Поделиться

Лекция № 1. История развития микробиологии, вирусологии и иммунологии. Предмет, методы, задачи.
1.Введение
Микробиология ( от греч. micros- малый, bios- жизнь, logos- учение, т.е. учение о малых формах жизни) - наука, изучающая организмы, неразличимые (невидимые) невооруженным какой- либо оптикой глазом, которые за свои микроскопические размеры называют микроорганизмы (микробы).
Предметом изучения микробиологии является их морфология, физиология, генетика, систематика, экология и взаимоотношения с другими формами жизни.
В таксономическом отношении микроорганизмы очень разнообразны. Они включают прионы, вирусы, бактерии, водоросли, грибы, простейшие и даже микроскопические многоклеточные животные.
По наличию и строению клеток вся живая природа может быть разделена на прокариоты (не имеющие истинного ядра), эукариоты (имеющие ядро) и не имеющие клеточного строения формы жизни. Последние для своего существования нуждаются в клетках, т.е. являются внутриклеточными формами жизни.
По уровню организации геномов, наличию и составу белоксинтезирующих систем и клеточной стенки все живое делят на 4 царства жизни: эукариоты, эубактерии, архебактерии, вирусы и плазмиды.
К прокариотам, объединяющим эубактерии и архебактерии, относят бактерии, низшие (сине- зеленые) водоросли, спирохеты, актиномицеты, архебактерии, риккетсии, хламидии, микоплазмы. Простейшие, дрожжи и нитчатые грибы- эукариоты.
Микроорганизмы- это невидимые простым глазом представители всех царств жизни. Они занимают низшие (наиболее древние) ступени эволюции, но играют важнейшую роль в экономике, круговороте веществ в природе, в нормальном существовании и патологии растений, животных, человека.
Микроорганизмы заселяли Землю еще 3- 4 млрд. лет назад, задолго до появления высших растений и животных. Микробы представляют самую многочисленную и разнообразную группу живых существ. Микроорганизмы чрезвычайно широко распространены в природе и являются единственными формами живой материи, заселяющими любые, самые разнообразные субстраты (среды обитания), включая и более высокоорганизованные организмы животного и растительного мира.
Можно сказать, что без микроорганизмов жизнь в ее современных формах была бы просто невозможна.
Микроорганизмы создали атмосферу, осуществляют кругоборот веществ и энергии в природе, расщепление органических соединений и синтез белка, способствуют плодородию почв, образованию нефти и каменного угля, выветриванию горных пород, многим другим природным явлениям.
С помощью микроорганизмов осуществляются важные производственные процессы - хлебопечение, виноделие и пивоварение, производство органических кислот, ферментов, пищевых белков, гормонов, антибиотиков и других лекарственных препаратов.
Микроорганизмы как никакая другая форма жизни испытывает воздействие разнообразных природных и антропических (связанных с деятельностю людей) факторов, что, с учетом их короткого срока жизни и высокой скорости размножения, способствует их быстрому эволюционированию.
Наибольшую печальную известность имеют патогенные микроорганизмы (микробы- патогены) - возбудители заболеваний человека, животных, растений, насекомых. Микроорганизмы, приобретающие в процессе эволюции патогенность для человека (способность вызывать заболевания), вызывают эпидемии, уносящие миллионы жизней. До настоящего времени вызываемые микроорганизмами инфекционные заболевания остаются одной из основных причин смертности, причиняют существенный ущерб экономике.
Изменчивость патогенных микроорганизмов составляет основную движущую силу в развитии и совершенствовании систем защиты высших животных и человека от всего чужеродного (чужеродной генетической информации). Более того, микроорганизмы являлись до недавнего времени важным фактором естественного отбора в человеческой популяции ( пример- чума и современное распространение групп крови). В настоящее время вирус иммунодефицита человека (ВИЧ) посягнул на святое святых человека- его иммунную систему.
2. Основные этапы развития микробиологии, вирусологии и иммунологии
1.Эвристический - период эмпирических знаний (до изобретения микроскопов и их применения для изучения микромира).
Дж.Фракасторо (1546г.) предположил живую природу агентов инфекционных заболеваний- contagium vivum.
2.Морфологический период занял около двухсот лет.
Антони ван Левенгук в 1675г. впервые описал простейших, в 1683г.- основные формы бактерий. Несовершенство приборов (максимальное увеличение микроскопов X300) и методов изучения микромира не способствовало быстрому накоплению научных знаний о микроорганизмах.
3.Физиологический период (с 1875г.)- эпоха Л.Пастера и Р.Коха.
Л.Пастер- изучение микробиологических основ процессов брожения и гниения, развитие промышленной микробиологии, выяснение роли микроорганизмов в кругообороте веществ в природе, открытие анаэробных микроорганизмов, разработка принципов асептики, методов стерилизации, ослабления (аттенуации) вирулентности и получения вакцин (вакцинных штаммов).
Р.Кох- метод выделения чистых культур на твердых питательных средах, способы окраски бактерий анилиновыми красителями, открытие возбудителей сибирской язвы, холеры (запятой Коха), туберкулеза (палочки Коха), совершенствование техники микроскопии. Экспериментальное обоснование критериев Хенле, известные как постулаты (триада) Хенле- Коха.
4.Иммунологический период.
И.И.Мечников- “поэт микробиологии” по образному определению Эмиля Ру. Он создал новую эпоху в микробиологии - учение о невосприимчивости (иммунитете), разработав теорию фагоцитоза и обосновав клеточную теорию иммунитета.
Одновременно накапливались данные о выработке в организме антител против бактерий и их токсинов, позволившие П.Эрлиху разработать гуморальную теорию иммунитета. В последующей многолетней и плодотворной дискуссии между сторонниками фагоцитарной и гуморальной теорий были раскрыты многие механизмы иммунитета и родилась наука иммунология.
В дальнейшем было установлено, что наследственный и приобретенный иммунитет зависит от согласованной деятельности пяти основных систем : макрофагов, комплемента, Т- и В- лимфоцитов, интерферонов, главной системы гистосовместимости, обеспечивающих различные формы иммунного ответа. И.И.Мечникову и П.Эрлиху в 1908г. была присуждена Нобелевская премия.
12 февраля 1892г. на заседании Российской академии наук Д.И.Ивановский сообщил, что возбудителем мозаичной болезни табака является фильтрующийся вирус. Эту дату можно считать днем рождения вирусологии, а Д.И.Ивановского- ее основоположником. Впоследствии оказалось, что вирусы вызывают заболевания не только растений, но и человека, животных и даже бактерий. Однако только после установления природы гена и генетического кода вирусы были отнесены к живой природе.
5. Следующим важным этапом в развитии микробиологии стало открытие антибиотиков. В 1929г. А.Флеминг открыл пенициллин и началась эра антибиотикотерапии, приведшая к революционному прогрессу медицины. В дальнейшем выяснилось, что микробы приспосабливаются к антибиотикам, а изучение механизмов лекарственной устойчивости привело к открытию второго- внехромосомного (плазмидного) генома бактерий.
Изучение плазмид показало, что они представляют собой еще более просто устроенные организмы, чем вирусы, и в отличии от бактериофагов не вредят бактериям, а наделяют их дополнительными биологическими свойствами. Открытие плазмид существенно дополнило представления о формах существования жизни и возможных путях ее эволюции.
6. Современный молекулярно- генетический этап развития микробиологии, вирусологии и иммунологии начался во второй половине 20 века в связи с достижениями генетики и молекулярной биологии, созданием электронного микроскопа.
В опытах на бактериях была доказана роль ДНК в передаче наследственных признаков. Использование бактерий, вирусов, а затем и плазмид в качестве объектов молекулярно- биологических и генетических исследований привело к более глубокому пониманию фундаментальных процессов, лежащих в основе жизни. Выяснение принципов кодирования генетической информации в ДНК бактерий и установление универсальности генетического кода позволило лучше понимать молекулярно- генетические закономерности, свойственные более высоко организованным организмам.
Расшифровка генома кишечной палочки сделало возможным конструирование и пересадку генов. К настоящему времени генная инженерия создала новые направления биотехнологии.
Расшифрованы молекулярно- генетическая организация многих вирусов и механизмы их взаимодействия с клетками, установлены способность вирусной ДНК встраиваться в геном чувствительной клетки и основные механизмы вирусного канцерогенеза.
Подлинную революцию претерпела иммунология, далеко вышедшая за рамки инфекционной иммунологии и ставшая одной из наиболее важных фундаментальных медико- биологических дисциплин. К настоящему времени иммунология- это наука, изучающая не только защиту от инфекций. В современном понимании иммунология- это наука, изучающая механизмы самозащиты организма от всего генетически чужеродного, поддержании структурной и функциональной целостности организма.
Иммунология в настоящее время включает ряд специализированных направлений, среди которых, наряду с инфекционной иммунологией, к наиболее значимым относятся иммуногенетика, иммуноморфология, трансплантационная иммунология, иммунопатология, иммуногематология, онкоиммунология, иммунология онтогенеза, вакцинология и прикладная иммунодиагностика.
Микробиология и вирусология как фундаментальные биологические науки также включают ряд самостоятельных научных дисциплин со своими целями и задачами: общую, техническую (промышленную), сельскохозяйственную, ветеринарную и имеющую наибольшее значение для человечества медицинскую микробиологию и вирусологию.
Медицинская микробиология и вирусология изучает возбудителей инфекционных болезней человека (их морфологию, физиологию, экологию, биологические и генетические характеристики), разрабатывает методы их культивирования и идентификации, специфические методы их диагностики, лечения и профилактики.
К отдельным наиболее важным разделам медицинской микробиологии и вирусологии можно отнести клиническую микробиологию, санитарную микробиологию, медицинскую микологию и протозоологию, медицинскую паразитологию, учение о сапронозах.
7.Перспективы развития.
На пороге 21 века микробиология, вирусология и иммунология представляют одно из ведущих направлений биологии и медицины, интенсивно развивающееся и расширяющее границы человеческих знаний.
Иммунология вплотную подошла к регулированию механизмов самозащиты организма, коррекции иммунодефицитов, решению проблемы СПИДа, борьбе с онкозаболеваниями.
Создаются новые генно- инженерные вакцины, появляются новые данные об открытии инфекционных агентов - возбудителей “соматических” заболеваний (язвенная болезнь желудка, гастриты, гепатиты, инфаркт миокарда, склероз, отдельные формы бронхиальной астмы, шизофрения и др.).
Появилось понятие о новых и возвращающийся инфекциях (emerging and reemerging infections). Примеры реставрации старых патогенов- микобактерии туберкулеза, риккетсии группы клещевой пятнистой лихорадки и ряд других возбудителей природноочаговых инфекций. Среди новых патогенов- вирус иммунодефицита человека (ВИЧ), легионеллы, бартонеллы, эрлихии, хеликобактер, хламидии (Chlamydia pneumoniae). Наконец, открыты вироиды и прионы - новые классы инфекционных агентов.
Вироиды - инфекционные агенты, вызывающие у растений поражения, сходные с вирусными, однако эти возбудители отличаются от вирусов рядом признаков: отсутствием белковой оболочки (голая инфекционная РНК), антигенных свойств, одноцепочечной кольцевой структурой РНК ( из вирусов - только у вируса гепатита D), малыми размерами РНК.
Прионы (proteinaceous infectious particle- белкоподобная инфекционная частица) представляют лишенные РНК белковые структуры, являющиеся возбудителями некоторых медленных инфекций человека и животных, характеризующихся летальными поражениями центральной нервной системы по типу губкообразных энцефалопатий- куру, болезнь Крейтцфельдта- Якоба, синдром Герстманна- Страусслера- Шайнкера, амниотрофический лейкоспонгиоз, губкообразная энцефалопатия коров (коровье “бешенство”), скрепи у овец, энцефалопатия норок, хроническая изнуряющая болезнь оленей и лосей. Предполагается, что прионы могут иметь значение в этиологии шизофрении, миопатий. Существенные отличия от вирусов, прежде всего отсутствие собственного генома, не позволяют пока рассматривать прионы в качестве представителей живой природы.
3. Задачи медицинской микробиологии.
К ним можно отнести следующие:
1.Установление этиологической (причинной) роли микроорганизмов в норме и патологии.
2.Разработка методов диагностики, специфической профилактики и лечения инфекционных заболеваний, индикации (выявления) и идентификации (определения) возбудителей.
3. Бактериологический и вирусологический контроль окружающей среды, продуктов питания, соблюдения режима стерилизации и надзор за источниками инфекции в лечебных и детских учреждениях.
4.Контроль за чувствительностью микроорганизмов к антибиотикам и другим лечебным препаратам, состоянием микробиоценозов (микрофлорой) повехностей и полостей тела человека.
4.Методы микробиологической диагностики.
Методы лабораторной диагностики инфекционных агентов многочисленны, к основным можно отнести следующие.
1. Микроскопический- с использованием приборов для микроскопии. Определяют форму, размеры, взаиморасположение микроорганизмов, их структуру, способность окрашиваться определенными красителями.
К основным способам микроскопии можно отнести световую микроскопию (с разновидностями- иммерсионная, темнопольная, фазово - контрастная, люминесцентная и др.) и электронную микроскопию. К этим методам можно также отнести авторадиографию (изотопный метод выявления).
2.Микробиологический (бактериологический и вирусологический) - выделение чистой культуры и ее идентификация.
3.Биологический - заражение лабораторных животных с воспроизведением инфекционного процесса на чувствительных моделях (биопроба).
4.Иммунологический ( варианты - серологический, аллергологический) - используется для выявления антигенов возбудителя или антител к ним.
5.Молекулярно- генетический - ДНК- и РНК- зонды, полимеразная цепная реакция (ПЦР) и многие другие.
Заключая изложенный материал, необходимо
отметить теоретическое значение современной микробиологии, вирусологии и
иммунологии. Достижения этих наук позволили изучить фундаментальные процессы
жизнедеятельности на молекулярно- генетическом уровне. Они обусловливают
современное понимание сущности механизмов развития многих заболеваний и
направления их более эффективного предупреждения и лечения.
Лекция № 2. Систематика и морфология микроорганизмов.
1. Систематика микроорганизмов.
Систематика- распределение микроорганизмов в соответствии с их происхождением и биологическим сходством. Систематика занимается всесторонним описанием видов организмов, выяснением степени родственных отношений между ними и объединением их в различные по уровню родства классификационные единицы- таксоны. Основные вопросы, решаемые при систематике (три аспекта, три кита систематики)- классификация, идентификация и номенклатура.
Классификация- распределение (объединение) организмов в соответствии с их общими свойствами (сходными генотипическими и фенотипическими признаками) по различным таксонам.
