

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по дисциплине "Медико-биологические основы обучения и воспитания детей с ограниченными возможностями здоровья" предназначены для студентов 2 курса СПО педагогического профиля, обучающихся по специальности "Специальное дошкольное образование", преподавателей, ведущих данную дисциплину, а также практикующих педагогов-дефектологов, воспитателей детских садов общеразвивающего вида
Основы генетики. Теория.docx
Предметом генетики является изучение закономерностей и механизмов, а также структуры и
функции реализации генетического материала и генетической информации. Т.о. генетика изучает два
фундаментальных свойств живого:
наследственность – свойство живых организмов передавать в ходе размножения особенности
структур и функций
изменчивость – это свойство живых систем существовать в различных состояниях, переходя из
одного состояния в другое.
Главным методом генетики является изучение процесса наследования, т.е. процессов передачи
генетической информации от родителей к потомкам, включая результаты этой передачи.
Наследственность связана с работами Грегора Менделя, который впервые сформулировал законы
наследования – основу дискретной теории наследственности.
Методы работы Менделя
1. впервые вводит т.н. гибридологический метод исследования, который подразумевает скрещивание
разных форм в нужном состоянии
2. выбор объекта. Горох – это самоопыляемое растение с большим количеством сортов. Для
различных сортов характерны альтернативные признаки. Признаки передаются из поколения в
поколение. Самоопыляемость позволяет получить чистые линии. Горох дает большое количество
семян, т.е. большая выборка исследуемого материала. Искусственным путем горох можно опылить
перекрестно, т.е. использовать гибридологический метод.
3. Мендель впервые ввел в науку статистические методы обработки. Эти методы подразумевают
анализ большого количества исследуемого материала, поэтому горох был достаточно удобным
объектом
4. повторение эксперимента
5. Мендель впервые использует методику рецепрокных скрещивании. Это скрещивание родителей
противоположных по признакам, которое дают одинаковые результаты.
6. Мендель впервые использует т.н. возвратное или бэккросс скрещивание. Частным случаем бэк
кросс скрещивания является анализирующее скрещивание, когда организм с неизвестным
генотипом скрещивают с гомозиготой по рецессиву.
Основные термины
o Ген – это участок молекулы ДНК (РНК), который несет информацию о специфической структуре
или функции.
o Гены локализованы в хромосомах. Место положения гена называется локусом
o Для многоклеточных характерен диплоидный набор хромосом. Он практически одинаков у
представителей разных полов и представлен парами гомологичных хромосом, однако по одной паре
хромосом они различаются. Эта пара получила название половых хромосом. Все остальные
хромосомы получили название аутосомными.
o Конкретной формой существования гена является аллель, т.е. аллель это конкретная
последовательность нуклеотидов, находящаяся в данном локусе.
o Аллель, который контролирует признак, всегда проявляющийся у потомства, называется
доминантным и, следовательно, такой признак тоже будет являться доминантным. Аллель, который
контролирует признак, необязательно проявляющийся у потомства, называется рецессивным
o Если в одинаковых локусах гомологичных хромосом располагается только один вариант аллелей,
то такой организм называется гомозиготой. Если в одних и тех же локусах располагаются разные
варианты аллелей, то такой организм называется гетерозиготой.
А… — доминанта а — рецессив
АА
Гомозигота по доминанте
Аа
гетерозигота
аа
Гомозигота по рецессиву
Генотип – это совокупность всех или анализируемых аллелей характерных для данного организма
(генотип – это индивидуальная характеристика)
Фенотип – это совокупность всех или анализируемых признаков, которые характерны для данного
организма.
Законы Менделя
Закон единообразия первого поколения (Первый закон Менделя). Определение: при
скрещивании двух родителей различающихся по одному признаку (гомозиготы) все потомство будет
единообразно. Моногибридными являются скрещивания двух организмов, которые различаются по
аллелям одного гена. Современная формулировка: при скрещивании двух гомозигот все потомки
будут единообразны по фенотипу и генотипу.
Доказательство:
1. при полном доминировании. Полное доминирование характеризуется тем, что у потомков
проявляется признак только одного из родителей. Докажем, исследуя форму семян у гороха.
Дано:
А гладкие
аа морщинистые
Доказательство:
Р: АА +аа
F: Аа
2. доказательство при промежуточном наследовании (неполное доминирование). Промежуточное
наследование характеризуется тем, что у потомков проявляются признаки обоих родителей.
Докажем на наследовании окраски цветков у "ночной красавицы". У белых цветков имеются
крупные межклетники, которые заполнены воздухом, а при скрещивании с красными цветками
оттенок становится розовым.
Дано:
А красные
аа белый
Доказательство:
Р: АА +аа
F: Аа
3. кодоминирование (совместное доминирование). Это такой тип наследования, при котором у
потомков проявляются признаки обоих родителей в равной степени. Докажем на наследовании
групп крови у человека в системе АВО.
О
A
B
I
II
Антиген Н
Антиген А
III
Антиген В
I0I
IA
IB
AB IV
Антиген А + В
IAIB
Доказательство:
Р: IAIA IBIB
F: IAIB
Генетическим основанием Первого закона Менделя является то, что в скрещивание вступают
гомозиготы. Цитологической основой являются закономерности мейоза, а именно правило чистоты
гамет, по которому в каждую гамету закономерно попадает только одна гомологичная хромосома из
пары (только один аллель из пары. Это правило впервые было сформулировано в конце 19 века Бетсоном. Закон не соблюдается, если аллели локализованы в половых хромосомах, т.е. наследование
сцеплено с полом.
Закон расщепления (Второй закон Менделя)
. Определение: при скрещивании двух
моногетерозигот в потомстве наблюдается закономерное расщепление по генотипу: 25% гомозигот
одного вида, 50% гетерозигот, 25% гомозигот другого вида или расщепление в соотношении 1:2:1.
Расщепление по фенотипу может быть различным. В случае неполного доминирования и
кодоминирования оно совпадает с расщеплением по генотипу 1:2:1. в случае полного доминирования
расщепление 3:1.
доказательство при полном доминировании.
1.
Дано:
А желтые
аа – зеленые
Доказательство:
Р: АА +аа
F1: Аа ⊗
F2: АА:2Аа:аа
По генотипу 1:2:1
По фенотипу 3:1
Докажем с точки зрения теории вероятности закономерность расщепления по генотипу. Для
этого составим решетку Пеннета.
½ А
½ А ¼ АА
½ а
¼ Аа
½ а
¼ Аа
¼ аа
Вероятность события измеряется в долях единиц от 0 до 1 или в процентах от 0% до 100%. Если
вероятность равна 0, то такое событие не совершится никогда. Если вероятность события равна 1, то
оно совершается всегда. Вероятность события, состоящего из двух независимых событий, равна
произведению вероятности составляющих. Вероятность повторяющихся событий равна сумме
вероятности составляющих.
2.
доказательство при промежуточном наследовании.
Дано:
А красные
аа белые
Доказательство:
Р2: Аа +Аа
F2: АА: 2Аа:аа
По генотипу: 1:2:1
По фенотипу 1:2:1
3.
доказательство при кодоминировании.
Дано:
IA – II
IB – III
IAIB – IV
I0 I
Доказательство:
Р2: IAIB IAIB
F2:IAIA: 2IAIB:IBIB
По генотипу: 1:2:1
По фенотипу 1:2:1
Генетическими основами Второго закона Менделя является то, что в скрещивание вступают
моногетерозиготы. Цитологической основой является закономерное поведение гомологичных
хромосом в мейозе или правило чистоты гамет, по которому в каждую гамету попадает только одна
гомологичная хромосома из пары. Второй закон Менделя носит статистический характер, т.е. полученное расщепление является
количественной величиной. Для получения расщепления близкого к теоретически ожидаемому
необходимо проанализировать большое количество потомков в F2. Статистичность определяется
вероятностным характером двух процессов: вероятностный характер оплодотворения; вероятностный
характер оогенеза.
Второй закон Менделя соблюдается, если признаки не сцеплены с полом и существуют
следующие условия соблюдения: нормальный мейоз, т.е. равновероятное образование гамет разного
типа; равная выживаемость гамет; одинаковая оплодотворяющая способность гамет; равная
♂XY; F: : ХХ:♀ ♂XY. Но реально больше
вероятность выживаемости зигот. Нарушения: Р: ХХ +
рождается мальчиков, т.к. сперматозоиды с Y хромосомой обладают большей скоростью.
♀
Анализирующее скрещивание. Помимо классической схемы скрещивания Мендель проводил
бэккросс скрещивание или возвратное скрещивание. Одним из вариантов такого скрещивания
является анализирующее, при котором особь с неизвестным генотипом скрещивается с гомозиготой по
рецессиву. По результатам расщепления можно определить генотип особи, вступающей в
скрещивание.
Дано:
А комолый
Аа рогатый
♀
Доказательство:
♂
Р: АА + аа
FA: Аа
♂ ♀
Р: Аа аа
FA: Аа:аа
Закон независимого наследования и комбинирования признаков (Третий закон Менделя)
. Этот
закон был сформулирован при исследовании полигибридного скрещивания, т.е. скрещивания, в
которое вступают родители различные более чем по паре признаков. Определение: при скрещивании
родителей, различающихся более чем по паре признаков, каждая пара признаков подчиняется
расщеплению независимо от других пар, и признаки комбинируются. В результате в потомстве
появляются особи с новыми по отношению к родителям комбинациями признаков. Закон справедлив
только в том случае, если гены лежат в разных парах гомологичных хромосом, т.е. они не сцеплены.
Докажем при полном доминировании.
