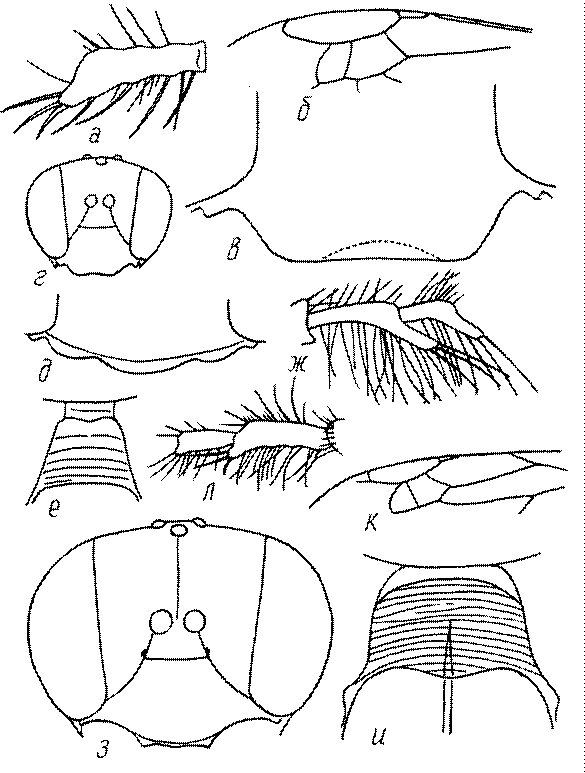
Рис. 84. Ammophila striata (а-г), A. tekkensis (д-ж), A. terminata terminata (з-л): а – 1-й членик передних лапок; б,к – часть переднего крыла; в,д – передний край наличника; г,з – голова спереди; е,и – переднеспинка сверху; ж,л – 1-й и 2-й членики передних лапок; а,б,д-л - ♀; в,г - ♂.
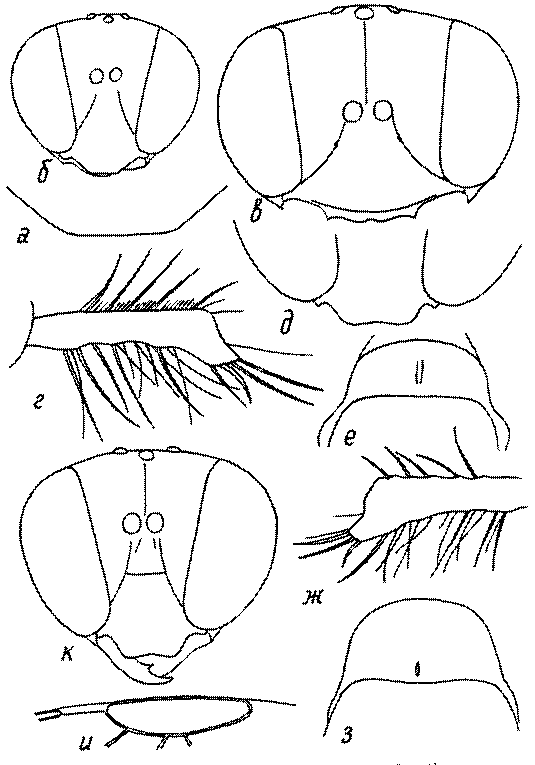
Рис. 85. Ammophila terminata terminata (а,б), A. t. turanica (в-е), A. t. turkestana (ж-к): а,д – передний край наличника; б,в,к – голова спереди; г,ж – 1-й членик передней лапки; е,з – переднеспинка сверху; и – маргинальная ячейка переднего крыла; а,б,д,е,к - ♂; в,г,ж-и - ♀.
Казенас В.Л. Роющие осы (Hymenoptera, Sphecidae) Казахстана. (Подсем. Ampulicinae, Sphecinae). Выпуск 1. Алматы, 1998. 377с. (Деп. в КазгосИНТИ
7.04.1998, № 8239 Ка98.)


















































Материалы на данной страницы взяты из открытых источников либо размещены пользователем в соответствии с договором-офертой сайта. Вы можете сообщить о нарушении.