Таксономия- наука о методах и принципах распределения (классификации) организмов в соответствии с их иерархией. Наиболее часто используют следующие таксономические единицы (таксоны)- штамм, вид, род. Последующие более крупные таксоны- семейство, порядок, класс.
Главной классификационной категорией является вид. В современном представлении вид в микробиологии- совокупность микроорганизмов, имеющих общее эволюционное происхождение, близкий генотип (высокую степень генетической гомологии, как правило более 60%) и максимально близкие фенотипические характеристики.
Нумерическая (численная) таксономия основывается на использовании максимального количества сопоставляемых признаков и математическом учете степени соответствия. Большое число сравниваемых фенотипических признаков и принцип их равной значимости затрудняло классификацию.
При изучении, идентификации и классификации микроорганизмов чаще всего изучают следующие (гено- и фенотипические) характеристики:
1.Морфологические- форма, величина, особенности взаиморасположения, структура.
2.Тинкториальные- отношение к различным красителям (характер окрашивания), прежде всего к окраске по Граму. По этому признаку все микроорганизмы делят на грамположительные и грамотрицательные.
Морфологические свойства и отношение к окраску по Граму позволяют как правило отнести изучаемый микроорганизм к крупным таксонам- семейству, роду.
3.Культуральные- характер роста микроорганизма на питательных средах.
4.Биохимические- способность ферментировать различные субстраты (углеводы, белки и аминокислоты и др.), образовывать в процессе жизнедеятельности различные биохимические продукты за счет активности различных ферментных систем и особенностей обмена веществ.
5.Антигенные- зависят преимущественно от химического состава и строения клеточной стенки, наличия жгутиков, капсулы, распознаются по способности макроорганизма (хозяина) вырабатывать антитела и другие формы иммунного ответа, выявляются в иммунологических реакциях.
6.Физиологические- способы углеводного (аутотрофы, гетеротрофы), азотного (аминоавтотрофы, аминогетеротрофы) и других видов питания, тип дыхания (аэробы, микроаэрофилы, факультативные анаэробы, строгие анаэробы).
7.Подвижность и типы движения.
8.Способность к спорообразованию, характер спор.
9.Чувствительность к бактериофагам, фаготипирование.
10.Химический состав клеточных стенок- основные сахара и аминокислоты, липидный и жинокислотный состав.
11.Белковый спектр (полипептидный профиль).
12.Чувствительность к антибиотикам и другим лекарственным препаратам.
13.Генотипические (использование методов геносистематики).
В последние десятилетия для классификации микроорганизмов, помимо их фенотипических характеристик (см. пп.1- 12), все более широко и эффективно используются различные генетические методы (изучение генотипа- генотипических свойств). Используются все более совершенные методы- рестрикционный анализ, ДНК- ДНК гибридизация, ПЦР, сиквенс и др. В основе большинства методов лежит принцип определения степени гомологии генетического материала (ДНК, РНК). При этом чаще исходят из условного допущения, что степень гомологии более 60% ( для некоторых групп микроорганизмов- 80%) свидетельствует о принадлежности микроорганизмов к одному виду (различные генотипы - один геновид), 40- 60%- к одному роду.
Идентификация.
Основные фено- и генотипические характеристики, используемые для классификации микроорганизмов, используются и для идентификации, т.е. установления их таксономического положения и прежде всего видовой принадлежности- наиболее важного аспекта микробиологической диагностики инфекционных заболеваний. Идентификация осуществляется на основе изучения фено- и генотипических характеристик изучаемого инфекционного агента и сравнения их с характеристиками известных видов. При этой работе часто применяют эталонные штаммы микроорганизмов, стандартные антигены и иммунные сыворотки к известным прототипным микроорганизмам. У патогенных микроорганизмов чаще изучают морфологические, тинкториальные, культуральные, биохимические и антигенные свойства.
Номенклатура- название микроорганизмов в соответствии с международными правилами. Для обозначения видов бактерий используют бинарную латинскую номенклатуру род/вид, состоящую из названия рода (пишется с заглавной буквы) и вида (со строчной буквы). Примеры- Shigella flexneri, Rickettsia sibirica.
В микробиологии часто используется и ряд других терминов для характеристики микроорганизмов.
Штамм- любой конкретный образец (изолят) данного вида. Штаммы одного вида, различающиеся по антигенным характеристикам, называют серотипами (серовариантами- сокращенно сероварами), по чувствительности к специфическим фагам- фаготипами, биохимическим свойствам- хемоварами, по биологическим свойствам- биоварами и т.д.
Колония- видимая изолированная структура при размножении бактерий на плотных питательных средах, может развиваться из одной или нескольких родительских клеток. Если колония развилась из одной родительской клетки, то потомство называется клон.
Культура- вся совокупность микроорганизмов одного вида, выросших на плотной или жидкой питательной среде.
Основной принцип бактериологической работы- выделение и изучение свойств только чистых (однородных, без примеси посторонней микрофлоры) культур.
2. Морфология бактерий.
Прокариоты отличаются от эукариот по ряду основных признаков.
1.Отсутствие истинного дифференцированного ядра (ядерной мембраны).
2.Отсутствие развитой эндоплазматической сети, аппарата Гольджи.
3.Отсутствие митохондрий, хлоропластов, лизосом.
4.Неспособность к эндоцитозу (захвату частиц пищи).
5.Клеточное деление не связано с циклическими изменениями строения клетки.
6. Значительно меньшие размеры (как правило). Большая часть бактерий имеет размеры 0,5- 0,8 микрометров (мкм) х 2- 3 мкм.
Основные формы бактерий:
а – микрококки; б, в – диплококки; г – стрептококки; д - стафилококки; е – тетракокки; ж – сарцины; з, и, к - палочки; л – вибрионы; м – спириллы
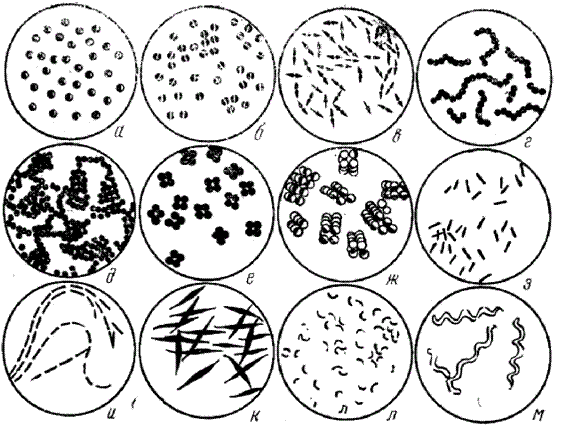
По форме выделяют следующие основные группы микроорганизмов. I. Шаровидные или кокки ( с греч.- зерно).
II. Палочковидные.
III. Извитые.
IV. Нитевидные
I. Кокковидные бактерии (кокки) (от лат. coccus — зерно) по характеру взаиморасположения после деления подразделяются на ряд вариантов.
1.Микрококки. Клетки расположены в одиночку. Входят в состав нормальной микрофлоры, находятся во внешней среде. Заболеваний у людей не вызывают.
2.Диплококки (от лат. diplodocus — двойной). Деление этих микроорганизмов происходит в одной плоскости, образуются пары клеток. Среди диплококков много патогенных микроорганизмов- гонококк, менингококк, пневмококк.
3.Стрептококки (от лат. streptococcus — витой). Деление осуществляется в одной плоскости, размножающиеся клетки сохраняют связь (не расходятся), образуя цепочки. Много патогенных микроорганизмов - возбудители ангин, скарлатины, гнойных воспалительных процессов.
4.Тетракокки. Деление в двух взаимоперпендикулярных плоскостях с образованием тетрад (т.е. по четыре клетки). Медицинского значения не имеют.
5.Сарцины (от лат. tetra— четыре). Деление в трех взаимоперпендикулярных плоскостях, образуя тюки (пакеты) из 8, 16 и большего количества клеток. Часто обнаруживают в воздухе. Болезнетворных форм нет.
6.Стафилококки (от лат.- staphylococcus - гроздь винограда). Делятся беспорядочно в различных плоскостях, образуя скопления, напоминающие грозди винограда. Вызывают многочисленные болезни, прежде всего гнойно- воспалительные.
II. Палочковидные формы микроорганизмов.
Средние размеры от 1 до 6 мкм в длину и 0,5—2 мкм в ширину.
К бактериям относятся палочковидные микроорганизмы, как правило, не образующие спор (кишечная палочка, брюшнотифозная, паратифозные, дизентерийные, дифтерийные, туберкулезные и др.).
По форме палочковидные бактерии бывают короткими (туляремийная), длинными (сибиреязвенная) с закругленными и заостренными концами.
По взаимному расположению палочковидные формы распределяются на три подгруппы:
1.Бактерии, не образующие спор. (кишечная палочка, паратифозные, дизентерийные, дифтерийные, туберкулезные)
2.Бациллы (от лат. bacillus — палочка) - аэробные спорообразующие микробы. Диаметр споры обычно не превышает размера (“ширины”) клетки (эндоспоры) (сенная, сибиреязвенная, столбнячная, возбудители анаэробной инфекции).
3.Клостридии (от лат. closter — веретено) - анаэробные спорообразующие микробы. Диаметр споры больше поперечника (диаметра) вегетативной клетки, в связи с чем клетка напоминает веретено или теннисную ракетку.
Необходимо иметь в виду, что термин “бактерия” часто используют для обозначения всех микробов- прокариот. В более узком (морфологическом) значении бактерии- палочковидные формы прокариот, не имеющих спор.
По взаимному расположению палочковидные формы распределяются на три подгруппы: диплобактерии и диплобациллы, располагающиеся попарно по длине (бактерии пневмонии); стрептобактерии (возбудитель мягкого шанкра) и стреп-тобациллы (бациллы сибирской язвы);
бактерии и бациллы, которые располагаются без определенной системы (большинство палочковидных форм).
Встречаются бактерии с булавовидными утолщениями на концах — возбудитель дифтерии, некоторые имеют ветвления — микробактерии туберкулеза и лепры.
Общее число палочковидных форм бактерий больше, чем корковидных. III. Извитые формы микроорганизмов.
1.Вибрионы (от лат. vibrio — изгибаюсь) и кампилобактерии- имеют один изгиб, могут быть в форме запятой, короткого завитка.
2.Спириллы (от лат. spira — изгиб) - имеют 2- 3 завитка (оборота спирали). Из патогенных известен один вид spirillum minor — возбудитель содоку, способный вызывать у человека болезнь, передающуюся через укус крыс и других грызунов.
3.Спирохеты (от лат. spirochaeta — бактерия в виде изогнутого длинного винта — штопорообразная форма) - имеют различное число завитков, аксостиль- совокупность фибрилл, специфический для различных представителей характер движения и особенности строения (особенно концевых участков). К патогенным относятся три рода: Treponema, Leptospira, Borrelia.
IV. Нитевидные
Различают два типа нитевидных бактерий: образующие временные нити и постоянные.
Временные нити, иногда с ветвлениями, образуют палочковидные бактерии при нарушении условий их роста или регуляции клеточного деления (микобактерии, коринебактерии, а также риккетсии, микоплазмы, многие грамотрицательные и грамположительные бактерии).
Постоянные нитевидные формы образуются из палочковидных клеток, соединяющихся в длинные цепочки либо с помощью слизи, либо чехлами, либо мостиками. Слизь может связывать отдельные клетки в длинные нити (Zoogloea) или пленки (Bacteriogloea). Нитевидные формы образуют серобактерии и железобактерии.
Следует особо отметить, что бактерии отличаются высоким полиморфизмом (индивидуальной изменчивостью формы, не передающейся по наследству), особенно при культивировании на искусственных питательных средах. Под действием различных факторов (антибиотиков, химических веществ) могут возникать необычные по форме и величине клетки, которые, однако, способны ревертировать в исходное состояние при снятии действия этих факторов. Способность микробов изменяться учитывается в лабораторной диагностике инфекций, при изготовлении биологических препаратов.
3.Строение бактериальной клетки.
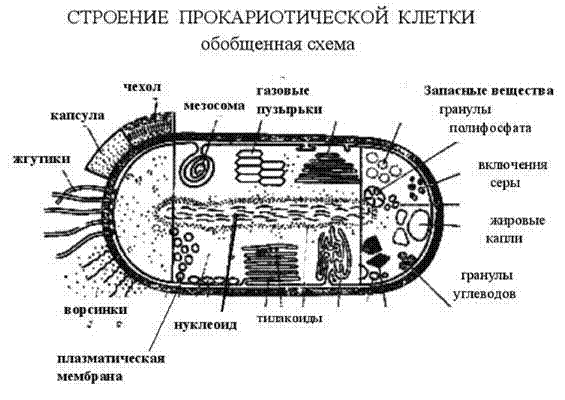
Обязательными органоидами являются: ядерный аппарат, цитоплазма, цитоплазматическая мембрана.
Необязательными (второстепенными) структурными элементами являются: клеточная стенка, капсула, споры, пили, жгутики.
1.В центре бактериальной клетки находится нуклеоид- ядерное образование, представленное чаще всего одной хромосомой кольцевидной формы. Состоит из двухцепочечной нити ДНК. Нуклеоид не отделен от цитоплазмы ядерной мембраной.
2.Цитоплазма- сложная коллоидная система, содержащая различные включения метаболического происхождения (зерна волютина, гликогена, гранулезы и др.), рибосомы и другие элементы белоксинтезирующей системы, плазмиды (вненуклеоидное ДНК), мезосомы (образуются в результате инвагинации цитоплазматической мембраны в цитоплазму, участвуют в энергетическом обмене, спорообразовании, формировании межклеточной перегородки при делении).
3.Цитоплазматическая мембрана ограничивает с наружной стороны цитоплазму, имеет трехслойное строение и выполняет ряд важнейших функций- барьерную (создает и поддерживает осмотическое давление), энергетическую (содержит многие ферментные системы- дыхательные, окислительно- восстановительные, осуществляет перенос электронов), транспортную (перенос различных веществ в клетку и из клетки).
4.Клеточная стенка- присуща большинству бактерий (кроме микоплазм, ахолеплазм и некоторых других не имеющих истинной клеточной стенки микроорганизмов). Она обладает рядом функций, прежде всего обеспечивает механическую защиту и постоянную форму клеток, с ее наличием в значительной степени связаны антигенные свойства бактерий. В составе - два основных слоя, из которых наружный- более пластичный, внутренний- ригидный.