Дано:
А желтый
аа – зеленый
В – гладкий
Вв морщинистый
Доказательство:
Р: ААВВ+ аавв
FАаВв
♀
♂
АВ
Ав
аВ
АВ
ААВВ
Ав
ААВв
аВ
АаВВ
ав
АаВв
желтые гладкие
желтые гладкие
желтые гладкие
желтые гладкие
ААВв
Аавв
АаВв
Аавв
желтые гладкие
желтые морщ.
желтые гладкие
желтые морщ
АаВВ
АаВв
ааВВ
ааВв
желтые гладкие
желтые гладкие
зеленые гладкие
зеленые гладкие ав
АаВв
Аавв
ааВв
аавв
желтые гладкие
желтые морщ
зеленые гладкие
зеленые морщ
9:
3:
3:
1
желт глад
желт морщин
зеленые глад
зеленые морщ
генотипические радикалы:
АВ : Авв : ааВ : аавв
↑ ↑
Рекомбинантные классы
Докажем математически, что каждый признак независимо от другого подчиняется закону
расщепления и докажем появление рекомбиннантных классов.
По цвету: 12 : 4 = 3:1
желтые
зеленый
по форме:
12 : 4 = 3 : 1
гл. мор
(3+1)∙(3+1)=(3=1)2=32 + 2∙1∙3 +12=9+2∙3 +1
генетическим основанием Третьего закона Менделя является то, что в скрещивание вступают
полигетерозиготы. В данном случае дигетерозиготы. Цитологическим основанием является
закономерное поведение гомологичных хромосом в мейозе, т.е. правило частоты гамет. Кроме того,
цитологическими основаниями являются произвольная ориентация бивалентов относительно полюсов
клетки в метафазе 1 мейоза и в дальнейшем их произвольное расхождение в анафазе.
Третий закон Менделя носит статистический характер. Статистичность основана на
вероятностном характере оплодотворения и оогенеза. Статистичность позволяет вывести формулы
для подсчета гамет и подсчет генотипов, которые образуются при скрещивании одинаковых
гетерозигот. Гаметы можно подсчитать по формуле 2n, где 2 максимальное число гамет, которое дает
моногетерозигота, а n это степень гетерозиготности.
АаВв
22 = 4
АаВвССDdFf
24 = 16
Количество генотипов, которые образуются при скрещивании двух одинаковых гетерозигот
определяется по формуле 3n, где n это степень гетерозиготности, а 3 это максимальное количество
разных генотипов, которые образуются при скрещивании двух моногетерозигот.
Третий закон Менделя не соблюдается, если наследование сцеплено с полом и если гены сцеплены
между собой, кроме того закон не соблюдается во всех случаях описанных для второго закона. К
АаВв АаВв
32 = 9 полигибридному скрещиванию относится вариант тригибридного скрещивания, при котором
расщепление будет равно 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1
Анализирующее скрещивание
А желтый
аа – зеленый
В – гладкий
вв морщинистый
РА: АаВв+ аавв
FA: АаВв:ааВв:Аавв:аавв
Расщепление: 1:1:1:1
Хромосомная теория наследования
Была сформулирована на основе законов Менделя в основном благодаря группе ученых под
руководством Томаса Ханта Моргана. К концу 19 века стали известны закономерности митоза и
мейоза. Молодой американский ученый Сеттен предположил, что менделевские наследственные
факторы локализованы в хромосомах, однако первым это смог доказать Морган. В современном виде
можно выделить следующие положения хромосомной теории наследственности:
1. гены локализованы в хромосомах. Место положения гена называется локусом.
2. гены, локализованные в одной хромосоме, образуют группу сцепления и наследуются совместно
3. гены располагаются в хромосомах линейно. Расстояние между генами зависит от частоты
кроссинговера, который может протекать между ними. Частота кроссинговера выражается в
процентах.
Сцепленное с полом наследование
Первые доказательства хромосомной теории наследования было получено Морганом на
плодовой мушке дрозофиле. Этот объект является наиболее удобным в генетике, т.к. существует
много вариантов признаков и поэтому можно легко получить гомозиготные линии, которые будут
отличаться по ряду альтернативных признаков. Мушка образует большое количество потомков срок
между двумя генерациями не велик, всего 10 дней. Заслуга Моргана состоит в том, что он первым
ввел этот объект в генетику. Еще до Моргана было известно, что представители разных полов
отличаются друг от друга по набору половых хромосом и возможны два варианта наборов:
XX – гомогаметный пол
XY – гетерогаметный пол
У дрозофилы гомогаметный пол женский, гетерогаметный – мужской. Морган вывел
гомозиготные линии мух с красными глазами и с белыми глазами и проводил гибридизацию в том
порядке, в котором это делал Мендель. Он предположил, основываясь на работах Сеттена, что ген,
определяющий цвет глаз, локализован в Xхромосоме, а в Yхромосоме его нет. Благодаря своим
работам Морган смог это доказать.
Дано:
ХАкрасные
Хабелый
Доказательство:
Р: Х♀ АХА + Х♂ аY
F: ХАХа Х♂ АY
Подтверждает первый закон Менделя, т.е. закон единообразия первого поколения.
Р: Х♀ АХа + Х♂ АY
F: Х♀ АХА: ХАХа
Х♂ аY: ХАY
Подтверждается закон расщепления. Однако Морган обратил внимание на то, что белоглазыми в
данном скрещивании были только самцы, следовательно, Морган делает вывод, что ген,
определяющий цвет глаз, локализован в Ххромосоме, а в Yхромосоме его нет. Морган проводит
рецепрокное скрещивание.
Р: : Х♀ аХа+ Х♂ АY F: Х♀ АХа Х♂ аY
1:1
Нарушается закон единообразия первого поколения. Наблюдается крисскросс наследование,
т.е. дочери наследуют признак отцов, а сыновья – матерей. Явление крисскросс наследования это
нарушение первого законы Менделя, которое возможно благодаря тому, что признак локализован в Х
хромосоме.
Р: Х♀ АХа + Х♂ аY
F: Х♀ аХа: ХАХа
Х♂ аY:ХАY
1:1
Полученные результаты Морган объяснял тем, что ген локализован в Ххромосоме, а в Y
хромосоме его нет. Т.о. Морган подтверждает, что гены находятся в половых хромосомах.
Сцепленное с полом наследование – это наследование генов, расположенных в половых хромосомах.
Существует три варианта наследования сцепленного с полом:
Х + Y полностью сцепленное с полом наследование. Это значит, что ген локализован в Х
хромосоме. У человека приблизительно 200 генов локализовано в Ххромосоме, которых нет в Y
хромосоме. Некоторые из них имеют медицинское значение: ген, контролирующий свертываемость
крови. Ннорма, hгемофилия; ген, контролирующий цветовое зрение. Dнорма, dдальтоник. Для
млекопитающих характерен ряд особенностей при полностью сцепленном с полом наследовании. Одна
из Ххромосом у женского гомогаметного пола в клетке сильно спирализуется, превращается в тельце
Барра и инактивируется, т.е. в соматических клетках у самок млекопитающих функционирует только
одна Ххромосома. Процесс инактивации случаен, поэтому полностью сцепленное наследование у
млекопитающих имеет свои особенности. Классическим примером является наследование окраски
шерсти у кошек.
Дано:
ХАчерный
Харыжий
Доказательство:
Р: Х♀ АХА + Х♂ аY
F: Х♀ АХа Х♂ АY
F2: Х♀ АХа Х♂ АY: Х♂ аY: Х♀ АХА
Иногда встречаются черепаховые самцы, которые имеют генотип ХХY, они стерильны. У
человека также возможна инактивация одной из Ххромосом, что приводит к определенному
мозаицизму признаков. Например, существует болезнь, связанная с отсутствием потовых желез, у
мужчин потовые железы могут полностью отсутствовать, а у женщин могут располагаться мозаично на
теле. Мозаичность может проявлятся у женщин при дальтонизме. Женщина может быть дальтоником
на один глаз, либо дальтонизм проявляется на уровне сетчатки. Полностью сцепленное с полом
наследование у бабочек и птиц имеет свои особенности, т.к. у них гомогаметный пол – мужской, а
гетерогаметный – женский.
Дано:
ХВполосатые
Хвсплошные
Доказательство:
Р: Х♂ ВХВ:+ Х♀ вY
F: Х♂ ВХв: Х♀ ВY
F2: Х♂ ВХВ : Х♂ ВХв: Х♀ вY: Х♀ ВY
Р: Х♂ вХв + Х♀ ВY
F1: Х♂ ВХв: : Х♀ вY
У некоторых насекомых сцепленное с полом наследование имеет свои особенности, связанные с
тем, что представители разных полов различаются по количеству хромосом. Самки таких насекомых
имеют набор ХХ, а самцы ХО гаметы
ХХ♀
ХО♂
2n
2n1
⊗ ⊗ ⊗O
F: ХХ: ХО
♂
♀
1:1
X + Y + частично сцепленное с полом наследование, ген локализован и в Х и в
Y – хромосомах.
Частично сцепленное с полом наследование. В данном случае признаки наследуются от
гетерогаметного пола к гетерогаметному, а от гомогаметного к гомогаметному, т.е. крисскросс
наследования не наблюдается. Биологический смысл частично сцепленного с полом наследования
заключается в том, что это один из механизмов полового диморфизма. Ген, который контролирует
толщину эмали на зубах располагается и в Х и в Y хромосомах. изза того, что одна из Ххромосом у
гомогаметного пола спирализуется, эмаль на зубах толще у мужчин.
Дано:
ХАдомин
Харецессив
Доказательство:
Р: Х♀ АХА + Х♂ аYа
F: Х♀ АХа Х♂ АYа
F2: Х♀ АХа: Х♀ АХА: Х♂ АYа: Х♂ аYа
Рецепроктное
Р: Х♀ аХа Х♂ АYА
Х♀ АХа: Х♂ аYА
F:
F2: : Х♂ аYА ХАYА:
Х♀ АХа :ХаХа
X Y +
голандрическое наследование. Признак локализован только в
Y хромосоме.
Голандрическое наследование, при котором аллели локализованы в Yхромосоме. В Yхромосоме
находится не много генов, но как правило они определяют развитие первичных и вторичных мужских
половых признаков. Голандрическое наследование лежит в основе полового деморфизма. В Y
хромосоме локализованы следующее гены: SRY – этот ген отвечает за синтез белкового фактора,
который стимулирует развитие семенников; SPY – ген, который отвечает за сперматогенез; HYA ген,
который отвечает за синтез белка, входящего в состав плазмы крови. Наследование идет по
следующему принципу: от гетерогаметного пола к гетерогаметному полу.
Р: ХХ +ХY+
F: ХХ:ХY+ ⊗
F2: ХХ:ХY+
Существуют признаки, которые могут быть локализованы в аутосомах, но которые в норме
проявляются только у одного пола. Это признаки ограниченные полом, например, оволосенение,
яйценоскость у кур, этот ген лаколизован в аутосомах. Существуют признаки, зависящие от пола, эти
признаки поразному проявляются у представителей разных полов, например, у человека ранняя
алопеция.
♂ АА
лысый
♀ АА
р. в.
Аа
плеш
Аа
норм
аа
норма
Аа
норм
Генетика пола Пол – это набор определенных признаков, которые обеспечивают размножение. Определение
пола у разных животных осуществляется на разных этапах онтогенеза:
Пол определяется до момента оплодотворения (прогамное определение пола). Характерно для тех
видов, у которых гетерогаметным полом является женский.
Пол определяется в момент оплодотворения. Это так называемое сингамное определение пола.
Наблюдается в том случае, когда гетерогаметный пол мужской
Пол определяется внешними условиями (эпигамное определение пола). На ранних этапах развития
зародыш бисексуален. Есть гены для развития как женских, так и мужских половых признаков.
Решающее значение в этом случае имеет баланс или соотношение этих генов. У женского пола в Х
хромосоме есть гены, определяющие развитие женских половых признаков. В Yхромосоме есть гены
SRY и SPY. Существуют женщины генотипом ХY, у которых данные гены не работают.
До восьми недель беременности у эмбрионов формируется совершенно одинаковая система
первично половых признаков.
После восьми недель беременности начинаются различия, если плод генотипом XX, то под
действием гормонов матери первичные гонады преобразуются в яичники, которые как железа
внутренней секреции начинают работать только после периода полового созревания, следовательно,
развитие женского эмбриона определяется только гормонами матери. Под их влиянием дегенерируют
вольфовы протоки, а мюллеровы превращаются в яйцеводы или в фаллопиевы трубы. Из
мочеполового синуса развивается шейка матки, матка и верхняя часть влагалища. Из мочеполового
бугорка развивается нижняя часть влагалища и наружные гениталии. Гормоны матери действуют на
гипоталамус зародыша и детерминируют поведение по женскому половому типу после полового
созревания. У зародышей с генотипом XY из первичных гонад развиваются семенники, которые сразу
же начинают функционировать как эндокринные железы, т.е. у эмбриона начинает синтезироваться
тестостерон. В результате мюллеровы протоки дегенерируют, из вольфовых развиваются
семяпроводы, из мочеполового синуса и мочеполового бугорка семяизвергательный канал и канал
уретры, из мочеполового бугорка – мошонка и половой член. Тестостерон действует на гипоталамус
зародыша и после периода полового созревания определяется поведение по мужскому половому
признаку. С медицинской точки зрения существуют следующее уровни определения пола:
генетический и хромосомальный; гонадный, на уровне развития половых желез; гормональный;
генетальносоматический; психосоциальный или поведенческий.
СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ
Сцепленным наследованием называется наследование, при котором признаки локализованы в
одной хромосоме и наследуются совместно. Сцепленным наследованием занимался Морган на
дрозофиле, для которой было известно, что гаплоидное число хромосом равно 4, а альтернативных
пар признаков намного больше, поэтому Морган предположил, что гены, контролирующие некоторые признаки, локализованы в одной хромосоме. Морган исследовал два признака: окраску тела и длину
крыльев, и провел два рецепрокных скрещивания.
Доказательство:
Дано:
Асерый
аачерный
Вдлинные
ввзачаточные
Морган делает вывод: раз наследуются только родительские комбинации, то данные признаки
сцеплены, они локализованы в одной хромосоме и образуют группу сцепления. Это является
доказательством как 1ого, так и 2ого положений хромосомной теории. В данном случае наблюдается
так называемое абсолютное сцепление (крайне редкий вариант). Морган проводит рецепрокное
анализирующее скрещивание.
Так как в результате рецепроконого скрещивания получились рекомбинантные классы, то можно
сделать вывод, что сцепление между генами неполное или частичное. В этом случае появление
рекомбинантных классов можно объяснить с точки зрения того, что между гомологичными
хромосомами в процессе мейоза происходит кроссинговер. К моменту данных опытов мейоз был известен и было известно, что в профазе мейоза
гомологичные хромосомы коньюгируют с образованием бивалентов. Однако кроссинговер был
доказан Морганом теоретически и только через 20 лет был обоснован цитологически. Родительских
гамет всегда больше чем кроссоверных, причиной этого является то, что кроссинговер при мейозе
идет не в 100% случаев, можно учитывать только вариант нечетного кроссинговера, так как
результаты четного кроссинговера в потомстве будут незаметны.
Поэтому чистота кроссинговера теоретически не может превышать 50%. Различие двух
реципрокных скрещиваний объясняется тем, что у самок дрозофилы кроссинговер проходит, а у
самцов он не идет никогда, поэтому в первом варианте наблюдается редкий вариант абсолютного
сцепления. В целом частота кроссинговера у гомогаметного пола выше, чем у гетерогаметного пола.
Таким образом, этими опытами Морган доказал, что гены лежат в хромосомах и гены, локализованные
в одной хромосоме, образуют группу сцепления и могут наследоваться совместно. Частота
кроссинговера и, следовательно, частота появления кроссоверных гамет будет зависеть от расстояния
между генами
А – В – х%
В – С – y %
А – С – z %
Если анализировать попарное сцепление генов, то можно определить, что гены лежат в
хромосоме линейно.
Х%+ Y %= Z %
На основании линейного положения хромосом можно строить генетические карты хромосом. На
них хромосомы изображены в виде линий, на которых указываются гены и расстояния между генами.
Один процент кроссинговера является генетической единицей расстояния на такой карте. Было
принято считать, что 100% кроссинговера равняется 1 Моргану.
100% 1 Морган
1% 1 Морганида Помимо генетических карт хромосом существуют цитологические карты хромосом, причем
карты коллинеарны, то есть для них характерен одинаковый порядок генов. Цитологические карты
получают при дифференцировочной окраске хромосом.
Существуют варианты сцепления трех генов, причем ситуация, когда между всеми генами
происходит кроссинговер. Существует общий алгоритм решения такой задачи.
2 n =8
yxy=y(1x)
xxy=x(1y)
ГЕНОТИП И ФЕНОТИП
род. 1(х(1у)+(у(1х)=1х(1у)у(1х)=1х+хуу+ху=1ху+2ху
которое определяется последовательностью нуклеотидов.
Фенотип – это совокупность всех или анализируемых признаков, характерных для данного
организма. Фенотип развивается на основе внутренних факторов, материнских факторов и за счет
влияния факторов внешней среды. К внутренним факторам относится наследственная информация,
которую на уровне вида можно определить как геном, а на уровне организма понятием генотип
(совокупность аллелей). Гены в организме представлены в определенном физикохимическом
состоянии,
Конкретной
последовательностью нуклеотидов является аллель. Как правило, для данного вида организмов
характерны различные варианты аллелей, то есть фенотип определяет явление множественного
аллелизма. Множественный аллелизм заключается в том, что различные аллели контролируют
развитие более чем двух вариантов конкретного признака (конкретный вариант признака называется
феном). Причиной множественного аллелизма является внутригенная мутация, внутригенный
кроссинговер. Классическим примером множественного аллелизма является наследование групп
крови у человека. Гены в системе АВО отвечают за синтез различных галактозилтрансфераз.
Множественный аллелизм характерен для окраски глаз у дрозофилы.
С abr – абрикосовый
C cr коралловый
C + красный
C ch вишневый
C eor эозинофиловый C – белые
Множественный аллелизм характерен для окраски шерсти у многих млекопитающих.
Существует ген С, которые контролирует структуру фермента тирозолазы, субстратом для этого
фермента является тирозин, который под действием фермента превращается в пигмент меланин.
Существует два варианта меланина:
Существуют различные аллели данного гена:
Эумеланин
Феомеланин
С черный
C ch шиншилловая
C h – гималайская
С – белый
Множественный аллелизм лежит в основе генетического полиморфизма популяции.
Множественный аллелизм существует у растений в основном у перекрестно опыляемых, причем
существуют механизмы, которые поддерживают этот генетический полиморфизм. К таким
механихмам относится явление самостерильности у покрытосемянных растений, у них существует ген
S , который определяет проростание пыльцевой трубки.
Существует несколько аллелей генов S ( S 1, S 2, S 3…).