Основное химическое соединение клеточной стенки, которое специфично только для бактерий- пептидогликан (муреиновые кислоты). От структуры и химического состава клеточной стенки бактерий зависит важный для систематики признак бактерий- отношение к окраске по Граму. В соответствии с ним выделяют две большие группы- грамположительные (“грам+”) и грамотрицательные (“грам - “) бактерии. Стенка грамположительных бактерий после окраски по Граму сохраняет комплекс йода с генциановым фиолетовым (окрашены в сине- фиолетовый цвет), грамотрицательные бактерии теряют этот комплекс и соответствующий цвет после обработки и окрашены в розовый цвет за счет докрашивания фуксином.
Особенности клеточной стенки грамположительных бактерий.
Мощная, толстая, несложно организованная клеточная стенка, в составе которой преобладают пептидогликан и тейхоевые кислоты, нет липополисахаридов (ЛПС), часто нет диаминопимелиновой кислоты.
Особенности клеточной стенки грамотрицательных бактерий.
Клеточная стенка значительно тоньше, чем у грамположительных бактерий, содержит ЛПС, липопротеины, фосфолипиды, диаминопимелиновую кислоту. Устроена более сложно- имеется внешняя мембрана, поэтому клеточная стенка трехслойная.
При обработке грамположительных бактерий ферментами, разрушающими пептидогликан, возникают полностью лишенные клеточной стенки структуры- протопласты. Обработка грамотрицательных бактерий лизоцимом разрушает только слой пептидогликана, не разрушая полностью внешней мембраны; такие структуры называют сферопластами. Протопласты и сферопласты имеют сферическую форму (это свойство связано с осмотическим давлением и характерно для всех безклеточных форм бактерий).
|
Гр+ бактерии |
Гр- бактерии |
|
Кокки: стафилококки, стрептококки Палочки (спорообразующие): бациллы, клостридии Палочки (неспорообразующие): коринебактерии, микобактерии, актиномицеты |
Кокки: нейссерии, вейлонеллы; Палочки (неспорообразующие): энтеробактерии, вибрионы; Извитые: спириллы, спирохеты, кампилобактерии. |
L- формы бактерий.
Под действием ряда факторов, неблагоприятно действующих на бактериальную клетку (антибиотики, ферменты, антитела и др.), происходит L- трансформация бактерий, приводящая к постоянной или временной утрате клеточной стенки. L- трансформация является не только формой изменчивости, но и приспособления бактерий к неблагоприятным условиям существования. В результате изменения антигенных свойств (утрата О- и К- антигенов), снижения вирулентности и других факторов L- формы приобретают способность длительно находиться (персистировать) в организме хозяина, поддерживая вяло текущий инфекционный процесс. Утрата клеточной стенки делает L- формы нечувствительными к антибиотикам, антителам и различным химиопрепаратам, точкой приложения которых является бактериальная клеточная стенка. Нестабильные L- формы способны реверсировать в классические (исходные) формы бактерий, имеющие клеточную стенку. Имеются также стабильные L- формы бактерий, отсутствие клеточной стенки и неспособность реверстровать которых в классические формы бактерий закреплены генетически. Они по ряду признаков очень напоминают микоплазмы и другие молликуты- бактерии, у которых клеточная стенка отсутствует как таксономический признак. Микроорганизмы, относящиеся к микоплазмам- самые мелкие прокариоты, не имеют клеточной стенки и как все бактериальные бесстеночные структуры имеют сферическую форму.
К поверхностным структурам бактерий (необязательным, как и клеточная стенка), относятся капсула, жгутики, микроворсинки.
Капсула или слизистый слой окружает оболочку ряда бактерий. Выделяют микрокапсулу, выявляемую при электронной микроскопии в виде слоя микрофибрилл, и макрокапсулу, обнаруживаемую при световой микроскопии. Капсула является защитной структурой (прежде всего от высыхания), у ряда микробов- фактором патогенности, препятствует фагоцитозу, ингибирует первые этапы защитных реакций- распознавание и поглощение. У сапрофитов капсулы образуются во внешней среде, у патогенов- чаще в организме хозяина. Существут ряд методов окраски капсул в зависимости от их химического состава. Капсула чаще состоит из полисахаридов (наиболее распространенная окраска- по Гинсу), реже- из полипептидов.
Жгутики. Подвижные бактерии могут быть скользящие (передвигаются по твердой поверхности в результате волнообразных сокращений) или плавающие, передвигающиеся за счет нитевидных спирально изогнутых белковых (флагеллиновых по химическому составу) образований- жгутиков.
По расположению и количеству жгутиков выделяют ряд форм бактерий.
1.Монотрихи- имеют один полярный жгутик.
2.Лофотрихи- имеют полярно расположенный пучок жгутиков.
3.Амфитрихи- имеют жгутики по диаметрально противоположным полюсам.
4.Перитрихи- имеют жгутики по всему периметру бактериальной клетки.
Способность к целенаправленному движению (хемотаксис, аэротаксис, фототаксис) у бактерий генетически детерминирована.
Фимбрии или реснички - короткие нити, в большом количестве окружающую бактериальную клетку, с помощью которых бактерии прокрепляются к субстратам (например, к поверхности слизистых оболочек). Таким образом, фимбрии являются факторами адгезии и колонизации.
F- пили (фактор фертильности) - аппарат конъюгации бактерий, встречаются в небольшом количестве в виде тонких белковых ворсинок.
Эндоспоры и спорообразование.
Спорообразование- способ сохранения определенных видов бактерий в неблагоприятных условиях среды. Эндоспоры образуются в цитоплазме, представляют собой клетки с низкой метаболической активностью и высокой устойчивостью (резистентностью) к высушиванию, действию химических факторов, высокой температуры и других неблагоплиятных факторов окружающей среды. При световой микроскопии часто используют метод выявления спор по Ожешко. Высокая резистентность связана с большим содержанием кальциевой соли дипиколиновой кислоты в оболочке спор. Расположение и размеры спор у различных микроорганизмов отличается, что имеет дифференциально- диагностическое (таксономическое) значение.
Основные фазы “жизненного цикла” спор- споруляция (включает подготовительную стадию, стадию предспоры, образования оболочки, созревания и покоя) и прорастание, заканчивающееся образованием вегетативной формы. Процесс спорообразования генетически обусловлен.
Некультивируемые формы бактерий.
У многих видов грамотрицательных бактерий, не образующих спор, существует особое приспособительное состояние- некультивируемые формы. Они обладают низкой метаболической активностью и активно не размножаются, т.е. не образуют колоний на плотных питательных средах, при посевах не выявляются. Обладают высокой устойчивостью и могут сохранять жизнеспособность в течение нескольких лет. Не выявляются классическими бактериологическими методами, обнаруживаются только при помощи генетических методов ( полимеразной цепной реакции- ПЦР).
4.Морфологическая характеристика грибов.
Грибы и простейшие имеют четко ограниченное ядро и относятся к эукариотам. Грибы крупнее бактерий, в эволюционном плане близки к растениям (наличие клеточной стенки, содержащей хитин или целлюлозу, вакуолей с клеточным соком, неспособность к перемещению, видимое движение цитоплазмы). Ядерный материал грибов отделен от цитоплазмы ядерной мембраной. Дрожжевые грибы образуют отдельные овальные клетки. Плесневые грибы формируют клеточные нитеподобные структуры- гифы. Мицелий- переплетение гифов- основная морфологическая структура. У низших грибов мицелий одноклеточный, не имеет внутренних перегородок (септ). Грибы размножаются половым и бесполым (вегетативным) способом. При вегетативном размножении образуются специализированные репродуктивные структуры- споры- конидии. Они могут располагаться в специализированных вместилищах- спорангиях (эндоспоры) или отшнуровываться от плодоносящих гиф (экзоспоры). Реже наблюдают образование спор внутри клеток (оидии), являющихся сегментами гиф. Дрожжевые клетки размножаются почкованием, мицелий не образуют. Половое размножение включает взаимодействие специализированных клеток, имеющих существенные различия в морфологии у различных грибов и часто используемых как дифференциально- диагностический признак.
Для большинства видов грибов, имеющих медицинское значение, характерно наличие конидий (или экзоспор), являющихся формами неполового размножения. Их классификация во многом основывается на морфологических формах конидий. Их наиболее частые формы- бластопоры, хламидоспоры, артроспоры, конидиоспоры. Бластоспоры- простые структуры, котрые образуются в результате почкования, с последующим отделением почки от родительской клетки, например у дрожжевых грибов.
Хламидоспоры образуются в результате увеличения гифальных клеток с образованием толстой оболочки, защищающей споры от неблагоприятных условий окружающей среды.
Артроспоры- споры, образующиеся путем фрагментации гиф на отдельные клетки. Они встречаются у дрожжеподобных грибов, возбудителя кокцидиоидоза, тканевых форм дерматофитов в волосе, кожных чешуйках и в ногтях.
Конидиоспоры- зрелые наружные споры, возникающие на дифференцирован-
ных конидиофорах (конидионосцах), отличающихся от других нитей мицелия по форме и размерам (у аспергилл, пеницилл) или располагающиеся по бокам и на концах любой ветви мицелия, прикрепляясь к ней непосредственно или тонкой ножкой.
К эндоспорам совершенных грибов относятся спорангиоспоры мукоровых грибов, развивающихся в специальных органах (спорангиях), располагающихся на вершине спорангиеносца. Споры освобождаются при разрыве стенки спорангия.
Эндоспоры обнаруживают также у тканевых форм возбудителей кокцидиоидоза. Они развиваются в круглых образованиях - сферулах, при разрыве стенки зрелой сферулы попадают во внешнюю среду.
Основное функциональное отличие спор у бактерий и грибов: у бактерий споры обеспечивают переживание в неблагоприятных условиях окружающей среды, у грибов образование спор- способ размножения.
5. Морфологическая характеристика актиномицетов (лучистых грибов по старым классификациям). Актиномицеты - формы бактерий, имеющие истинный, не имеющий перегородок мицелий. Мицелиальный (в виде ветвящихся нитей) рост этих грамположительных бактерий придает им внешнее сходство с грибами. Это сходство усиливается вследствие наличия у высших форм актиномицетов наружных неполовых спор, которые называются конидиями.
В отличие от грибов, актиномицеты имеют прокариотическое строение клетки, не содержат в клеточной стенке хитина или целлюлозы, размножаются только бесполым путем. У низших актиномицетов мицелий фрагментируется на типичные одноклеточные бактерии. Мицелий актиномицетов подразделяют на субстратный (в субстрате) и воздушный. К мицелиальным бактериям относят микобактерии, рода накардий и актиномицетов, несколько родов высших актиномицет.
Представители рода Mycobacterium, в который входят возбудители туберкулеза, являются кислотоустойчивыми микроорганизмами, плохо воспринимающими краски. Их высокая резистентность во внешней среде , кислотоустойчивость и ряд других свойств связан с особым составом клеточной стенки, большим содержанием липидов и воска.
У представителей родов Actinomyces и Nocardia мицелий выражен в значительно большей степени, чем у микобактерий, однако в старых культурах они также проявляют тенденцию фрагментироваться на отдельные клетки неправильной формы. Микроорганизмы рода Actinomyces являются анаэробами, Nocardia - аэробами, многие из которых проявляют кислотоустойчивость.
Микроорганизмы, относящиеся к высшим актиномицетам (рода Streptomyces, Micromonospora) образуют мицелий и размножаются наружными неполовыми спорами или конидиями. Обычным местом обитания для большинства из них является почва. Однако ряд видов актиномицет и нокардий могут инфицировать раны и вызывать образование абсцессов. Для актиномицетов характерно образование друз - плотных “зерен” в гное, представляющих собой беспорядочно переплетенные в центре нити мицелия с радиально отходящими на периферию колбовидно расширенными на концах “дубинками”. С некоторыми актиномицетами (например, стрептомицетами) связана способность выработки антибиотиков.
6. Морфологическая характеристика простейших.
Имеют эукариотическое строение клетки и
значительно более сложную функциональную и морфологическую организацию по
сравнению с бактериями и грибами. Снаружи тело простейших покрывает эластичная
и ригидная пелликула, образованная внешним слоем цитоплазмы. У некоторых
видов клеточная мембрана может включать опорные фибриллы и даже минеральный
скелет. Простейшие могут иметь несколько ядер. Многие простейшие способны
активно двигаться за счет псевдоподий, жгутиков или ресничек. Жизненный цикл
паразитических простейших нередко включает образование промежуточных форм в
различных хозяевах. Основные классы простейших: саркодовые или амебы- наиболее
просто устроенные простейшие, споровики (малярийные плазмодии, токсоплазмы,
пневмоцисты, бабезии), жгутиконосцы (трихомонады, лейшмании), инфузории.
Простейшие очень широко распространены, достаточно сказать, что малярийными
плазмодиями и токсоплазмами в сумме поражено до трети населения земного шара.
Всего известно около 7 тысяч видов простейших, патогенных для различных
растений, животных, человека, непатогенных- во много раз больше. Простейших
изучает наука протозоология.
Лекция № 3. Химическая структура, биохимические свойства и ферменты бактерий.
Клетка- универсальная единица живой материи. По химическому составу существенных отличий прокариотических и эукариотических клеток нет.
Химические элементы, входящие в состав живой материи, можно разделить на три основные группы.
1.Биогенные химические элементы (С, О, N, H). На их долю приходится 95% сухого остатка, в т.ч. 50%- C, 20%- O, 15%- N, 10%- H).
2.Макроэлементы- P, S,Cl, K, Mg, Ca, Na. На них приходится около 5 %.
3.Микроэлементы- Fe, Cu, I, Co, Mo и др. На них приходятся доли процента, однако они имеют важное значение в обменных процессах.
Химические элементы входят в состав различных веществ- воды, белков, липидов, нейтральных жиров, углеводов, нуклеиновых кислот. Синтез соединений контролируется генами. Многие вещества бактериальная клетка может получать извне- из окружающей среды или организма хозяина.
Вода составляет от 70 до 90 % биомассы. Содержание воды больше у капсульных бактерий, меньше всего- в спорах.
Белки встречаются во всех структурных элементах клетки. Белки могут быть более простые (протеины) и сложные (протеиды), в чистом виде или в комплексе с липидами, сахарами. Выделяют структурные (структурообразующие) и функциональные (регуляторные) белки, к последним относятся ферменты.
В состав белков входят как обычные для эукариотов аминокислоты, так и оригинальные- диаминопимелиновая, D-аланин, D-глютанин, входящие в состав пептидогликанов и капсул некоторых бактерий. Только в спорах находится дипиколиновая кислота, с которой связана высокая резистентность спор. Жгутики построены из белка флагеллина, обладающего сократительной способностью и выраженными антигенными свойствами. Пили (ворсинки) содержат особый белок- пилин.