Влияние генотипа на фенотип как правило очень сложное и один ген может влиять не на один, а
на целую группу признаков. Такое явление получило название плейотропия или множественное
действие генов. Различают два варианта плейотропии: первичная или структурная; вторичная или
функциональная. При первичной плейотропии патологичен продукт гена, но это влияет на множество
признаков в организме. Синдром Марфана, болезнь, связанная с патологией в синтезе каллогена. В
результате патологично большинство каллогеновых структур, в организме наблюдается деформация
скелета, рост, как правило, выше среднего, непропорционально удлинены конечности,
арахнодактелия, патологии суставных сумок, различные пороки сердца, патологии органов зрения.
Вторичная плейотропия. Классическим примеро является серповидноклеточная анемия. Эта аномалия
вызвана одной аминокислотной заменой в Вцепи гемоглобина, в результате гемоглобин плохо
связывается с кислородом и ткани накодятся в постоянном дефиците кислорода. Это доминантное
заболевание, гомозиготы не выживают, у гетерозигогт наблюдается увеличение селезенки,
гипертрофия сердца, гипертрофия красного костного мозга. Гетерозиготы оказываются в некоторых
условиях более жизнеспособны, чем гомозиготы по рецессиву (устойчивость к малярии).
Патологичные гены не всегда проявляются в фенотипе, причем они могут проявляться с разной
степенью выраженности. Вероятность проявления данного аллеля в фенотипе называется
пенетрантностью. Если аллель всегда проявляется в фенотипе, то говорят о полной пенетрантности,
если аллель не проявляется в фенотипе в различных случаях, то говорят о неполной пенетрантности.
Например, полидактилия – это болезнь с неполной пенетрантностью. Из 100 носителей аллелей она
проявляется в фенотипе с разной степенью выраженности. Например, полидактилия – это появление пятого пальца на всех конечностях, либо только на одной, в виде нормально развитого пальца, либо
только в виде зачатка фаланги, в этом случае говорят о вариабельной экспрессивности развития
признаков. Экспрессивность развития признаков бывает константной, если данный аллель всегда
проявляется в фенотипе. Влияние генотипа на фенотип проявляется в явлении взаимодействия генов.
ВЗАИМОДЕЙСТВИЕ ГЕНОВ
Выделяют две крупные ситуации:
Взаимодействие аллельных генов. Аллельные гены физически могут взаимодействовать между
собой во варианте внутреннего кроссинговера. Под взаимодействием аллельных генов понимают
взаимодействие белковых продуктов генов, это взаимодействие возможно в следующих ситуациях:
полное доминирование; промежуточное наследование; кодоминирование. Причем генотип однозначно
определяет фенотип не во всех ситуациях, а только в ситуации промежуточного наследования и
кодоминирования. Полное доминирование характерно для генов, которые контролируют структуру
ферментов, например, ген IА полностью доменирует над геном I0. промежуточное наследование
характерно для структурных белков, белковрецепторов, белковпереносчиков, для ферментов со
средней активностью, классическим примером является наследование структуры волоса.
Кодоминирование является наиболее встечающимся вариантом наследования на биохимическом
уровне. Классическим примером кодоминирования является проявление 4 группы крови в системе
АВО.
Взаимодействие неаллельных генов. При формирование фенотипа на развитие одного признака,
как правило, влияет несколько генов, это так называемый контроль развития признака или
взаимодействие неаллельных генов. Физически взаимодействие неаллельных генов возможно только в
группе сцепления, поэтому рассматривается взаимодействие на уровне белковых продуктов генов.
Возможно два варианта этого взаимодействия:
последовательные реакции
группа реакций. Например, это синтез глазных пигментов у мухи дрозофилы.
Выделяют три варианта взаимодействия неаллельных генов:
комплиментарность – это такой тип взаимодействия неаллельных генов, при котором наличие в
генотипе доминантных аллелей всех взаимодействующих генов определяет развитие самостоятельного
признака. Классическим примером является синтез глазных пигментов у дрозофилы.
Р: ВВСС+ААdd aaDD
F: ВВСС+АаDd F2: BBCC+
AD
9:
Add
3:
красные
коричнив.
aaD
3:
белые
aadd
1
Белые
9:3:4
FA:АаDd aadd
Расщепление 1:2:1
Р: ААВВ+ССdd ccDD
F: ААВВ+СсDd
F2: ААВВ+
СD
9:
Cdd
3:
красные
коричн
ccD
3:
ярк. красн
9:3:3:1
Ccdd
1
белые
в анализирующем скрещивании расщепление 1:1:1:1
При наследовании окраски глаз у дрозофилы возможны варианты расщепления 9:7, в
анализирующем скрещивании 3:1. существует еще один вариант расщепления при комплиментарности
9:6:1, доказывается при изучении наследования формы плода у тыквы.
Дано:
АВдисковидные
Аввсферическии
АаВсферическии
ааввбутылковидные
Доказательство:
Р:Аавв ааВВ
F:АаВв
F2:
АВ
АаВ
Аавв
дисков
9:
3:
сф
Расщепление 9:6:1
Авв
3:
сф
1
бутыл
В анализирующем скрещивании 1:2:1
По типу комплиментарности наследуется окраска волнистых попугайчиков. У них
синтезируется желтый и голубой пигменты, при взаимодействии этих пигментов формируется зеленая
окраска.
Эпистаз – это такой тип взаимодействия неаллельных генов, при котором действие аллелей
одного гена подавляет действие аллелей другого гена, ген подавитель называется эпистатическим,
геном ингибитором, геном супрессором. Ген, действие которого подавляется, называется
гипостатическим. С точки зрения характера аллелей ингибиторов в эпистазе выделяют 2 ситуации:
доминантный эпистаз; рецессивный эпистаз. Рассмотрим на примере окраски мышей.
Дано:
Доказательство: В анализирующем 1:2:1
Дано:
А черный
аакоричн
Весть рапред
пигм
ввнет распред
Доказательство:
Р: Аавв ааВВ
F:АаВв
F2:
АВ
9:
черн
АаВ
3:
корич
Авв
3:
белые
Аавв
1
белый 9:3:4
Рецессивный эпистаз может трактоваться и как эпистаз и как комплиментарность.
Полимерия – это такой тип взаимодействия неаллельных генов, при котором действие
доминантного аллеля одного гена полностью или частично дублирует действие доминантного аллеля
другого гена), есть на проявление признака необходимы доминантные аллели одного из
взаимодействующих генов). Существует два варианта полимерии: кумулятивная полимерия касается в
основном количества признаков; некумулятивная полимерия касается в основном качества признаков.
Классическим примером некумулятивной полимерии является наследование махноногости у лошадей
(кур).
Дано:
Амахноногии
аанорма
Вмахноногии
ввнорма
Доказательство:
Р:Аавв ааВВ
F:АаВв
F2:
АВ
9:
махноног
9:3:3:1
15:1
Авв
3:
мах
Аавв
1
норма
АаВ
3:
мах
Однако реально в данном случае расщепление 9:6:1.
Некумулятивную полимерию можно трактовать как комплиментарность. По данному типу
взаимодействия неаллельных генов наследуется форма плодов пастушьей сумки. В анализирующем
скрещивании расщепление 1:2:1. Кумулятивная полимерия связана с неполным дублировании генов с
неполным доминировании по каждому гену. Классическим примером является степень пигментации
кожи. Реально степень пигментации зависит от 5 генов, однако для простоты будем считать, что она
зависит только от 2 генов.
Дано:
ААВВчерн
ааввбелые
Доказательство:
Р:ААВВ аавв
F:АаВв мулаты
АВ
Ав
аВ
АВ
ААВВ
черн
ААВв
Тем. мулат
Ав
ААВв
Тем. мул
Аавв
Мулат
аВ
АаВВ
Тем. мул
АаВв
Мулат
Ав
АаВв
Мулат
Аавв
Св. мулат
АаВВ
Тем. мулат
АаВв
Мулат
АаВВ
АаВв ав
АаВв
Аавв
ааВв
Аавв
1:
черные
4:
светлые
мул
6:
мулат
4:
светлые
мул
1
белые
В анализирующем скрещивании расщепление 1:2:1
Ген и его свойства
Ген функционально един. Функциональное единство проявялется в процессе транскрибции,
причем ген ограничен положением инициатора и терминатора. Ген структурно делим. Структурная
делимость проявляется в процессе внутреннего кроссинговера. Ген относительно устойчив. Эта
устойчивость проявляется в процессе передачи аллелей от одной клетки к другой без изменений,
однако эта устойчивость относительна т.к. гены могут изменяться в процессе мутации либо
внутреннего кроссинговера. Относительность этой устойчивости демонстрирует явление
множественного аллелизма. Стабильность гена – это значит, что каждый аллель характеризуется
определенной стабильностью своего проявления в фенотипе. Гены обладают плейотропным
эффектом. Роль гена в биосинтезе белка заключается в том, что в процессе транскрибции
считываются РНК, необходимые для образования всех компонентов белоксинтезирующего аппарата. В
процессе транскрибции помимо различных функциональных белков считываются регуляторные белки,
которые способны активировать или ингибировать действие структурных генов, т.к. гены эукориот
имеют мозаичное строение, а матричные РНК интронэкзонную структуру, то синтез различных
белков и их активность регулируется за счет альтернативного сплайсинга.
Генотип, как целостная система
Геном включает в себя: совокупность ядерных генов или нуклиом, а также совокупность генов
митохондрий и пластид. Совокупность митохондриальных генов, получило название хондриом.
Совокупность генов пластид называется пластидом. Гены, локализованные в митохондриях и
пластидах, называется плазмогенами, и существуют определенные закономерности их наследования,
которые получили название цитоплазматическое наследование. Расхождение митохондрий и пластид в
мейозе случайно, поэтому наследование плазмогенов не подчиняется законам Менделя и Моргана.
Определенные закономерности наследования плазмогенов проявляется на фоне анизогамии. Во
варианте анизогамии половые клетки отличаются друг от друга по количеству митохондрий и
пластид, а следовательно, плазмогенов в сперматозоидах этих органов практически нет, кроме того
при попадании в яйцеклетку митохондрий и пластид часто разрушаются, поэтому для будущего
зародыша имеет значение только плазмогены матери. Такой вариант цитоплазматического
наследования получил название матрокления. Классическим примером является наследование
пестролистности у растений (например, герань).
♂
пестролист зеленые растения
♀
↓
зеленый
♀
пестролист
♂
зеленые листья
↓
появление во втором случае разных листьев у герани объясняется с точки зрения явления
митотической сегрегации.
зеленые, пестрые, белые Для человека также характерно явление матроклении. В митохондриях человека содержится 36
генов, 22 для тРНК, 2 гена для собственных рРНК, а все остальное для белков цепи переносчиков,
следовательно, при потологии этих генов могут наблюдаться дифекты окислительного
фосфолилирования, энергообмен сдвигается к гликолизу и развивается лактоацидоз. Наблюдается
митохондриальные болезни различной степени тяжести.
Различная тяжесть заболевания является причиной митатической сигригации. Существуют
определенные варианты псевдоматроклинии, например, у отрядя легочных брюхоногих моллюсков
бывает левозакрученная и правозакрученная раковины. Оказалось, что закрученность раковины
является результатом спирального дробления зиготы на ранних этапах развития зародыша. В это
время гены зиготы не работают и правозакрученность раковины определяется только наличием белкка
в цитоплазме зиготы, если данного белка нет, то раковина закручена влево. Наличие белка
определяется доминантным аутосомным геном. Фенотип зависит от генотипа и это отражает явление
изменчивости.
Изменчивость.
Это свойство живых систем и их компонентов существовать в различных вариантах. Выделяют
следующии варианты изменчивости: генотипическая изменчивость. В этом случае изменение генотипа
приводит к имзменению фенотипических признаков; эпигеномная изменчивость. Изменеие фенотипа
проходит без имзменения генотипа и основано на изменении регуляции действия гена. Эпигиномная
изменчивость является механизмом деференцировки клеток у многоклеточных; модификационная
изменчивость. Проходит без изменения генотипа под действием факторов внешней среды.
ГЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ
Определение смотри выше. Делится на два крупных варианта:
комбинативная изменчивость – это изменчивость в основе которой лежат кроссинговер,
произвольная ориентация бивалентов в метофазе 1, вероятностный характер оплодотворения.
определение кроссинговера, история открытия (анализирующие скрещивание на дрозофилах) рисунок
метофазы 1, объяснить вероятностный характер оплодотворения.
мутационная изменчивость. Мутации – это качественные изменения генетического материала.
Термин мутации был впервые введен в 19021903 тремя учеными Гуго де Фрец, Корренс и Черман.
Существует много классификаций мутаций: по месту их прохождения. В этом случае мутации делят на соматические мутации, проявляются у
родителей ввиде новообразований, могут передаваться по наследству только в случае вегетативного
размножения. Для человека характерно вегетативное размножение только как патология, это
появление однояйцевых близнецов. Генеративные мутации проходят в половых клетках и всегда
передаются по наследству.
мутации можно классифицировать по их влиянию на жизнеспособность; витальные мутации не
оказывают влияние на жизнеспособность; супервитальные они способствуют повышению
жизнеспособности, примером является гетерозоготы по серповидноклеточной анемии; полулетальные
мутации снижение жизнеспособности, например альбинизм, синдром Дауна; летальные мутации
несовместимы с жизнью синдром Патау, синдром Эдвардца.
мутации можно классифицировать по масштабу генетических структур, которые затрагиваются
входе мутации: генные мутации – изменение структуры гена; хромосомные мутации изменение
структуры хромосом; геномная мутация это изменение количества информосом в нуклеоме.
Генные мутации
Изменение структуры гена – это изменение числа нуклеотидов, либо изменение
последовательности нуклеотидов. Такие мутации могут возникать спонтанно или могут быть вызваны
ошибками "трех R ": репликация, рекомбинация (кроссинговер, произвольная ориентация бивалентов
в метофазе 1), репарация. Наименьшие изменения структуры гена касаются изменения только в одной
паре нуклеотидов, так называемые точковые мутации. При мутациях пуриновые нуклеотиды могут
менятся на пуриновые, а перемидиновые на перемединовые, такие мутации называются транзиции.
Входе мутации меняются пуриновые на перемединовые и наоборот такие муьтации называются
трансверзии. Возможно три варианта точковых мутаций: замена, выпадение, вставка.
Последствия замены: не происходит изменения в порлипептидной цепи (вырожденность
генетического кода). Такие мутации не ведут к изменению жизнеспособности и называются сейнсис
мутации. Может измениться первичная структура белка на одну аминокислоту (спецефичность
генетического кода). Последствия такой замены различны, т.е. это либо миссенс мутации, когда белок
функционирует плохо. Первичная структура белка может стать короче изза появления нонсенс
кодона, нонсенс мутации, прекращается синтез белка. Белок может удлиниться, если замена
произойдет в нонсенс кодане. Белок может стать короче изза изменения кадона инициации.
Последствия вставки или выпадения более серьезны: Меняется структура полипептида с места
выпадения или вставки (неперекрываемость и непрерывность генетического кода) изза сдвина рамки
считывания. С большой вероятностью появляется нонсенс кодон и синтез белка прекращается.
Генные мутации являются одной из причин возникновения новых аллелей и поэтому являются
причиной множественного аллелизма. С этой точки зрения они бывают доминантными или
рецессивными в ряде случаев генные мутации ведут к фенотипическим изменениям и к появлению
различных наследственных заболеваний (полидактилия, дальтонизм, серповидно клеточная анемия,
различные формы сахарного диабета, витамин Д независимый рахит).
Спонтанные генные мутации являются ошибками "трех R ". Существуют индуцируемые мутации,
которые вызваны действиями физикохимичеких агентов на ДНК. Частота спонтанных мутаций 106,
индуцируемых 103 (тяжелые металлы, ультрафиолет, подавляющие большинство лекарственных
препаратов). Если такие мутации произошли в генеративных клетках, то они передаются потомству,
если в соматических , то возможно формирование раковой опухоли. Поэтому большинство мутагенов
является канцерогенов. В клетках существует механизмы исправления генетических повреждений и
восстановления нормальной структуры ДНК. Такие механизмы получили название репарации.
Репарации Существуют несколько вариантов репарации: темновая репарация; пострепликативная
репарация.
Темновая репарация происходит в темноте и связана с вырезанием патологичного участка ДНК
β
ДНКполимеразой по правилу
ферментом экзонуклеазой, затем брешь достраивается
комплиментарности,
это
ферментативный процесс, который идет с затратой АТФ. У человека существуют патологии при
которых данный вид репарации не работает, в результате возникает заболевание пигментная
ксеродерма (вид рака кожи).
окончательное сшивание цепи ДНК осуществляется лигазой,
Пострепликативная проходит в том случае, если не срабатывают механизмы темновой
репорации, проходит после репликации, когда синтезируется патологичная ДНК.
Если не включается механизм репарации, то патологичные ДНК расщепляются. Механизмы
пострепликативной репарации основаны на внутригенном кроссинговере.
В первой молекуле по нашему рисунку брешь достраивается ДНКполимеразой и включается
лигаза. Вторая молекула подвергается темновой репорации. Если патологична пострепликативная
репарация, то возникает пигментная ксеродерма.
Хромосомные мутации
β
Это изменения структуры хромосом. Выделяют два варианта:
внутрихромосомные мутации (перестройки, обберации) касаются структуры одной хромосомы, либо
пары гомологичных хромосом. К внутрихромосомным мутациям относятся:
Делеция или нехватка заключается в выпадении какоголибо участка хромосом.
Дуплекация – это удвоение какоголибо участка ДНК, возможны триплекации, тетропликации и
т.д. Инверсия – это поворот участка хромосом на 1800.
Транспозиция – это перемещение участка хромосом.
Образование кольцевых хромосом – в этом случае слипаются концы хромосом, образуя кольцо.
При репликации возможно разрушение кольцевых хромосом.
Наиболее опасны последствия делеции. Причем важно какие гены выпали, домашнего хозяйства
или гены роскоши. Последствия при делеции как правило несовместимы с жизнью. У гетерозигот по
делеции наблюдаются тяжелые последствия связанные с нарушением баланса генов. У дрозофилы
существует делеция "ночь", которая заключается в возникновении вырезки на крыле. У человека
известна дилеция по пятой паре хромосом, которое происходит в коротком плече хромосомы.
отставание в физическом и умственном развитие погибают в первые годы жизни. характерно
изменение гортани, при котором новорожденный издает специфический крик, синдром кошачьего
крика.
Последствия дупликации обычно более мягкие, чем последствия дилеции. Однако нарушается
баланс генов, что отражается на фенотипе. У дрозофилы известна дуплекация Bar , которая приводит
к уменьшению фасеток в глазах. у гетерозигот число фасеток в глазах небольшое, у гомозигот число
фасеток резко уменьшается, возникает полосковидный глаз.
Механизмы делеции и дупликации. До недавнего времени считалось, что данные мутации
возникают изза разрыва ДНК, а затем изза ее дальнейшего сшивания. В настоящее время считают, что
это возможно, но только под действием очень сильных мутагенов. Основной причиной спонтанного
возникновения дилеции и дупликации является кроссинговер. В ДНК очень много гомологичных
последовательностей, которые могут конъюгировать между собой. Изза неравноценного
кроссинговера в гомологичных хромосомах возникает делеция или дуплекация.