Пептидную природы имеют капсулы представителей рода Bacillus, возбудителя чумы, поверхностные антигены ряда бактерий, в том числе стафилококков и стрептококков. Белок А - специфический белок S.aureus - фактор, обусловлавливающий ряд свойств этого возбудителя. Белок М - специфический белок гемолитических стрептококков серогруппы А, позволяющий дифференцировать серовары (около 100), что имеет эпидемиологическое значение.
Ряд белков содержит наружная мембрана грамотрицательных бактерий, из которых 3 - 4 мажорных (основных) и более 10- второстепенных, выполняющих различные функции. Среди мажорных белков- порины, образующие диффузные поры, через которые в клетку могут проникать мелкие гидрофильные молекулы.
Белки входят в состав пептидогликана- биополимера, составляющего основу бактериальной клеточной стенки. Он состоит из остова (чередующиеся молекулы двух аминосахаров) и двух наборов пептидных цепочек- боковых и поперечных. Наличие двух типов связей- гликозидных (между аминосахарами) и пептидных, которые соединяют субъединицы пептидогликанов, придают этому гетерополимеру структуру молекулярной сети. Пептидогликан- наиболее устойчивое соединение, которое образует ригидную мешковидную макромолекулу, определяющую постоянную форму бактерий и ряд их свойств.
1.Пептидогликан содержит родо- и видоспецифические антигенные детерминанты.
2.Он запускает классический и альтернативный пути активации системы комплемента.
3.Пептидогликан тормозит фагоцитарную активность и миграцию макрофагов.
4.Он способен инициировать развитие гиперчувствительности замедленного типа (ГЗТ).
5.Пептидогликан обладает противоопухолевым действием.
6.Он оказывает пирогенное действие, т.е. вызывает лихорадку.
Из соединений белков с небелковыми компонентами наибольшее значение имеют липопротеиды, гликопротеиды и нуклеопротеиды.
Удивительное таинство жизни- синтез белка осуществляется в рибосомах. Существует два основных типа рибосом - 70S (S- константа седиментации, единица Сведберга) и 80S. Рибосомы первого типа встречаются только у прокариотов. Антибиотики не действуют на синтез белка в рибосомах типа 80S, распространенных у эукариотов.
Липиды (главным образом форфолипиды) содержатся в цитоплазматической мембране (липидный бислой), в также в наружной мембране грамотрицательных бактерий. Есть микроорганизмы, содержащие большое количество липидов (до 40% сухого остатка)- микобактерии. В состав липидов входят различные жирные кислоты, весьма специфичные для разных групп микроорганизмов. Их определение имеет в ряде случаев диагностическое значение, например у анаэробов, микобактерий.
У микобактерий туберкулеза в составе липидов имеется ряд кислотоустойчивых жирных кислот- фтионовая, миколовая и др. Высокое содержание липидов и их состав определяют многие свойства микобактерий туберкулеза:
-устойчивость к кислотам, щелочам и спиртам;
-трудная окрашиваемость красителями (используют специальные методы окраски, чаще- по Цилю- Нильсену);
-устойчивость возбудителя к солнечной радиации и дезосредствам; - патогенность.
Тейхоевые кислоты встречаются в клеточных стенках грамположительных бактерий. Представляют собой водорастворимые линейные полимеры, содержащие остатки глицерина или рибола, связанные фосфодиэфирными связыми. С тейхоевыми кислотами связаны главные поверхностные антигены ряда грамположительных бактерий.
Углеводы встречаются чаще в виде полисахаридов, кторые могут быть экзо- и эндоклеточными. Среди экзоклеточных полисахаридов выделяют каркасные (входят в состав капсул) и истинно экзополисахариды (выходят во внешнюю среду). Среди бактериальных полисахаридов многие находят медицинское применение. Декстраны- полисахариды с большой молекулярной массой, по виду напоминают слизь. 6% раствор- кровезаменитель полиглюкин. Декстрановый гель сефадекс используется в колоночной хроматографии как молекулярное сито. Эндоклеточные полисахариды- запасные питательные вещества клетки (крахмал, гликоген и др.).
Липополисахарид (ЛПС) - один из основных компонентов клеточной стенки грамотрицательных бактерий, это соединение липида с полисахаридом. ЛПС состоит из комплекса: 1.Липид А.
2.Одинаковое для всех грамотрицательных бактерий полисахаридное ядро.
3.Терминальная сахаридная цепочка (О- специфическая боковая цепь). Синонимы ЛПС- эндотоксин, О- антиген.
ЛПС выполняет две основные функции- определяет антигенную специфичность и является одним из основных факторов патогенности. Это- эндотоксин, токсические свойства которого проявляются преимущественно при разрушении бактериальных клеток. Его токсичность определяется липидом А. ЛПС запускает синтез более 20 биологически активных веществ, определяющих патогенез эндотоксикоза, обладает пирогенным действием.
Нуклеиновые кислоты- ДНК и РНК. Рибонуклеиновые кислоты (РНК) находятся главным образом в рибосомах (р-РНК- 80- 85%), т(транспортные)- РНК- 10%, м(матричные)- РНК- 1- 2%, главным образом в одноцепочечной форме. ДНК (дезоксирибонуклеиновая кислота) может находиться в ядерном аппарате (хромосомная ДНК) или в цитоплазме в специализированных образованиях- плазмидах- плазмидная (внехромосомная) ДНК. Микроорганизмы отличаются по структуре нуклеиновых кислот, содержанию азотистых оснований. Генетический код состоит всего из четырех букв (оснований) - А (аденин), Т (тимин), Г (гуанин) и Ц (цитозин). Наиболее часто для характеристики микроорганизмов используют как таксономический признак процентное соотношение Г/Ц, которое существенно отличается у различных групп микроорганизмов.
Микроорганизмы синтезируют различные ферменты- специфические белковые катализаторы. У бактерий обнаружены ферменты 6 основных классов.
1.Оксидоредуктазы- катализируют окислительно- восстановительные реакции.
2.Трансферазы- осуществляют реакции переноса групп атомов.
3.Гидролазы- осущесвляют гидролитическое расщепление различных соединений.
4.Лиазы- катализируют реакции отщепления от субстрата химической группы негидролитическим путем с образованием двойной связи или присоединения химической группы к двойным связям.
5.Лигазы или синтетазы- обеспечивают соединение двух молекул, сопряженное с расщеплением пирофосфатной связи в молекуле АТФ или аналогичного трифосфата.
6.Изомеразы - определяют пространственное расположение групп элементов.
В соответствии с механизмами генетического контроля у бактерий выделяют три группы ферментов:
- конститутивные, синтез которых происходит постоянно;
- индуцибельные, синтез которых индуцируется наличием субстрата;
- репрессибельные, синтез которых подавляется избытком продукта реакции.
Ферменты бактерий делят на экзо- и эндоферменты. Экзоферменты выделяются во внешнюю среду, осуществляют процессы расщепления высокомолекулярных органических соединений. Способность к образованию экзоферментов во многом определяет инвазивность бактерий- способность проникать через слизистые, соединительнотканные и другие тканевые барьеры.
Примеры: гиалуронидаза расщепляет гиалуроновую кислоту, входящую в со-
став межклеточного вещества, что повышает проницаемость тканей (клостридии, стрептококки, стафилококки и многие другие микроорганизмы); нейраминидаза облегчает преодоление слоя слизи, проникновение внутрь клеток и распространение в межклеточном пространстве (холерный вибрион, дифтерийная палочка, вирус гриппа и многие другие). К этой же группе относятся энзимы, разлагающие антибиотики.
В бактериологии для дифференциации микроорганизмов по биохимическим свойствам основное значение часто имеют конечные продукты и результаты действия ферментов. В соответствии с этим существует микробиологическая (рабочая) классификация ферментов.
1.Сахаролитические.
2.Протеолитические.
3.Аутолитические.
4.Окислительно- восстановительные.
5.Ферменты патогенности (вирулентности).
Ферментный состав клетки определяется геномом и является достаточно постоянным признаком. Знание биохимических свойств микроорганизмов позволяет идентифицировать их по набору ферментов. Основные продукты ферментирования углеводов и белков- кислота, газ, индол, сероводород, хотя реальный спектр для различных микроорганизмов намного более обширный.
Основные ферменты вирулентности- гиалуронидаза, плазмокоагулаза, лецитиназа, нейраминидаза, ДНК-аза. Определение ферментов патогенности имеет значение при идентификации ряда микроорганизмов и выявления их роли в патологии. Ряд ферментов микроорганизмов широко используется в медицине и биологии для получения различных веществ (аутолитические, протеолитические), в генной инженерии (рестриктазы, лигазы).
Лекция № 4. Физиология и принципы культивирования микроорганизмов.
Метаболизм микроорганизмов.
Для роста и размножения микроорганизмы нуждаются в веществах, используемых для построения структурных компонентов клетки и получения энергии. Метаболизм (т.е. обмен веществ и энергии) имеет две составляющих- анаболизм и катаболизм. Анаболизм- синтез компонентов клетки (конструктивный обмен). Катаболизм- энергетический обмен, связан с окислительно- восстановительными реакциями, расщеплением глюкозы и других органических соединений, синтезом АТФ. Питательные вещества могут поступать в клетку в растворимом виде (это характерно для прокариот)- осмотрофы, или в виде отдельных частиц- фаготрофы.
Основным регулятором поступления веществ в бактериальную клетку является цитоплазматическая мембрана. Существует четыре основных механизма поступления веществ: -пассивная диффузия- по градиенту концентрации, энергонезатратная, не имеющая субстратной специфичности;
- облегченная диффузия- по градиенту концентрации, субстратспецифичная, энергонезатратная, осуществляется при участии специализированных белков пермеаз;
- активный транспорт- против градиента концентрации, субстратспецифичен (специальные связывающие белки в комплексе с пермеазами), энергозатратный (за счет АТФ), вещества поступают в клетку в химически неизмененном виде;
- транслокация (перенос групп)- против градиента концентрации, с помощью фосфотрансферазной системы, энергозатратна, вещества (преимущественно сахара) поступают в клетку в форфорилированном виде.
Основные химические элементы- органогены, необходимые для синтеза органичеких соединений- углерод, азот, водород, кислород. В зависимости от источника потребляемого углерода микробы подразделяют на аутотрофы (используют CO2) и гетеротрофы (используют готовые органические соединения). В зависимости от источника энергии микроорганизмы делят на фототрофы (энергию получают за счет фотосинтеза- например, цианобактерии) и хемотрофы (энергия добывается за счет химических, окислительно- восстановительных реакций). Если при этом донорами электронов являются неорганические соединения, то это литотрофы, если органические- органотрофы. Если бактериальная клетка в состоянии синтезировать все необходимые для жизнедеятельности вещества, то это прототрофы. Если бактерии нуждаются в дополнительных веществах (факторах роста), то это ауксотрофы. Основными факторами роста для труднокультивируемых бактерий являются пуриновые и пиримидиновые основания , витамины, некоторые (обычно незаменимые) аминокислоты, кровяные факторы (гемин) и др.
Дыхание микроорганизмов.
Путем дыхания микроорганизмы добывают энергию. Дыхание- биологический процесс переноса электронов через дыхательную цепь от доноров к акцепторам с образованием АТФ. В зависимости от того, что является конечным акцептором электронов, выделяют аэробное и анаэробное дыхание. При аэробном дыхании конечным акцептором электронов является молекулярный кислород (О2), при анаэробном- связанный кислород ( -NO3 , =SO4, =SO3).
Примеры.
О2
![]() Аэробное
дыхание донор водорода H2O
Аэробное
дыхание донор водорода H2O
Анаэробное дыхание
нитратное окисление NO3
(факультативные анаэробы) донор водорода N2
 сульфатное
окисление SO4
(облигатные анаэробы) донор водорода H2S
сульфатное
окисление SO4
(облигатные анаэробы) донор водорода H2S
По типу дыхания выделяют четыре группы микроорганизмов.
1.Облигатные (строгие) аэробы. Им необходим молекулярный (атмосферный) кислород для дыхания.
2.Микроаэрофилы нуждаются в уменьшенной концентрации (низком парциальном давлении) свободного кислорода. Для создания этих условий в газовую смесь для культивирования обычно добавляют CO2, например до 10- процентной концентрации.
3.Факультативные анаэробы могут потреблять глюкозу и размножаться в аэробных и анаэробных условиях. Среди них имеются микроорганизмы, толерантные к относительно высоким (близких к атмосферным) концентрациям молекулярного кислорода - т.е. аэротолерантные, а также микроорганизмы которые способны в определенных условиях переключаться с анаэробного на аэробное дыхание.
4.Строгие анаэробы размножаются только в анаэробных условиях т.е. при очень низких концентрациях молекулярного кислорода, который в больших концентрациях для них губителен. Биохимически анаэробное дыхание протекает по типу бродильных процессов, молекулярный кислород при этом не используется.
Аэробное дыхание энергетически более эффективно (синтезируется большее количество АТФ).
В процессе аэробного дыхания образуются токсические продукты окисления (H2O2- перекись водорода, -О2 - свободные кислородные радикалы), от которых защищают специфические ферменты, прежде всего каталаза, пероксидаза, пероксиддисмутаза. У анаэробов эти ферменты отсутствуют, также как и система регуляции окислительно- восстановительного потенциала (rH2).
Основные методы создания анаэробных условий для культивирования микроорганизмов.
1.Физический- откачивание воздуха, введение специальной газовой безкислородной смеси (чаще- N2- 85%, CO2- 10%, H2- 5%). 2.Химический- применяют химические поглотители кислорода.
3.Биологический- совместное культивирование строгих аэробов и анаэробов (аэробы поглощают кислород и создают условия для размножения анаэробов).
4.Смешанный- используют несколько разных подходов.
Необходимо отметить, что создание оптимальных условий для строгих анаэробов- очень сложная задача. Очень непросто обеспечить постоянное поддержание безкислородных условий культивирования, необходимы специальные среды без содержания растворенного кислорода, поддержание необходимого окислительно- восстановительного потенциала питательных сред, взятие и доставка, посев материала в анаэробных условиях.
Существует ряд приемов, обеспечивающих более подходящие условия для анаэробов- предварительное кипячение питательных сред, посев в глубокий столбик агара, заливка сред вазелиновым маслом для сокращения доступа кислорода, использование герметически закрывающихся флаконов и пробирок, шприцев и лабораторной посуды с инертным газом, использование плотно закрывающихся эксикаторов с горящей свечой. Используются специальные приборы для создания анаэробных условий- анаэростаты. Однако в настоящее время наиболее простым и эффективным оборудованием для создания анаэробных и микроаэрофильных условий является система “Газпак” со специальными газорегенерирующими пакетами, действующими по принципу вытеснения атмосферного воздуха газовыми смесями в герметически закрытых емкостях.
Основные принципы культивирования микроорганизмов на питательных средах.