Последствия инверсии наблюдаются как в гомозиготах, так и в гетерозиготах, однако инверсия
приводит только к изменению порядка в расположении генов, поэтому они могут вообще не оказывать
влияние на фенотип. У гомозигот по инверсии последствия наблюдаются редко, так как между
гомологичными хромосомами может проходить нормальный кроссинговер. У гетерозигот по инверсии
наблюдается высокий процент стерильности изза образования несбалансированных гамет и высокий
процент возникновения патологий у потомства. Это происходит изза того, что гомологичные
хромосомы не могут нормально конъюгировать и при конъюгации образуется так называемая
инверсионная петля.
Инверсии являются эволюционным механизмом, который создает изоляцию между многими
близкими видами, так как для близких видов часто характерны одинаковые инверсии.
Механизм образования инверсии. Гомологичные участки ДНК в пределах одной молекул могут
конъюгировать между собой. При транспозиции все гены остаются на месте, но часть из них меняют свое положение.
Последствий может не наблюдаться однако, механизм образования транспозиций связан с
образованием кольцевых хромосом при разрыве такой кольцевой хромосомы происходит потеря генов
и, следовательно, последствия будут аналогичны дилеции.
Образование кольцевых хромосом. Механизм смотри выше. При репликации кольцевых
хромосом они характеризуются очень высоким уровнем нестабильности. При репликации кольцевых
хромосом часто наблюдается перекручивание ДНК, когда кольцо попадает в кольцо.
Тогда при расхождении кольцевых ДНК будет наблюдаться разрыв молекул и утраты генов.
Последствия будут сходны с последствиями дилеции.
межхромосомные мутации затрагивают структуру двух и более негомологичных хромосом. Этот вид
мутации получил название транслокации. При транслокации происходит обмен участками ДНК входе
кроссинговера между негомологичными хромосомами. Это возможно благодаря тому, что в ДНК
негомологичных хромосом существуют короткие гомологичные последовательности.
Существуют как гомо, так и гетерозиготы по транслокации. У гомозигот как правило
последствий не наблюдается, т.к. все гены остаются на месте. У гетергзигот по транслокации, как
правило, наблюдается высокий процент стерильности в потомстве и аномалии. Это связано с тем, что
не гомологичные хромосомы в мейозе выстраиваются в форме креста. Геномные мутации
Это мутации связанные с изменением количества отдельных хромосом, либо с изменением целых
хромосомальных наборов. Делятся на два варианта:
эуплоидия. В этом случае число хромосом изменяется на число равное гоплоидному набору
хромосом. Эуплоидия может быть вызвана двумя различными механизмами и сэтой точки зрения
делятся на: автоплоидию – изменение хромосомального набора, за счет хромосом одного и того же
вида; аллоплоидия – изменение хромосомального набора за счет хромосом разного вида. С этой точки
зрения аллоплоидия это всегда результат межвидовой гибритизации
аннеуплоидия – это изменение числа отдельных хромосом.
2n1
2n+1
2n1+1
2 n 2
Моносомик по 5 паре
Трисомик по 5 паре
Моносомик по 3 паре
Трисомик по 5 паре
Нулесомик по…
Автоплоидия
Может привести не расхождение сестринских хроматид во втором делении мейоза, при этом
может быть патологично веретено деления, либо кинетохорный комплекс. Существуют определенные
химические вещества, которые могут блокировать сборку веретена деления или работу кинетохорного
комплекса, например, алкоголь, либо растительный алкалоид колхицин. Однако, этот механизм может
привести только к появлению четных полиплоидов. Для нечетных полиплоидов нужна внутривидовая
гибридизация. Гоплоидность практически несовместима с жизнью у многоклеточных организмов.
Проявляются все рецессивные патологичные аллели. Гоплоиды практически не имеют применений,
однако, у растений получают гоплоиды, проращивая пыльцевое зерно, и их используют для получения
чистых линий. Полиплоидия также оказывает влияние на жизнеспособность. для животных
полиплоидия практически нехарактерна, исключения некоторые птицы и караси, у которых возможно
существование триплоидов. Такие виды могут размножаться путем партеногенеза, то есть яйцеклетка
невступает в мейоз и без оплодотворения приступает к дроблению. Полиплоидия у растений явление более обычное. Полиплоиды у растений существуют до 5 n и 6 n . Нечетные полиплоиды стерильны и
не образуют семян. Такая ситуация возможна изза невозможности нормального протекания мейоза, то
есть гаметы у нечетных полиплоидов несбалансированны. В результате образуются бессемянные
сорта, например, виноград без косточек, бессемянные сорта огурцов, бананов, арбузов, которые
размножаются вегетативно. При потери такой культуры такие растения можно получить за счет
внутривидовой гибридизации. Схему смотри выше. Полиплоидия и у животных, и у растений ведет к
появлению гетерозиса, когда полученные гибриды крупнее и более мощные, чем нормальные
диплоидные организмы.
Аллоплоидия.
Предпосылкой аллоплоидии является межвидовая гибридизация. У растений гибридизация
может быть межродовой. Существует общее правило, чем родственные виды, тем успешнее проходит
гибридизация. Для животных известны следующие гибриды: лошадь+осел=мул, ослица+конь=лошак,
лев+тигр=лигр, белуга+стерлиц=бестер. У растений изучением аллоплоидов занимался Корпеченко.
Он скрещивал растения в пределах крестоцветных и получил гибрид, который назвал рафанобрассика.
Для аллоплоидов животных и растений характерно явление гетерозиса и общим правилом является их
стерильность. Корпеченко получил нестерильный гибрид рафанобрассики. Хромосомы редьки и
капусты частично гомологичны.
В промышленности используют аллоплоиды растений таких, как третикале гибрид ржы и
пшеницы, фертильный.
Аннеуплоидия.
♂
Это изменение числа отдельных хромосом как в сторону увеличения так и в сторону
уменьшения. Причиной аннеуплоидии является потеря хромосом, либо их не расхождение в мейозе.
По последствиям аннеуплоидия аналогична дилеции и дуплекации, но только более крупного
масштаба. Резко изменяется баланс генов, поэтому аннеуплоидия по крупным аутосомам практически
несовместима с жизнью. Возможно существование аннэуплоидов по половым хромосомам, такие
патологии получили название хромосомальный болезни.
47, XYY синдром "вайвай". Это мужчины в среднем выше ростом, чем норма фертильны.
Характеризуются нестандартным поведением в стрессовых ситуациях. Впервые "вайвай" был изучен в
тюрьмах.
45; ХО синдром ТернераШиншевского. Женщины, как правило, ниже среднего роста. Много
гибнет до рождения, до периода полового созревания развивается в целом нормально, после перестает
расти, гаметогенез нарушен, то есть, стерильны, интеллект ниже нормы, характеризуются женским
инфантилизмом.
♀ ♀
47; ХХХ синдром трисомии по Х. Женщины – рост выше среднего, фертильны, причем в
потомстве очень редко наблюдается трисомии по Х. Интеллект в среднем ниже нормы. Характерна
склонность к наркомании и алкоголизму. Повышенная склонность к дромамании.
47 ХХ Y синдром Клейнтфельтера. Мужчины рост в среднем выше среднего, стерильны. До
периода полового созревания в целом развивается нормально. После полового созревания становится
евнуховидного сложения, развиваются молочные железы, интеллект ниже среднего, легко управляемы.
♂
Аннеуплоиды у человека по аутосомам практически не совместимы с жизнью.
47, 13+1 синдром Патау
47, 18+1 синдром Эндвардца
Погибают либо на стадии внутриутробного развития, либо в первые дни жизни. Совместимы с жизнью
только аннеуплоиды по 22 и 21 хромосоме синдром Дауна.
47, 21+1
47, 22+1
очень много гибнет до рождения. Тяжелые формы умственного отсталости, доживают до половой
зрелости, фертильны, в потомстве 4050% с синдромом Дауна.
Эпигеномная изменчивость.
Этот вариант изменчивости заключается в закономерном и необратимом изменении фенотипа
клеток в ходе индивидуального развития без изменения генотипа. Эпигеномная мутация лежит в
основе диференцировки клеток в многоклеточном организме. Она основана на регуляции действия
генов и в первую очередь на регуляцию действия генов роскоши. При дроблении зиготы образуются
бластомеры, для которых характерен полный набор генов, содержится в зиготе. Однако в цитоплазме
бластомеров локализованы различные белковые материнские факторы, которые включают различные
гены роскоши. Чем дальше идет дробление, тем сильнее различие по активности генов роскоши.
Кроме того на определенной стадии развития зародыша бластомеры оказываются в условиях
различной позиционной информации. Таким образом, в основе эпигеномной изменчивости находятся:
собственные регуляторные белки; факторы, поступающие от внешней среды, либо со стороны других
местах; эпигиномная изменчивость основана на каскадной регуляции действия генов.
Модификационная изменчивость.
Изменение фенотипа под влиянием факторов внешней среды, которое не сопровождается
изменением генотипа. Одна и та же генетическая информация при разных внешних условиях может
быть поразному реализована. Изменение фенотипа зависит от того, насколько жестко развитие
данного признака контролируется генотипом. Однако для каждого признака существует некая
средняя величина и отклонением от нее, то есть вероятный характер и описывается кривой нормы.
Кривые нормы распределения носят статистический характер, то есть для анализа
вырожденности признака необходимо взять большую выборку особей. Тогда существуют
определенные пределы изменчивости, в которых данный фенотипический признак может измениться без изменения генотипа. Этот параметр получил название норма реакции, норма реакции жестко
контролируется генотипом и зависит от всего генотипического фона. Таким образом наследуется не
данный признак, а норма реакций. Впервые понятие нормы реакции ввел Иогансен, исследуя развитие
признаков на фасоли. Фасоль – это самоопыляемое растение, у которого легко можно получить
чистые линии. Иогансен брал чистые линии и высаживая их в различных условиях.
В конкретных условиях проявляется только часть нормы реакции. Классическим примером
этого явления является растение стрелолист. На одном растении возможно формирование различных
листьев в зависимости от условий внешних среды. Подводные листья растения нитевидные, листья,
находящиеся в воздухе, стреловидные. Иогансен доказал важное свойство модификации, а именно их
ненаследуемость. Иогансен брал семена с одного растения, но различные по весу и высаживал их в
абсолютно одинаковых условиях. =х3
х=х1=х2
Вторым важным свойством модификации является их обратимость (загар у
человека). Причины модификации: изменение регуляции действия гена; прямое действие
внешних факторов на развитие признаков. Регуляция действия генов подразумевает
включение определенных рецепторных систем в ответ на сигнал, поступающий из внешней
среды. При этом активируются гены роскоши. При прекращении поступлении сигнала,
данные гены ингибируются (почвенная амеба, активируются гены для денеина и тубулина).
Прямое влияние внешних факторов внешней среды. В основном это влияние оказывается
на степень пигментации и распределение пигмента. Например, окраска у примулы.
RR – красные
rr белые
R ` R ` окраска разная при различной температуре
Если температура окружающей среды ниже 250, то цветы красные, если температура,
при которой развиваются бутоны выше 250, то цветы белые.
Например, гемолайская окраска у кроликов
СС – черный
сс – белый
С h C h – гемолайский.
Оказалось, что аллель С h контролирует определенное распределение пигмента, при
котором темную окраску приобретают только терминале (уши, морда, хвостик, лапки), так
как на терминалах температура тела ниже.
Существуют варианты необратимых модификаций: длительные модификации. У
простейших классическим примером является устойчивость к повышенной температуре. У
инфузорий. Если клоны инфузорий выращивать при повышенной температуре, то у них
развивается устойчивость к ней. Если такой клон поместить в нормальные условия, то
устойчивость к повышенным температурам сохраняется еще несколько поколений. Это
явление получило название длительной модификации. В настоящее время считается, что
причиной являются шоковые белки, которые вырабатываются у инфузорий, а затем как
материнский фактор передаются следующему поколению; необратимые модификации
возникают при действии на зародыш различными терагенными факторами. Эти факторы
вызывают появление морфоз или уродств (морфозы не наследуются)
Генетика человека.
Все закономерности генетики, открытые на других объектах справедливы и для
человека. Человек – это биосоциальное существо, которое характеризуется
определенными особенностями: Биологические особенности: малоплоидность, малоплодовитость. Нормой для
человека является одноплодная беременность. Близнецы являются патологией.
Редуктивный период от 1314 лет до 5055 лет. За это время у женщины может
овулировать 300400 яйцеклеток, однако, изза продолжительного срока беременности на
свет может появиться не более 30 детей. Относительно длительный срок полового
созревания 1314 лет. Это делает невозможным исследования по классической
мендалевской схеме. Кариотип человека n =23. с трудом поддается анализу.
Социальные особенности: невозможность экспериментов; если есть уверенность , что
данный ребенок это ребенок данной матери, то установить отцовство затруднительно;
социальноэкономические причины приводят к тому, что биологическая плодовитость не
реализуется; невозможность выравнивать существования всех семей, что особенно важно
для человека с неполной пенетрантностью и вариабельной экспрессивностью,
следовательно, у человека невозможно использование классического гибридологических
методов и поэтому используют его эквивалент.
Генеалогический метод
Он основан на сборе информации о фенотипах родственников и на основе этой
информации строится генеалогическое древо. Оно отражает: степень родства; показывает
наличие анализируемого признака у данной группы родственников. Исследуемый признак
обозначают цветом. Существуют определенные правила составления родословного древа:
представители одного поколения изображаются на одной горизонтали; представители
старшего поколения изображаются выше младшего; поколения нумеруются римскими
цифрами; генеалогический метод позволяет определить характер наследования признаков;
доминантный или рецессивный признак; аутосомный или сцепленный с полом;
контролируется одним геном или несколькими; наследование признаков сцеплено или не. В
родословной обращается особое внимание на так называемые узловые моменты или
критические моменты. Для доминантных признаков узловым моментом является передача
в ряду поколений без перерывов. Для рецессивных признаков узловым моментом является
появление детей, несущих данный признак от родителей, которые данный признак не
имеют.
Особенно велика вероятность проявления рецессивных признаков в
близкородственных браках, так как у одной группы родственников высока степень
гетерозиготности по данному признаку. Для доминантного сцепленного с полом признака
узловым моментом является, когда у отца, имеющего данный признак, все дочери несут
данный признак (крисскросс наследование), если мать не имеет признака, то все мальчики
здоровы. Для рецессивного сцепленного с полом признака узловым является ситуация,
когда у родителей, несущих данный признак, в потомстве он проявляется только у
мальчиков. Для абсолютного большинства признаков у человека характерен полигенный
контроль, основанный на взаимодействии неаллельных генов.
Близнецовый метод
С помощью близнецового метода можно оценить относительный вклад генотипа и
факторов внешней среды в процесс формирования признаков. Метод базируется на том,
что существуют близнецы двух видов: однояйцовые. Причина их появления заключается в
полиэмбрионии, которая зависит от нарушения механических контактов между клетками, в
результате дробящаяся зигота расходится на несколько бластомеров, каждый из которых
дает начало зародышу. Они одинаковых генотипов и их изменчивость зависит от внешних
факторов. Разнояйцовые близнецы. Причиной появления является полиагуляция, которая заключается в овуляции сразу нескольких яйцеклеток в полость малого таза. Генотипы
разные, поэтому изменчивость разнояйцовых близнецов как генотипическая, так и
модификационная. Статистика многоплодных беременностей возрастает после 37 лет, 38
40 рождаемость детей повышается в 5 раз, рост выше среднего, полнота, генетическая
предрасположенность: если женщина из пары близнецов; риск повторной многоплодной
беременности возрастает в 10 раз; в родословной встречаются многоплодные
беременности. Близнецовый метод основан на сравнении изменчивости однояйцовых и
разнояйцовых близнецов. Сходство близнецов описывают термином конкордантность,
различие термином дисконкордантность 100%=%К+%Д.
Можно вывести следующий коэффициент сравнения Н, который лежит в пределах от
0 до 1. Если Н=1, то изменчивость зависит только от внешней среды. Если Н=1, то только
от генотипа. Н=%Ко.б.%Кр.б./100%%Кр.б. Для формы носа Н=1, для разреза глаз Н=0,98,
для туберкулеза Н=0,67. туберкулез развивается только при определенных условиях
среды, но необходима наследственная предрасположенность. Для оксидицита и коклюша
Н=0,3. В близнецовом методе иногда используют метод разлученных близнецов.
Цитологический метод
Это метод визуального анализа строения и поведения хромосом при делении клетки.
Он дает возможность определить нормальные изменения структуры хромосом;
диагностировать хромосомные и геномные мутации; применяется при эволюционных
построениях.
У человека гоплоидное число равно 23. в первую очередь при составлении пар
гомологичных хромосом учитывают их размер. Однако у человека к моменту метафазы
размер хромосом практически одинаковый. Анализируют положение центромера на
хромосоме: центромер находится посередине хромосомы, то есть плечи у такой
хромосомы приблизительно одинаковые. Центромер занимает метацентрическое
положение; центромер несколько сдвинут к концу хромосомы, плечи неравной длинны.
Положение субметоцентрическое; центромер находится на теломерном участке,
следовательно, положение телоцентрическое.
В цитогенетическом методе используют диференцировочную окраску хромосом. При
этом каждая пара гомологичных хромосом имеет свой индивидуальный рисунок. С
помощью дифиренцировочной окраски можно составлять цитогенетические карты
хромосом. Тогда по рисунку можно судить о хромосомных мутациях. С помощью
цитогенетического метода можно определить геномные мутации. Классическим примером
является определение трисомиии по Х. В норме в соматических клетках одна из Х
хромосом конденсируется, превращаясь в тельце Барро, которое прокрашивается в виде
компактного тельца. У женщин с синдромом в ядре заметно два тельца Барро. Показана
определенная гомологичность хромосом человека и обезьяны. Гоплоидное число у обезьян
равно 24, однако было доказано, что одна крупная хромосома человека формировалась за
счет слияния двух мелких обезьяньих.
Молекулярногенетический метод
Является частным случаем биохимического метода. При этом исследуются
нормальная и патологичная структура конечных продуктов биохимических реакций,
количество продуктов, вступающих в реакции, молекулярные структуры полипептидов,
мРНК, ДНК. С помощью этого метода в основном диагностируются генные болезни, причем
наиболее распространенным является исследование на 4 и 5 уровне. По конечным
продуктам диагностируется такая болезнь как альбинизм. По веществам, вступающим в
реакцию, диагностируются галактозимия, фенилкетонурия (при ней в организме
накапливается ядовитый фенилаланин, который не может быть переведен в пигмент). На 4
и 5 уровне в настоящее время диагностируется около 200 генных болезней. На третьем
уровне около 100 генных болезней. На 1 и 2 около 10 генных болезней.
Сравнительнопопуляционный метод
Популяция – это группа особей одного вида, которая характеризуется общими
морфофизиологическими чертами, общими биохимическими параметрами, которые
свободно скрещиваются друг с другом и дают плодовитое потомство и занимают
определенный ареал в природе. Для изучения популяционной генетики человека
используется модель так называемой идеальной популяции. Она характеризуется
следующими параметрами: численность стремиться к бесконечности; наблюдается
свободное скрещивание, то есть панмексия.