1.Использование всех необходимых для соответствующих микробов питательных компонентов.
2.Оптимальные температура, рН, rH2, концентрация ионов, степень насыщения кислородом, газовый состав и давление.
Микроорганизмы культивируют на питательных средах при оптимальной температуре в термостатах, обеспечивающих условия инкубации.
По температурному оптимуму роста выделяют три основные группы микроорганизмов.
1.Психрофилы- растут при температурах ниже +20 градусов Цельсия.
2.Мезофилы- растут в диапозоне температур от 20 до 45 градусов (часто оптимум- при 37 градусах С).
3.Термофилы- растут при температурах выше плюс 45 градусов.
Краткая характеристика питательных сред.
По консистенции выделяют жидкие, плотные (1,5- 3% агара) и полужидкие (0,3- 0,7 % агара) среды.
Агар- полисахарид сложного состава из морских водорослей, основной отвердитель для плотных (твердых) сред. В качестве универсального источника углерода и азота применяют пептоны- продукты ферментации белков пепсином, различные гидролизаты- мясной, рыбный, казеиновый, дрожжевой и др.
По назначению среды разделяют на ряд групп:
- универсальные (простые), пригодные для различных нетребовательных микроорганизмов (мясо- пептонный бульон- МПБ, мясо- пептонный агар- МПА);
- специальные- среды для микроорганизмов, не растущих на универсальных средах (среда Мак- Коя на туляремию, среда Левенштейна- Иенсена для возбудителя туберкулеза);
- дифференциально- диагностические- для дифференциации микроорганизмов по ферментативной активности и культуральным свойствам ( среды Эндо, Плоскирева, Левина, Гисса);
- селективные (элективные)- для выделения определенных видов микроорганизмов и подавления роста сопутствующих- пептонная вода, селенитовая среда, среда Мюллера.
По происхождению среды делят на естественные, полусинтетические и синтетические.
Рост и размножение микроорганизмов.
Бактериальные клетки размножаются в результате деления. Основные стадии размножения микробов в жидкой среде в стационарных условиях:
- лаг- фаза (начальная стадия адаптации с медленным темпом прирости биомассы бактерий);
- экспоненциальная (геометрического роста) фаза с резким ростом численности популяции микроорганизмов (2 в степеии n);
- стационарная фаза (фаза равновесия размножения и гибели микробных кле-
ток);
- стадия гибели - уменьшение численности популяции в связи с уменьшением и отсутствием условий для размножения микроорганизмов (дефицит питательных веществ, изменение рH, rH2, концентрации ионов и других условий культивирования).
Данная динамика характерна для периодических культур с постепенным истощением запаса питательных веществ и накоплением метаболитов.
Если в питательной среде создают условия для поддержания микробной популяции в экспоненциальной фазе- это хемостатные (непрерывные) культуры.
Характер роста бактерий на плотных и жидких питательных средах: сплошной рост, образование колоний, осадок, пленка, помутнение.
Чистая культура- популяция одного вида микроорганизмов.
Основные принципы получения чистых культур: механическое разобщение, рассев, серийные разведения, использование элективных сред, особых условий культивирования (с учетом устойчивости некоторых микробов к определенным температурам, кислотам, щелочам, парциальному давлению кислорода, рН и мн.др).
Лекция № 5. Общая вирусология. Классификация, структура и особенности биологии вирусов. Бактериофаги.
Открытие вирусов Д.И.Ивановским в 1892г. положило начало развитию науки вирусологии. Более быстрому ее развитию способствовали: изобретение электронного микроскопа, разработка метода культивирования микроорганизмов в культурах клеток.“Вирус” в переводе с латинского - яд (животного происхождения). Этот термин применяют для обозначения уникальных представителей живой природы, не имеющих клеточного (эукариотического или прокариотического) строения и обладающих облигатным внутриклеточным паразитизмом, т.е. которые не могут жить без клетки.
В настоящее время вирусология- бурно развивающаяся наука, что связано с рядом причин: ведущей ролью вирусов в инфекционной патологии человека (примеры- вирус гриппа, ВИЧ- вирус иммунодефицита человека, цитомегаловирус и другие герпесвирусы) на фоне практически полного отсутствия средств специфической химиотерапии; использованием вирусов для решения многих фундаментальных вопросов биологии и генетики.
Основные свойства вирусов (и плазмид), по которым они отличаются от остального живого мира.
1.Ультрамикроскопические размеры (измеряются в нанометрах). Крупные вирусы (вирус оспы) могут достигать размеров 300 нм, мелкие- от 20 до 40 нм. 1мм=1000мкм, 1мкм=1000нм.
2.Вирусы содержат нуклеиновую кислоту только одного типа- или ДНК (ДНК- вирусы) или РНК (РНК- вирусы). У всех остальных организмов геном представлен ДНК, в них содержится как ДНК, так и РНК.
3.Вирусы не способны к росту и бинарному делению.
4.Вирусы размножаются путем воспроизводства себя в инфицированной клетке хозяина за счет собственной геномной нуклеиновой кислоты.
5.У вирусов нет собственных систем мобилизации энергии и белок- синтензирующих систем, в связи с чем вирусы являются абсолютными внутриклеточными паразитами.
6.Средой обитания вирусов являются живые клетки- бактерии (это вирусы бактерий или бактериофаги), клетки растений, животных и человека.
Все вирусы существуют в двух качественно разных формах: внеклеточной- вирион и внутриклеточной- вирус. Таксономия этих представителей микромира основана на характеристике вирионов- конечной фазы развития вирусов.
Строение (морфология) вирусов.
1.Геном вирусов образуют нуклеиновые кислоты, представленные одноцепочечными молекулами РНК (у большинства РНК- вирусов) или двухцепочечными молекулами ДНК (у большинства ДНК- вирусов).
2.Капсид - белковая оболочка, в которую упакована геномная нуклеиновая кислота. Капсид состоит из идентичных белковых субъединиц- капсомеров. Существуют два способа упаковки капсомеров в капсид- спиральный (спиральные вирусы) и кубический (сферические вирусы).
При спиральной симметрии белковые субъединицы располагаются по спирали, а между ними, также по спирали, уложена геномная нуклеиновая кислота (нитевидные вирусы). При кубическом типе симметрии вирионы могут быть в виде многогранников, чаще всего- двадцатигранники - икосаэдры.
3.Просто устроенные вирусы имеют только нуклеокапсид, т.е. комплекс генома с капсидом и называются “голыми”.
4. У других вирусов поверх капсида есть дополнительная мембраноподобная оболочка, приобретаемая вирусом в момент выхода из клетки хозяина- суперкапсид.
Такие вирусы называют “одетыми”.
Кроме вирусов, имеются еще более просто устроенные формы способных передаваться агентов - плазмиды, вироиды и прионы.
Основные этапы взаимодействия вируса с клеткой хозяина.
1.Адсорбция- пусковой механизм, связанный со взаимодействием специфических рецепторов вируса и хозяина (у вируса гриппа- гемагглютинин, у вируса иммунодефицита человека- гликопротеин gp 120).
2.Проникновение- путем слияния суперкапсида с мембраной клетки или путем эндоцитоза (пиноцитоза).
3.Освобождение нуклеиновых кислот- “раздевание” нуклеокапсида и активация нуклеиновой кислоты.
4.Синтез нуклеиновых кислот и вирусных белков, т.е. подчинение систем клетки хозяина и их работа на воспроизводство вируса.
5.Сборка вирионов- ассоциация реплицированных копий вирусной нуклеиновой кислоты с капсидным белком.
6.Выход вирусных частиц из клетки, приобретения суперкапсида оболочечными вирусами.
Исходы взаимодействия вирусов с клеткой хозяина.
1.Абортивный процесс- когда клетки освобождаются от вируса:
- при инфицировании дефектным вирусом, для репликации которого нужен вирус- помощник, самостоятельная репликация этих вирусов невозможна ( так называемые вирусоиды). Например, вирус дельта (D) гепатита может реплицироваться только при наличии вируса гепатита B, его Hbs - антигена, аденоассоциированный вирус- в присутствии аденовируса);
- при инфицировании вирусом генетически нечувствительных к нему клеток; - при заражении чувствительных клеток вирусом в неразрешающих условиях.
2.Продуктивный процесс- репликация (продукция) вирусов:
- гибель (лизис) клеток (цитопатический эффект)- результат интенсивного размножения и формирования большого количества вирусных частиц - характерный результат продуктивного процесса, вызванного вирусами с высокой цитопатогенностью. Цитопатический эффект действия на клеточные культуры для многих вирусов носит достаточно узнаваемый специфический характер;
- стабильное взаимодействие, не приводящее к гибели клетки (персистирующие и латентные инфекции) - так называемая вирусная трансформация клетки.
3.Интегративный процесс- интеграция вирусного генома с геномом клетки хозяина. Это особый вариант продуктивного процесса по типу стабильного взаимодействия. Вирус реплицируется вместе с геномом клетки хозяина и может длительно находиться в латентном состоянии. Встраиваться в ДНК- геном хозяина могут только ДНК- вирусы (принцип “ДНК- в ДНК”). Единственные РНК- вирусы, способные интегрироваться в геном клетки хозяина- ретровирусы, имеют для этого специальный механизм. Особенность их репродукции- синтез ДНК провируса на основе геномной РНК с помощью фермента обратной транскриптазы с последующим встраиванием ДНК в геном хозяина.
Основные методы культивирования вирусов.
1.В организме лабораторных животных.
2.В куриных эмбрионах.
3.В клеточных культурах - основной метод.
Типы клеточных культур.
1.Первичные (трипсинизированные) культуры- фибробласты эмбриона курицы (ФЭК), человека (ФЭЧ), клетки почки различных животных и т.д. Первичные культуры получают из клеток различных тканей чаще путем их размельчения и трипсинизации, используют однократно, т.е. постоянно необходимо иметь соответствующие органы или ткани.
2.Линии диплоидных клеток пригодны к повторному диспергированию и росту, как правило не более 20 пассажей (теряют исходные свойства).
3.Перевиваемые линии (гетероплоидные культуры), способны к многократному диспергированию и перевиванию, т.е. к многократным пассажам, наиболее удобны в вирусологической работе- например, линии опухолевых клеток Hela, Hep и др.
Специальные питательные среды для культур клеток.
Используются разнообразные синтетические вирусологические питательные среды сложного состава, включающие большой набор различных факторов роста- среда 199, Игла, раствор Хэнкса, гидролизат лактальбумина. В среды добавляют стабилизаторы рН (Hepes), различные в видовом отношении сыворотки крови (наиболее эффективной считают эмбриональную телячью сыворотку), L-цистеин и L-глютамин.
В зависимости от функционального использования среды могут быть ростовые (с большим содержанием сыворотки крови) - их используют для выращивания клеточных культур до внесения вирусных проб, и поддерживающие (с меньшим содержанием сыворотки или ее отсутствием)- для содержания инфицированных вирусом клеточных культур.
Выявляемые проявления вирусной инфекции клеточных культур.
1.Цитопатический эффект.
2.Выявление телец включений.
3. Выявление вирусов методом флюоресцирующих антител (МФА), электронной микроскопией, авторадиографией.
4.Цветная проба. Обычный цвет используемых культуральных сред, содержащих в качестве индикатора рН феноловый красный, при оптимальных для клеток условиях культивирования (рН около 7,2)- красный. Размножение клеток меняет рН и соответственно- цвет среды с красного на желтый за счет смещения рН в кислую сторону. При размножении в клеточных культурах вирусов происходит лизис клеток, изменения рН и цвета среды не происходит.
5.Выявление гемагглютинина вирусов- гемадсорбция, гемагглютинация.
6.Метод бляшек (бляшкообразования). В результате цитолитического действия многих вирусов на клеточные культуры образуются зоны массовой гибели клеток.
Выявляют бляшки- вирусные “ клеточно- негативные” колонии.
Номенклатура вирусов.
Название семейства вирусов заканчивается на “viridae”, рода- “virus”, для вида обычно используют специальные названия, например - вирус краснухи, вирус иммунодефицита человека- ВИЧ, вирус парагриппа человека типа 1 и т.д.
Вирусы бактерий (бактериофаги).
Естественной средой обитания фагов является бактериальная клетка, поэтому фаги распространены повсеместно (например, в сточных водах). Фагам присущи биологические особенности, свойственные и другим вирусам.
Наиболее морфологически распространенный тип фагов характеризуется наличием головки- икосаэдра, отростка (хвоста) со спиральной симметрией (часто имеет полый стержень и сократительный чехол), шипов и отростков (нитей), т.е. внешне несколько напоминают сперматозоид.
Взаимодействие фагов с клеткой (бактерией) строго специфично, т.е. бактериофаги способны инфицировать только определенные виды и фаготипы бактерий.
Основные этапы взаимодействия фагов и бактерий.
1.Адсорбция (взаимодействие специфических рецепторов).
2.Внедрение вирусной ДНК (инъекция фага) осуществляется за счет лизирования веществами типа лизоцима участка клеточной стенки, сокращения чехла, вталкивания стержня хвоста через цитоплазматическую мембрану в клетку, впрыскивание ДНК в цитоплазму.
3.Репродукция фага.
4.Выход дочерних популяций.
Основные свойства фагов.
Различают вирулентные фаги, способные вызвать продуктивную форму процесса, и умеренные фаги, вызывающие редуктивную фаговую инфекцию (редукцию фага). В последнем случае геном фага в клетке не не реплицируется, а внедряется (интегрируется) в хромосому клетки хозяина (ДНК в ДНК), фаг превращается в профаг. Этот процесс получил название лизогении. Если в результате внедрения фага в хромосому бактериальной клетки она приобретает новые наследуемые признаки, такую форму изменчивости бактерий называют лизогенной (фаговой) конверсией. Бактериальную клетку, несущую в своем геноме профаг, называют лизогенной, поскольку профаг при нарушении синтеза особого белка- репрессора может перейти в литический цикл развития, вызвать продуктивную инфекцию с лизисом бактерии.
Умеренные фаги имеют важное значение в обмене генетическим материалом между бактериями- в трансдукции (одна из форм генетического обмена). Например, способностью вырабатывать экзотоксин обладают только возбудитель дифтерии, в хромосому которого интегрирован умеренный профаг, несущий оперон tox, отвечающий за синтез дифтерийного экзотоксина. Умеренный фаг tox вызывает лизогенную конверсию нетоксигенной дифтерийной палочки в токсигенную.
По спектру действия на бактерии фаги разделяют на :
- поливалентные (лизируют близкородственные бактерии, например сальмонеллы);
- моновалентные (лизируют бактерии одного вида);
- типоспецифические (лизируют только определенные фаговары возбудителя).
На плотных средах фаги обнаруживают чаще с помощью spot-теста (образование негативного пятна при росте колоний), методом агаровых слоев (титрования по Грациа).
Практическое использование бактериофагов для:
✓ идентификации (определение фаготипа).
✓ фагопрофилактики (купирование вспышек).
✓ фаготерапии (лечение дисбактериозов).
✓
оценки санитарного состояния окружающей среды и
эпидемиологического анализа.
Лекция № 6. Генетика бактерий и вирусов.
Молекулярная биология, изучающая фундаментальные основы жизни, является в значительной степени детищем микробиологии. В качестве основных объектов изучения в ней используют вирусы и бактерии, а основное направление- молекулярная генетика основана на генетике бактерий и фагов.
Бактерии- удобный материал для генетики. Их отличает:
- относительная простота генома (сопокупности нуклеотидов хромосом);
- гаплоидность (один набор генов), исключающая доминантность признаков;
- различные интегрированные в хромосомы и обособленные фрагменты ДНК; - половая дифференциация в виде донорских и реципиентных клеток; - легкость культивирования, быстрота накопления биомасс.
Общие представления о генетике.
Ген-уникальная структурная единица наследственности, носитель и хранитель жизни. Он имеет три фундаментальные функции.
1.Непрерывность наследственности- обеспечивается механизмом репликации ДНК.
2.Управление структурами и функциями организма - обеспечивается с помощью единого генетического кода из четырех оснований (А- аденин, Т- тимин, Г- гуанин, Ц- цитозин). Код триплетный, поскольку кодон- функциональная единица, кодирующая аминокислоту, состоит из трех оснований (букв).
3.Эволюция организмов- благодаря мутациям и генетическим рекомбинациям.
В узкоспециальном плане ген чаще всего представляет структурную единицу ДНК, расположение кодонов в которой детерминирует первичную структуру соответствующей полипептидной цепи (белка). Хромосома состоит из особых функциональных единиц- оперонов. Основные этапы развития (усложнения) генетической системы можно представить в виде следующей схемы: кодон → ген → оперон → геном вирусов и плазмид → хромосома прокариот (нуклеоид) → хромосомы эукариот (ядро).
Генетический материал бактерий.
1.Ядерные структуры бактерий- хроматиновые тельца или нуклеоиды (хромосомная ДНК). У бактерий одна замкнутая кольцевидная хромосома (до 4 тысяч отдельных генов). Бактериальная клетка гаплоидна, а удвоение хромосомы (репликация ДНК) сопровождается делением клетки. Вегетативная репликация хромосомной (и плазмидной) ДНК обусловливает передачу генетической информации по вертикали- от родительской клетки- к дочерней. Передача генетической информации по горизонтали осуществляется различными механизмами- в результате конъюгации, трансдукции, трансформации, сексдукции.
2.Внехромосомные молекулы ДНК представлены плазмидами, мигрирующими генетическими элементами- транспозонами и инсервационными (вставочными) или IS- последовательностями.
Плазмиды- экстрахромосомный генетический материал (ДНК), более просто устроенные по сравнению с вирусами организмы, наделяющие бактерии дополнительными полезными свойствами. По молекулярной массе плазмиды значительно меньше хромосомной ДНК, содержат от 40 до 50 генов.
Их объединение в одно царство жизни с вирусами связано с наличием ряда общих свойств- отсутствием собственных систем мобилизации энергии и синтеза белка, саморепликацией генома, абсолютным внутриклеточным паразитизмом.
Их выделение в отдельный класс определяется существенными отличиями от вирусов.
1.Среда их обитания- только бактерии (среди вирусов, кроме вирусов бактерий- бактериофагов имеются вирусы растений и животных).
2.Плазмиды сосуществуют с бактериями, наделяя их дополнительными свойствами. У вирусов эти свойства могут быть только у умеренных фагов при лизогении бактерий, чаще же всего вирусы вызывают отрицательный последствия, лизис клеток.
3.Геном представлен двунитевой ДНК.
4.Плазмиды представляют собой “голые” геномы, не имеющие никакой оболочки, их репликация не требует синтеза структурных белков и процессов самосборки.
Плазмиды могут распространяться по вертикали (при клеточном делении) и по горизонтали, прежде всего путем конъюгационного переноса. В зависимости от наличия или отсутствия механизма самопереноса (его контролируют гены tra- оперона) выделяют конъюгативные и неконъюгативные плазмиды. Плазмиды могут встраиваться в хромосому бактерий- интегративные плазмиды или находиться в виде отдельной структуры- автономные плазмиды ( эписомы).
Классификация и биологическая роль плазмид.
Функциональная классификация плазмид основана на свойствах, которыми они наделяют бактерии. Среди них- способность продуцировать экзотоксины и ферменты, устойчивость к лекарственным препаратам, синтез бактериоцинов.
Основные категории плазмид.
1.F- плазмиды - донорские функции, индуцируют деление (от fertility - плодовитость). Интегрированные F - плазмиды- Hfr- плазмиды (высокой частоты рекомбинаций).
2.R- плазмиды (resistance) - устойчивость к лекарственным препаратам.
3.Col- плазмиды- синтез колицинов (бактериоцинов)- факторов конкуренции близкородственных бактерий (антогонизм). На этом свойстве основано колицинотипирование штаммов.
4.Hly- плазмиды- синтез гемолизинов.
5.Ent- плазмиды- синтез энтеротоксинов.
6.Tox- плазмиды- токсинообразование.
Близкородственные плазмиды не способны стабильно сосуществовать, что позволило объединить их по степени родства в Inc- группы (incompatibility- несовместимость).
Биологическая роль плазмид многообразна, в том числе:
- контроль генетического обмена бактерий; - контроль синтеза факторов патогенности; - совершенствование защиты бактерий.
Бактерии для плазмид- среда обитания, плазмиды для них- переносимые между ними дополнительные геномы с наборами генов, благоприятствующих сохранению бактерий в природе.
Мигрирующие генетические элементы - отдельные участки ДНК, способные определять свой перенос между хромосомами или хромосомой и плазмидой с помощью фермента рекомбинации транспозазы. Простейшим их типом являются инсерционные последовательности (IS- элементы) или вставочные элементы, несущие только один ген транспозазы, с помощью которой IS- элементы могут встраиваться в различные участки хромосомы. Их функции- координация взаимодействия плазмид, умеренных фагов, транспозонов и генофора для обеспечения репродукции, регуляция активности генов, индукция мутаций. Величина IS- элементов не превышает 1500 пар оснований.
Транспозоны (Tn- элементы) включают до 25 тысяч пар нуклеотидов, содержат фрагмент ДНК, несущий специфические гены, и два Is- элемента. Каждый транспозон содержит гены, привносящие важные для бактерии характеристики, как и плазмиды (множественная устойчивость к антибиотикам, токсинообразование и т.д.). Транспозоны- самоинтегрирующиеся фрагменты ДНК, могут встраиваться и перемещаться среди хромосом, плазмид, умеренных фагов, т.е. обладают потенциальной способностью распространяться среди различных видов бактерий.
Понятие о генотипе и фенотипе.
Генотип- вся совокупность имеющихся у организма генов.
Фенотип- совокупность реализованных (т.е. внешних) генетически детерминированных признаков, т.е. индивидуальное (в определенных условиях внешней среды) проявление генотипа. При изменении условий существования фенотип бактерий изменяется при сохранении генотипа.
Изменчивость у бактерий может быть ненаследуемой (модификационной) и генотипической (мутации, рекомбинации).
Временные, наследственно не закрепленные изменения, возникающие как адаптивные реакции бактерий на изменения окружающей среды, называются модификациями (чаще - морфологические и биохимические модификации). После устранения причины бактерии реверсируют к исходному фенотипу.
Стандартное проявление модификации- распределение однородной популяции на две или более двух типов- диссоциация. Пример- характер роста на питательных средах: S- (гладкие) колонии, R- (шероховатые) колонии, M- (мукоидные, слизистые) колонии, D- (карликовые) колонии. Диссоциация протекает обычно в направлении S→ R. Диссоциация сопровождается изменениями биохимических, морфологических, антигенных и вирулентных свойств возбудителей.
Мутации- скачкообразные изменения наследственного признака. Могут быть спонтанные и индуцированные, генные (изменения одного гена) и хромосомные (изменения двух или более двух участков хромосомы).
Одновременно у бактерий имеются различные механизмы репарации мутаций, в том числе с использованием ферментов- эндонуклеаз, лигаз, ДНК- полимеразы.
Генетические рекомбинации- изменчивость, связанная с обменом генетической информации. Генетические рекомбинации могут осуществляться путем трансформации, трансдукции, конъюгации, слияния протопластов.
1.Трансформация- захват и поглощение фрагментов чужой ДНК и образование на этой основе рекомбинанта.
2.Трансдукция- перенос генетического материала фагами (умеренными фагами- специфическая трансдукция).
3.Конъюгация- при непосредственном контакте клеток. Контролируется tra (transfer) опероном. Главную роль играют конъюгативные F- плазмиды.
Генетика вирусов.
Геном вирусов содержит или РНК, или ДНК (РНК- и ДНК- вирусы соответственно). Выделяют позитивную (+) РНК, обладающую матричной активностью и соответственно- инфекционными свойствами, и негативную ( - ) РНК, не проявляющую инфекционные свойства, которая для воспроизводства толжна транскрибироваться (превращаться) в +РНК. Механизмы репродукции различных вирусов очень сложные и существенно отличаются. Основные их схематические варианты представлены ниже.
1. вирионная (матричная) +РНК → комплементарная -РНК (в рибосомах) → вирионная +РНК.
2. - РНК → вирусная (информационная) +РНК → - РНК (формируется на геноме зараженной клетки).
3. однонитевая ДНК: +ДНК → +ДНК -ДНК → +ДНК -ДНК +ДНК → +ДНК.
4. ретровирусная однонитевая РНК: РНК → ДНК (провирус) → РНК.
5. двунитевая ДНК: разделение нитей ДНК и формирование на каждой комплементарной нити ДНК.
Генофонд вирусов создается и пополняется из четырех основных источников:
двух внутренних (мутации, рекомбинации) и двух внешних (включение в геном
генетического материала клетки хозяина, поток генов из других вирусных популяций).
Комплементация- функциональное взаимодействие двух дефектных вирусов, способствующее их репликации и горизонтальной передаче.
Фенотипическое смешивание- при заражении клетки близкородственными вирусами с образованием вирионов с гибридными капсидами, кодируемыми геномами двух вирусов.
Популяционная изменчивость вирусов связана с двумя разнонаправленными процессами - мутациями и селекцией, связанными с внешней средой как индуктором мутаций и фактором стабилизирующего отбора. Гетерогенность вирусных популяций- адаптационный генетический механизм, способствующий пластичности (устойчивости, приспособляемости) популяций, фактор эволюции и сохранения видов во внешней среде.
Генофонд вирусных популяций сохраняется за счет нескольких механизмов:
- восстановления изменчивости за счет мутаций;
- резервирующих механизмов (возможность перехода любых, даже негативных мутаций в следующую генерацию)- комплементация, рекомбинация;
- буферных механизмов (образование дефектных вирусных частиц, иммунных комплексов и др.), способствующие сохранению вируса в изменяющихся внешних условиях.
Лекция № 7. Медицинская биотехнология и генная инженерия. Микробиологические основы антимикробной профилактики и терапии.
Достижения научно-технического прогресса способствовали развитию новых биологических технологий создания диагностических, лечебных и профилактических препаратов, решению проблем сбалансированности питания, экологических проблем. Основные принципы биотехнологии- ферментация, культивирование микроорганизмов, растительных и животных клеток, генная и клеточная инженерия. На основе достижений генетики разработаны высокоточные методы диагностики и идентификации микроорганизмов- определение плазмидного профиля, рестрикционный анализ, ДНК- гибридизация, полимеразная цепная реакция (ПЦР), секвенирование и мн.др. Методы основаны на использовании ряда специфических ферментов-рестриктаз (ферментов, расщепляющих ДНК в специфических участках), лигаз или синтетаз (обеспечивают соединение двух молекул), в частности ДНК- лигаз (получение рекомбинантных молекул ДНК), полимераз (ДНК- зависимая ДНК- полимераза обеспечивает ПЦР- многократное реплицирование специфического участка нуклеотидной последовательности). Плазмиды (F- плазмиды) и вирусы (бактериофаги) используют в генной инженерии в качестве векторов для переноса генетического материала (генов). Метод клонирования заключается в том, что выделенный фрагмент (ген) вводится в состав плазмиды или другой самореплицирующейся системы и накапливается в размножающихся клетках. Практический вариант использования: микроорганизмы- продуценты биологически активных веществ (в том числе вакцин). Гибридомную технологию используют для получения моноклональных антител (МКА).
Кроме клонирования для получения генов используют секвенирование и химический синтез. С помощью генно- инженерных методов получают вакцины, антигены, диагностикумы, гормоны, иммуномодуляторы. Одним из крупных разделов биотехнологии является производство антибиотиков и различных химиотерапевтических препаратов антибактериального действия.
Методы воздействия на микроорганизмы по виду использованного фактора можно разделить на физические и химические, по характеру воздействия- на неизбирательные (обеззараживание- дезинфекция, стерилизация) и избирательные (химиотерапевтические).
Физические методы.
1.Термическая обработка- прокаливание, кипячение, пастеризация, автоклавирование.
2.Облучение- ультрафиолетовое, гамма- и рентгеновское, микроволновое.
3.Фильтрование (оптимально- бактериологические фильтры с диаметром 200 нм). Химические методы.
1.Неспецифического действия- дезинфектанты (обработка помещений и др., антисектики- обработка живых тканей). Среди них- препараты йода и хлора, спирты, альдегиды, кислоты и щелочи, соли тяжелых металлов, катионные детергенты, фенолы, окислители, природные препараты- деготь, ихтиол, хлорофиллипт.
2.Избирательно подавляющие жизнедеятельность микроорганизмов- антибиотики и химиотерапевтические препараты.
Эре антибиотикотерапии предшествовал период разработки антимикробных химиопрепаратов. Некоторые вехи: в 1891г. Д.А.Романовский сформулировал основные принципы химиотерапии инфекционных болезней, предложил хинин для лечения малярии, П.Эрлих в 1906г. предложил принцип химической вариации. Синтезированы производные мышьяка сальварсан и неосальварсан, предложен химиотерапевтический индекс. Круг химиопрепаратов постепенно расширялся. В 1932г. открыты подходы к созданию сульфаниламидных препаратов. Однако поистинне революционное значение имело открытие антибиотиков.