Для любой популяции: генотипическая структура популяции или ГСП. Эта
характеристика отражает частоту встречаемости особей различных генотипов в популяции
и изменении этой частоты в ряду поколений. Если исследовать один ген, представленный
двумя аллелями, то в популяции будут встречаться следующие генотипы.
АА
Аа
аа
Д
Н
R
Д+Н+ R =1(100%)
Генофонд популяции или ГФП. Этот параметр отражает частоту встречаемости
различных аллелей в популяции и изменение этой частоты в ряду поколений. Было принято
частоту встречаемости Д—р, R — q
f(A)=p
f(a)=q
тогда зная ГСП популяции, можно определить p и q
p=D+H/2
q=R+H/2
p+q=D+R+2H/2=1
Зная ГФП, можно вычислить ГСП. С этой точки зрения определенно изменяется и
частота аллелей, и частота генотипов в ряду поколений.
Пусть дано:
D 0 +H 0 +R 0 =1 p 0+ q 0=1
в идеальной популяции
Условия панмексии дает возможность изобразить матрицу скрещивания в виде
скрещивания различных аллелей.
A
q o
Aa
ро q o
♀
♂
А
ро
a
q o
А
ро
AA
ро ро
Aa
ро q o
P 1 =D 1 +H 1 /2=p o
2 +2 p o q o /2= p o
p o = p 1
aa
q o q o
2 + p o q o = p o (p o + q o )= p o
частота доминантного аллеля в ряду поколений в идеальной популяции не изменяется.
q 1= R 1+ H 1/2= q o
2+ p o q o = q o ( q o + p o )= q o
2+2 p o q o /2= q o
q 1= q o
в идеальной популяции в ряду поколений частота рецессивного аллеля не изменяется.
q n+1 = q o
p n+1 = p o
Закон ХардиВайнберга
В идеальной популяции частота встречаемости аллелей и генотипов в ряду поколений
остается неизменной. Для расчета частоты аллелей используют формулу ХВ.
p o
2 +2 p o q o + q o
2 =1
за счет закона ХВ можно вычислить частоту встречаемости неблагоприятных рецессивных
аллелей, частоту встречаемости гетерозигот.
Сравнительногенетический метод
Этот метод является частным случаем биологического моделирования и позволяет
изучать заболевания человека, а также действие лекарственных препаратов на других
биологических объектах. Этот закон основан на законе гомологичных рядов
наследственной изменчивости, который был сформулирован Вавиловым. Закон гласит, что
в филогенетически близких группах наблюдаются сходные ряды наследственной
изменчивости, причем это сходство тем полнее, чем родственнее группы организмов. С
этой точки зрения удобнее всего исследовать заболевания человека на обезьянах.
Например, фенилкетонурия изучается на мышах, причем мыши погибают через 10 дней,
сахарный диабет изучается на собаках, гемофилия на собаках и лошадях, ожирение на
свиньях и крысах, атеросклероз на голубях.
Медикогенетическое консультирование
Наследственные болезни – это болезни, которые зависят от изменения генотипа и
находятся под определенными влияниями внешней среды. Наследственные болезни
делятся на две группы: генные; хромосомные (геномные и хромосомные). В лечении
наследственных заболеваний выделяют несколько уровней: симптоматический.
Устраняются симптомы заболевания; патогенетический. Проходит лечение на
биохимическом уровне; этиологический уровень. При помощи метода генных векторов.
Выделяют клетки организма, клонируют их и вводят туда векторные клетки. Важнейшее значение имеет диагностика наследственных болезней, которая
осуществляется в рамках медикогенетического консультирования, основной метод –
генеалогический. Предупреждение осуществляется: неблагоприятной беременности;
принатальная диагностика или на уровне плода. Берут клетки зародыша, выращивают в
культуру и наблюдают аномалии; берут околоплодную жидкость, в которой есть клетки
зародыша, культивируют их.
Р:Аавв ааВВ
F :АаВв
F
2:
А
В
9:
бел
АаВ
3:
Бел
Авв
3:
черн
Аавв
1
кор
Расщепление 12:3:1
Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья
Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья
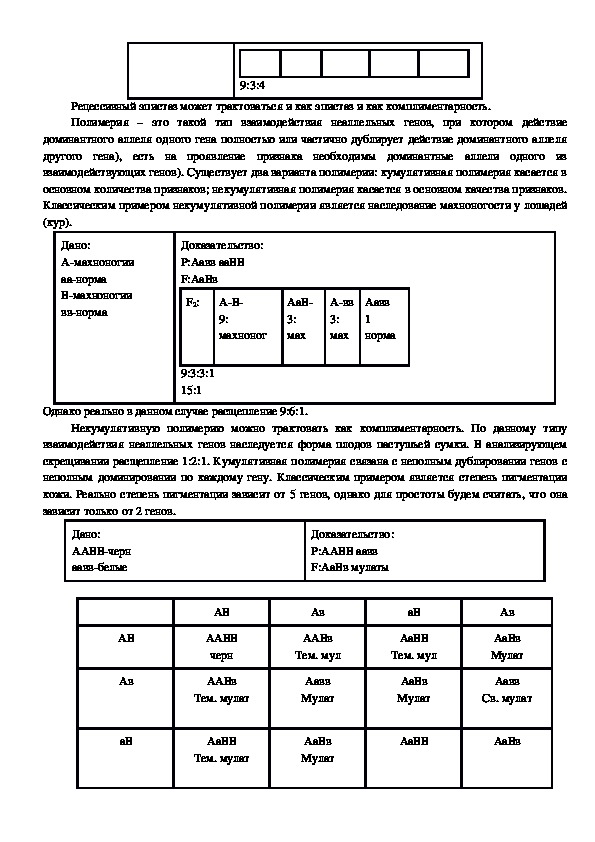
Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья
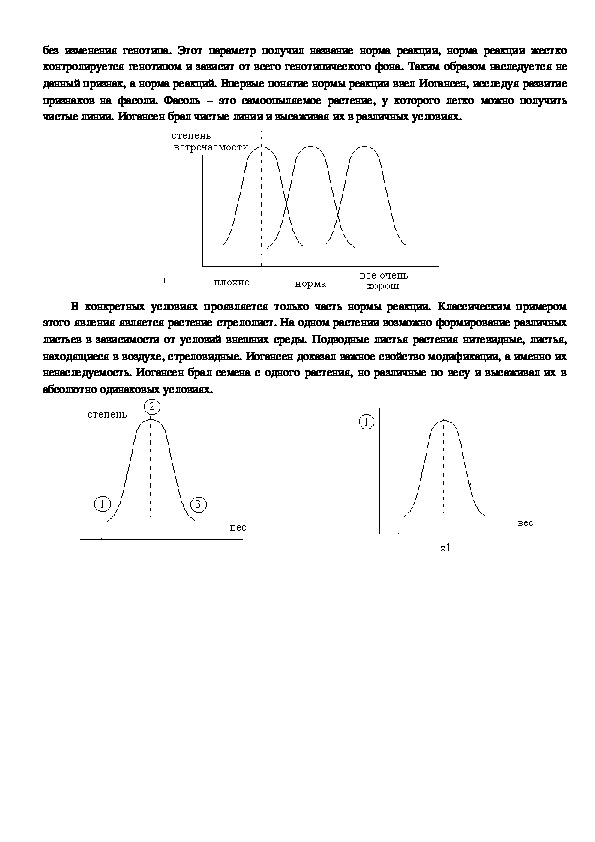
Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья
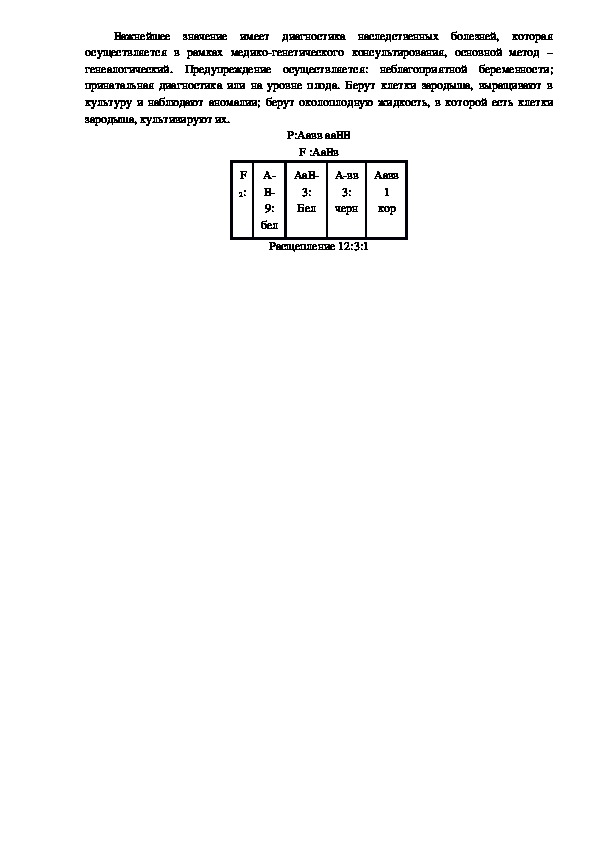
Конспекты лекций по Медико-биологическим основам обучения и воспитания детей с ограниченными возможностями здоровья

Материалы на данной страницы взяты из открытых истончиков либо размещены пользователем в соответствии с договором-офертой сайта. Вы можете сообщить о нарушении.
10.10.2018
Посмотрите также:
© ООО «Знанио»
С вами с 2009 года.
![]()
О портале