Одним из универсальных механизмов антогонизма микроорганизмов является синтез антибиотиков, которые тормозят рост и размножение микроорганизмов (бактериостатическое действие) или убивают их (бактерицидное действие). Антибиотики- вещества, которые могут быть получены из микроорганизмов, растений, животных тканей и синтетическим путем, обладающие выраженной биологической активностью в отношении микроорганизмов. Таких веществ известно несколько тысяч, однако реально используют значительно меньше. Существует ряд требований к антибиотикам, существенно ограничивающих их терапевтическое применение:
- эффективность в низких концентрациях;
- стабильность в организме и в различных условиях хранения;
- низкая токсичность или ее отсутствие;
- выраженный бактериостатический и (или) бактерицидный эффект; - отсутствие выраженных побочных эффектов; - отсутствие иммунодепрессивного воздействия.
Первыми открытыми антибиотиками были пенициллин (Флеминг) и стрептомицин (Ваксман).
Антибиотики могут быть разделены по происхождению, направленности и спектру действия, по механизму действия.
По происхождению антибиотики могут быть:
- бактериального (полимиксин, грамицидин);
- актиномицетного (стрептомицин, левомицетин, эритромицин);
- грибкового (пенициллин);
- растительного (рафанин, фитонциды);
- животного происхождения (интерфероны, лизоцим).
Больше всего известно антибиотиков актиномицетного происхождения. Актиномицеты- преимущественно почвенные микроорганизмы. В условиях большого количества и разнообразия почвенных микроорганизмов их антогонизм, в том числе с помощью выработки антибиотиков- один из механизмов их выживания.
По спектру действия антибиотики разделяют на:
- действующие преимущественно на грамположительную микрофлору- пенициллин, эритромицин;
- действующие преимущественно на грамотрицательную микрофлору- полимиксин; - широкого спектра действия (на грам-плюс и грам-минус флору)- стрептомицин, неомицин;
- противогрибковые- нистатин, амфотеррицин, леварин, низорал;
- противотуберкулезные- стрептомицин, канамицин;
- противоопухолевые- рифампицин;
- противовирусные- интерферон, зовиракс, ацикловир.
Антибиотики разделяют по механизму действия:
- ингибиторы синтеза пептикогликана клеточной стенки (пенициллин, цефалоспорин, ванкомицин, ристомицин). Действуют на имеющих клеточную стенку растущие бактерии, не действуют на L- формы, покоящиеся формы бактерий; - ингибиторы синтеза белка (стрептомицин, левомицетин, тетрациклин);
- ингибиторы синтеза нуклеиновых кислот, пуринов и аминокислот (налидиксовая кислота, рифампицин);
- ингибиторы синтеза мембраны и цитоплазматической мембраны грибов (нистатин, полимиксин).
Побочное действие антибиотиков.
Для макроорганизма:
- токсическое действие;
- дисбактериозы;
- аллергические реакции;
- иммунодепрессивное действие; - эндотоксический шок.
Для микроорганизмов :
- формирование атипичных форм микробов;
- формирование антибиотикорезистентных и антибиотикозависимых форм микроорганизмов.
Биохимические и генетические механизмы лекарственной устойчивости микроорганизмов.
Существует два типа лекарственной устойчивости- естественная (природная) и приобретенная (в результате мутаций, обмена R- плазмидами др.).
Естественная лекарственная устойчивость является видовым признаком, чаще связана с недоступностью антибиотика к его мишени, т.е. невозможностью осуществления его механизма действия. В природных условиях, особенно в почве, микроорганизмы находятся в конкурентной борьбе за субстраты. Антибиотики- один из селективных факторов отбора. Микроорганизмы- продуценты антибиотиков защищены от синтезируемых антибиотиков генетическими механизмами (генетически детерминированная устойчивость, кодируемая в хромосоме или обусловленная наличием R- плазмид). Микроорганизмы в условиях совместного обитания вынуждены вырабатывать устойчивость к антибиотикам.
Резистентность к антибиотикам у микробов может быть связана с негенетическими факторами (низкая метаболическая активность, переход в L- форму).
Основную роль в лекарственной устойчивости принадлежит R- плазмидам, способным передаваться в другие бактерии и формировать своеобразный генофонд лекарственной устойчивости микроорганизмов. Резистентность современных стафилококков к пенициллину доходит до 100%.
На биохимическом уровне в формировании резистентности могут участвовать различные механизмы.
1.Разрушение молекулы антибиотика (пенициллины и другие бета- лактамные антибиотики разрушаются ферментом бета- лактамазой).
2.Модификация структуры молекулы антибиотика, приводящая к утрате биологической активности (так действуют изоферменты).
3.Изменение структуры мишеней, чувствительных к антибиотику (белков 70S рибомос- устойчивость к тетрациклинам, стрептомицину, макролидам, гираз- к хинолонам, рнк- полимераз- к рифампицину, пенициллинсвязывающих белков- транспептидаз- к бета- лактамам).
4.Образование бактериями “обходного” пути метаболизма.
5.Формирование механизмов активного выведения антибиотика из клетки.
Из-за формирования антибиотикоустойчивых популяций микроорганизмов с целью эффективного лечения необходимо предварительно определять чувствительность данного антибиотика к выделенной культуре возбудителя.
Основными методами определения антибиотикочувствительности бактерий in vitro является метод серийных разведений, диффузии в агар (бумажных дисков), определение способности к продукции бета- лактамазы, in vivo- на модели безмикробных животных, определение концентрации антибиотиков в крови и моче.
Метод диффузии в агар с применением стандартных дисков, пропитанных различными антибиотиками в определенных концентрациях (зависят от терапевтической дозы и соотвествуют рекомендациям ВОЗ). Основан на использовании стандартных питательных сред, дисков и методов. Оценка результатов связана с существованием зависимости между размером зоны подавления роста исследуемых культур вокруг дисков и значениями минимальных подавляющих концентраций (МПК)соответствующих антибиотиков (чувствительностью микроорганизмов). Имеются специальные таблицы для оценки результатов, в соответствии с которыми культуры определяют как чувствительные, умеренно устойчивые и устойчивые (резистентные) к тестируемому антибиотику.
Метод серийных разведений антибиотиков позволяет более точно определить МПК, однако из-за громоздкости применяется реже.
Бета-лактамазный тест (определение способности к образованию бета- лактамаз) чаще определяют методом дисков с нитроцефином - цефалоспорином, изменяющим окраску дисков при гидролизе. Положительный тест свидетельствует о резистентности бактерий ко всем бета- лактамаза- чувствительным пенициллинам.
Существует ряд причин, обусловливающих различную чувствительность микроорганизмов к антибиотикам in vitro и in vivo.
На антимикробную активность in vitro влияют многие факторы, в том числе :
- рН среды;
- компоненты среды;
- концентрация микроорганизмов; - условия и время культивирования.
На антимикробную активность препаратов in vivo также влияют различные факторы, из которых необходимо отметить:
- фармакодинамику препарата в организме (скорость всасывания, выведения, расщепления и т.д.);
- локализацию микробов в организме (особенно внутриклеточную локализацию).
Лекция № 8. Экология микроорганизмов.
Микроорганизмы распространены повсюду. Они заселяют почву, воду, воздух, растения, организмы животных и людей- экологические среды обитания микробов.
Выделяют свободноживущие и паразитические микроорганизмы. Всюду, где есть хоть какие- то источники энергии, углерода, азота, кислорода и водорода (кирпичиков всего живого), обязательно встречаются микроорганизмы, различающиеся по своим физиологическим потребностям и занимающих свои экологические ниши. Титаническая роль микроорганизмов в круговороте веществ в природе имеет исключительное значение для поддержания динамического равновесия биосферы. Микроорганизмы в экологических нишах сосуществуют в виде сложных ассоциаций- биоценозов с различными типами взаимоотношений, в конечном счете обеспечивающих сосуществование многочисленных видов прокариот и различных царств жизни. Все типы взаимоотношений микроорганизмов объединяются понятием симбиоз. Он может быть антогонистическим и синэргическим.
Роль микроорганизмов в круговороте веществ в природе.
Под круговоротом веществ в природе понимают циклы превращения химических элементов, из которых построены живые существа, происходящие вследствие разнообразия и гибкости метаболизма микроорганизмов.
Наибольшее значение для всего живого имеет обмен (кругооборот) углерода, кислорода, водорода, азота, серы, фосфора и железа. Этапы кругооборота различных химических элементов осуществляется микроорганизмами разных групп. Непрерывное существование каждой группы зависит от химических превращений элементов, осуществляемых другими группами микроорганизмов. Жизнь на Земле непрерывна, поскольку все основные элементы жизни подвергаются циклическим превращениям, в значительной степени определяемых микроорганизмами.
Микрофлора почвы. Почва является основным местом обитания микробов. Состав микрофлоры складывается из многих тысяч видов бактерий, грибов, простейших и вирусов. Количество микробов зависит от состава почв и ряда других факторов, в одном грамме пахотной почвы может содержаться до 10 млрд. микроорганизмов. Среди них сапрофиты ( “гнилое растение”), т.е. микроорганизмы, живущие за счет мертных органических субстратов. В процессе самоочищения почвы и кругооборота веществ принимают участие также нитрифицирующие, азотфиксирующие, денитрифицирующие и другие группы микроорганизмов. Патогенные микроорганизмы попадают в почву с биовыделениями людей и животных (калом, мочой, мокротой, слюной, гноем, потом и др.), а также с трупами. Дольше всего в почве сохраняются спорообразующие патогенные микроорганизмы- возбудители сибирской язвы, столбняка, газовой гангрены, ботулизма, что определяет эпидемическое значение почвы при этих инфекциях. Возбудители сапронозов могут автономно обитать в почве и воде и быть связанными с почвенными и водными организмами, т.е. эта природная среда обитания для них- основной резервуар возбудителей. Почва и вода в случае сапронозов выступает в качестве источника заражения животных и людей.
Микрофлора воды. Вода- древнейшее место обитания микроорганизмов. Пресноводные водоемы и реки отличаются богатой микрофлорой. Многие виды галофильных микробов обитает в морской воде, в том числе на глубинах в несколько тысяч метров. Численность микроорганизмов в воде в определенной степени связано с содержанием органических веществ. Серьезной экологической проблемой являются сточные воды, содержащие значительное количество микроорганизмов и органических веществ, не успевающих самоочищаться.
Санитарно- гигиеническое качество воды оценивается различными способами. Чаще определяют коли- титр и коли- индекс, а также общее количество микроорганизмов в мл. Коли- индекс- количество E.coli (кишечной палочки) в одном литре, коли- титр- наименьшее количество воды, в котором обнаруживается одна клетка кишечной палочки. Санитарно- эпидемиологическое значение определения в различных объектах микроорганизмов изучает санитарная микробиология. К числу ее основных принципов можно отнести индикацию (выявление) патогенов в объектах окружающей среды, к косвенным методам- выявление санитарно- показательных микроорганизмов, определение общей микробной обсемененности.
Вода имеет существенное значение в эпидемиологии кишечных инфекций. Их возбудители могут попадать с испражнениями во внешнюю среду (почву), со сточными водами- в водоемы и в некоторых случаях- в водопроводную сеть.
Микрофлора воздуха. Воздух как среда обитания менее благоприятен, чем почва и вода- мало питательных веществ, солнечные лучи, высушивание. Главным источником загрязнения воздуха микроорганизмами является почва, меньше- вода. В видовом отношении преобладают кокки (в т.ч. сарцины), споровые бактерии, грибы, актиномицеты. Особое значение имеет микрофлора закрытых помещений (накапливается при выделении через дыхательные пути человека). Воздушно-капельным путем (за счет образования стойких аэрозолей) распространяются многие респираторные инфекции (грипп, коклюш, дифтерия, корь, туберкулез и др.). Микробиологическая чистота воздуха имеет большое значение в больничных условиях (операционные и другие хирургические отделения).
Микрофлора человека и ее значение. Ребенок развивается в организме матери в норме в стерильных условиях. Формирование новой экологической системы “организм человека + населяющая его микрофлора” начинается в момент рождения, причем основой ее является микрофлора матери и окружающей ребенка внешней среды (прежде всего воздуха). В течение короткого времени кожные покровы и слизистые оболочки, сообщающиеся со внешней средой, заселяются разнообразными микроорганизмами. В формировании микрофлоры детей первого года (главным образом- бифидобактерии и лактобактерии) существенную роль имеет естественное (грудное) вскармливание. Нормальная (т.е. в условиях здорового организма) микрофлора в количественном и качественном отношении представлена на различных участках тела (экотопах) неодинаково. Причины- неодинаковые условия обитания.
Аутохтонная (т.е. присущая данной области) микрофлора может быть разделена на резидентную (постоянную) и транзиторную (непостоянную). На слизистых оболочках, особенно желудочно- кишечного тракта, представители нормальной микрофлоры обитают в виде двух форм- часть из них располагается в просвете (просветная), другая заключена в мукозный пристеночный матрикс, образующий биопленку (пристеночная микрофлора).С ней связана колонизационная резистентность кишечника- естественный барьер защиты кишечника (и организма в целом) от инфекционных агентов.
Нормальная микрофлора кожи. Наиболее заселены микроорганизмами места, защищенные от действия света и высыхания. Наиболее постоянен состав микрофлоры в области устьев сально- волосяных фолликулов. Чаще выявляют Staphylococcus epidermidis и S.saprophyticus, грибы рода Candida, реже- дифтероиды и микрококки.
Микрофлора дыхательных путей. Слизистые оболочки гортани, трахеи, бронхов и альвеолы здорового человека не содержат микроорганизмов. Основная масса микрофлоры рото- и носоглотки приходится на зеленящего стрептококка, реже выявляются нейссерии, дифтероиды и стафилококки.
Микрофлора мочеполового тракта. Микробный биоценоз скуден, верхние отделы обычно стерильны. Во влагалище здоровой женщины преобладают молочнокислые палочки Додерлейна (лактобактерии), создающие кислую рН, угнетающую рост грамотрицательных бактерий и стафилококков, и дифтероиды. Существует баланс между лактобактериями с одной стороны и гарднереллами и анаэробами с другой.
Микрофлора желудочно- кишечного
тракта. Наиболее активно бактерии обживают желудочно- кишечный тракт. При
этом колонизация осуществляется четко “по этажам”. В желудке с кислой реакцией
среды и верхних отделов тонкой кишки количество микроорганизмов не превышает
1000 в мл, чаще обнаруживают лактобациллы, энтерококки, дрожжи, бифидобактерии,
E.coli. Микрофлора толстого кишечника наиболее стабильна и многообразна. Это
поистинне резервуар бактерий всего организма- обнаружено более 250 видов, общая
биомасса микробов может достигать 1,5 кг. Доминирующей группой в норме являются
бесспоровые анаэробные бактерии (бифидобактерии и бактероиды)- до 99%. Выделяют
мукозную (пристеночную) и просветную микрофлору. Пристеночная микрофлора
обеспечивает колонизационную резистентность кишечника, играющую важную роль в
предупреждении (в норме) и в развитии (при патологии) экзо- и эндогенных
инфекционных заболеваний. Нормальная микрофлора и особенно микрофлора толстого
кишечника оказывает существенное влияние на организм. Основные ее функции: защитная
(антагонизм к другим, в том числе патогенным микробам); иммуностимулирующая
(антигены микроорганизмов стимулируют развитие лимфоидной ткани); пищеварительная
(прежде всего обмен холестерина и желчных кислот); метаболическая
(синтез витаминов группы В- В1,2,6,12, К, никотиновой, пантотеновой, фолиевой
кислот). Существуют различные методы изучения роли нормальной
микрофлоры. Гнотобионты (безмикробные животные) используются для
изучения роли микроорганизмов для функционирования физиологических систем.
Гнотобиологические технологии используют для лечения иммунодефицитов, ожогов. В
результате разнообразных воздействий, снижающих естественную резистентность,
при тяжелых инфекционных и соматических заболеваниях и особенно при
нерациональном применении антибиотиков возникают дисбактериозы. Дисбактериоз-
изменения количественного и качественного состава микрофлоры, главным образом
кишечника. Чаще сопровождаются увеличением факультативно- анаэробной или
остаточной микрофлоры (грамотрицательных палочек - кишечной палочки, протея,
псевдомонад), стафилококков, грибов рода Candida. Эти микроорганизмы как
правило устойчивы к антибиотикам и при подавлении нормофлоры антибиотиками и
снижении естественной резистентности получают возможность беспрепятственно
размножаться. Наиболее тяжелые формы дисбактериозов- стафилококковые пневмонии,
колиты и сепсис, кандидомикозы, псевдомембранозный колит, вызываемый
Clostridium difficile. Для лечения используют биопрепараты, восстанавливающие
нормальную микрофлору- эубиотики- колибактерин (используют специальный штамм
E.coli, антогонист шигелл), лактобактерин, бифидумбактерин, бификол,
бактисубтил и другие, а также специальные бактериофаги.
Лекция № 9. Учение об инфекции. Инфекция” (лат. inficio- заражать) впервые было введено для обозначения венерических болезней. Инфекция- совокупность всех биологических явлений и процессов, возникающих в организме при внедрении и размножении в нем микроорганизмов, результат взаимоотношений между макро- и микроорганизмом в виде адаптационных и патологических процессов в организме т.е. инфекционного процесса. Инфекционная болезнь-выраженная форма инфекционного процесса. В общебиологическом плане взаимоотношения микро- и макроорганизмов представляют собой симбиоз (т.е. сожительство), так как все живые существа сосуществуют в природе. Человек сосуществует на планете Земля с микроорганизмами, растениями, животными. Основными формами взаимодействия микро- и макроорганизмов (их симбиоза) являются: мутуализм, комменсализм, паразитизм. Мутуализм- взаимовыгодные отношения (пример- нормальная микрофлора). Комменсализм- выгоду извлекает один партнер (микроб), не причиняя особого вреда другому. Необходимо отметить, что при любом типе взаимоотношений микроорганизм может проявить свои патогенные свойства (пример- условно- патогенные микробы- комменсалы в иммунодефицитном хозяине). Паразитизм- крайняя форма антогонистического симбиоза, когда микроорганизм питается за счет хозяина, т.е. извлекает выгоду, нанося при этом вред хозяину. Микробный паразитизм носит эволюционный характер. В процессе перехода от свободноживущего к паразитическому типу жизнедеятельности микроорганизмы теряют ряд ферментных систем, необходимых для существования во внешней среде, но приобретают ряд свойств, обеспечивающих возможность паразитизма.
Основные этапы инфекционного процесса.
1.Адгезия- прикрепление микроорганизма к соответствующим клеткам хозяина.
2.Колонизация- закрепление микроорганизмов в соответствующем участке.
3.Размножение (увеличение количества- мультипликация).
4.Пенетрация- проникновение в нижележащие слои и распространение инфекта.
5.Повреждение клеток и тканей (размножение, распространение инфекта). Инфекционный процесс может быть: по длительности- острый и хронический. Острая циклическая инфекция заканчивается элиминацией (удалением) возбудителя или смертью больного. При хронической инфекции возбудитель длительно сохраняется в организме (это состояние называется персистенция). Для персистенции микроорганизмы имеют ряд механизмов- внутриклеточная локализация (укрываются в клетке), переход в не имеющие клеточной стенки L- формы, антигенная мимикрия (совпадение по химическому составу антигенных детерминант микроба и клеток хозяина), укрытие в локальных очагах и забарьерных органах (головной мозг). Для вирусов дополнительными факторами персистенции является интеграция генома вируса с хромосомой клетки- мишени, недоступность действию антител, наличие дефектных вирусных частиц и слабая индукция иммунного ответа и др. Персистенция в организме и периодическая смена хозяина- два основных механизма поддержания микробных популяций.
по степени распространения- локальный и генерализованный. Локальный инфекционный процесс- возбудитель сосредоточен в определенном очаге, не выходя за его пределы, что сдерживает механизмы защиты. Если микроорганизм способен диссеминировать по организму, возникает генерализованный процесс. Существует два основных пути распространения-лимфогенный (по лимфатической сист.) и гематогенный (по кровяным сосудам).
по выраженности- манифестный и инаппарантный. Манифестный (ярко выраженный) инфекционный процесс- инфекционная болезнь- типичная, атипичная, хроническая и т.д. Бессимптомный (инаппарантный) инфекционный процесс характерен для латентной инфекции. Размножение возбудителя в организме не сопровождается клиническими проявлениями, а только иммунными реакциями. Инфекционные заболевания имеют ряд отличий от соматических, в том числе- наличие возбудителя, заразность, цикличность течения.
Динамика развития инфекционной болезни. Инфекционные заболевания характеризуются цикличностью, сменой периодов.
1.Инкубационный период- от момента заражения до первых клинических признаков (процесс активного размножения возбудителя).
2.Продромальный период (предвестников) характеризуется общими неспецифическими проявлениями- недомоганием, головной болью, повышением температуры и другими симптомами преимущественно токсического генеза.
3.Период развития (разгара) болезни характеризуется типичными (специфическими) для данной инфекции клиническими проявлениями.
4.Период реконвалесценции (выздоровления). В качестве исхода болезни может наступить выздоровление, развиться носительство или летальный исход.
Бактерионосительство может иметь большое значение в распространении многих инфекций. Может наблюдаться как при латентной инфекции, так и после перенесенного инфекционного заболевания. Особое значение при некоторых инфекциях имеют хронические носители (брюшной тиф, вирусный гепатит В). Инфекционное заболевание возникает не при каждом попадании патогенного микроорганизма в организм человека. Требуются определенные условия для реализации: достаточная доза микроорганизмов (понятие о критических дозах). Чума- несколько бактериальных клеток, дизентерия- десятки, для некоторых возбудителей- тысячи- сотни тысяч; естественный путь проникновения. Существует понятие о входных воротах инфекции, различных для различных групп инфекций- раневых, респираторных, кишечных, урогенитальных с различными механизмами заражения (глаза, кожа, дыхательные пути, ЖКТ, мочеполовая система и др.); характеристики возбудителя, его болезнетворные свойства, способность преодолевать защитные механизмы хозяина; состояние организма хозяина (наследственность- гетерогенность человеческой популяции по восприимчивости к инфекции, пол, возраст, состояние иммунной, нервной и эндокринной систем, образ жизни, природные и социальные условия жизни человека и др.).
Патогенность (“рождающий болезнь”)- способность микроорганизма вызвать заболевание. Это свойство характеризует видовые генетические особенности микроорганизмов, их генетически детерминированные характеристики, позволяющие преодолеть защитные механизмы хозяина, проявить свои патогенные свойства.
Вирулентность - фенотипическое (индивидуальное) количественное выражение патогенности (патогенного генотипа). Вирулентность может варьировать и может быть определена лабораторными методами (чаще- DL50- 50% летальная доза- количество патогенных микроорганизмов, позволяющая вызвать гибель 50% зараженных животных). По способности вызывать заболевания микроорганизмы можно разделить на патогенные, условно- патогенные, непатогенные. Условно-патогенные микроорганизмы обнаруживают как в окружающей среде, так и в составе нормальной микрофлоры. В определенных условиях (иммунодеф. состояния, травмы и операции с проникновением микроорганизмов в ткани) они могут вызывать эндогенные инфекции.
Основные факторы патогенности микроорганизмов - адгезины, ферменты патогенности, подавляющие фагоцитоз вещества, микробные токсины, в определенных условиях- капсула, подвижность микробов. Вирулентность связана с токсигенностью (способностью образования токсинов) и инвазивностью (способностью проникать в ткани хозяина, размножаться и распространяться). Токсигенность и инвазивность имеют самостоятельный генетический контроль, часто находятся в обратной зависимости (возбудитель с высокой токсигенностью может обладать низкой инвазивностью и наоборот). Патогенность - т.е. способность микроорганизма вызывать заболевание- более широкое понятие, чем паразитизм. Патогенными свойствами могут обладать не только паразитические виды микробов, но и свободно живущие, в т.ч. возбудители сапронозов (иерсинии, легионеллы и др.). Естественной средой для последних является почва и растительные организмы, однако они способны перестраивать свой метаболизм в организме теплокровных животных и оказывать патогенное действие. Адгезины и факторы колонизации- чаще поверхностные структуры бактериальной клетки, с помощью которых бактерии распознают рецепторы на мембранах клеток, прикрепляются к ним и колонизируют ткани. Функцию адгезии выполняют пили, белки наружной мембраны, ЛПС, тейхоевые кислоты, гемагглютинины вирусов. Адгезия- пусковой механизм реализации патогенных свойств возбудителей. Факторы инвазии, проникновения в клетки и ткани хозяина. Микроорганизмы могут размножаться вне клеток, на мембранах клеток, внутри клеток. Бактерии выделяют вещества, способствующие преодолению барьеров хозяина, их проникновению и размножению. У грамотрицательных бактерий это обычно белки наружной мембраны. К этим же факторам относятся ферменты патогенности. Ферменты патогенности- это факторы агрессии и защиты микроорганизмов. Способность к образованию экзоферментов во многом определяет инвазивность бактерий- возможность проникать через слизистые, соединительнотканные и другие барьеры. К ним относятся различные литические ферменты- гиалуронидаза, коллагеназа, лецитиназа, нейраминидаза, коагулаза, протеазы. Более подробно их характеристика дана в лекции по физиологии микроорганизмов. Важнейшими факторами патогенности считают токсины, которые можно разделить на две большие группы- экзотоксины и эндотоксины.
Экзотоксины продуцируются во внешнюю среду (организм хозяина), обычно белковой природы, могут проявлять ферментативную активность, могут секретировать как грамположительными, так и грамотрицательными бактериями. Они обладают очень высокой токсичностью, термически нестойки, часто проявляют антиметаболитные свойства. Экзотоксины проявляют высокую иммуногенность и вызывают образование специфических нейтрализующих антител- антитоксинов. По механизму действия и точке приложения экзотоксины отличаются- цитотоксины (энтеротоксины и дерматонекротоксины), мембранотоксины (гемолизины, лейкоцидины), функциональные блокаторы (холероген), эксфолианты и эритрогенины. Микробы, способные продуцировать экзотоксины, называют токсигенными.
Эндотоксины высвобождаются
только при гибели бактерий, характерны для грамотрицательных бактерий,
представляют собой сложные химические соединения клеточной стенки (ЛПС)-
подробнее смотри лекцию по химическому составу бактерий. Токсичность
определяется липидом А, токсин относительно термостоек; иммуногенные и
токсические свойства выражены более слабо, чем у экзотоксинов. Наличие капсул у
бактерий затрудняет начальные этапы защитных реакций- распознавание и
поглощение (фагоцитоз). Существенным фактором инвазивности является подвижность
бактерий, обусловливающая проникновение микробов в клетки и в межклеточные
пространства. Факторы патогенности контролируются: генами хромосомы; генами
плазмид; генами, привнесенными умеренными фагами.
Лекция № 10. Иммунитет, виды и формы. Структура иммунной системы.
Факторы неспецифической защиты.
Первоначально иммунология возникла как наука о невосприимчивости (иммунитете) к инфекционным болезням. Наиболее существенный вклад в ее создание внесли И.И.Мечников (фагоцитарная или клеточная теория иммунитета) и П.Эрлих (гуморальная теория), в творческой дискуссии между которыми совершенствовались представления об иммунитете.
В настоящее время считается, что наследственный (врожденный, видовой) и приобретенный иммунитет зависит от согласованной деятельности пяти основных систем: макрофагов, комплемента, интерферонов, Т- и В- лимфоцитов, главной системы гистосовместимости (МНС- в английском варианте), обеспечивающих различные формы иммунного ответа.
В современном понимании иммунология- это не только наука, изучающая защиту от инфекционных заболеваний. Иммунология- наука, изучающая механизмы самозащиты организма от всего генетически чужеродного, поддержания структурной и функциональной целостности организма (гомеостаза организма). Подробнее - см. лекцию 1.
Центральным биологическим механизмом иммунитета является механизм распознавания “своего” и “чужого”. Пример- необходимость защиты от собственных мутантных и раковых клеток (одномоментно в организме находится около 10 млн. измененных клеток).
Иммунитет- целостная система биологических механизмов самозащиты организма, с помощью которых он распознает и уничтожает все чужеродное (генетически отличающееся).
Выделяют две основные формы иммунитета- видовой (врожденный) и приобретенный. Приобретенный иммунитет может быть естественный (результат встречи с возбудителем) и искусственный (иммунизация), активный (вырабатываемый) и пассивный (получаемый), стерильный (без наличия возбудителя) и нестерильный (существующий в присутствии возбудителя в организме), гуморальный и клеточный, системный и местный, по направленности- антибактериальный, антивирусный, антитоксический, противоопухолевый, антитрансплантационный.
В основе видового иммунитета лежат различные механизмы естественной неспецифической резистентности. Среди них- кожные покровы и слизистые оболочки, нормальная микрофлора организма, фагоцитоз, воспаление, лихорадка, система комплемента, барьерные механизмы лимфоузлов, противомикробные вещества, выделительные системы организма, главная система гистосовместимости.
Кожа и слизистые- первая линия защиты против возбудителей. Кроме функции механического (анатомического) барьера кожа обладает бактерицидной активностью. Слизь, лизоцим, желудочный сок, слезная жидкость, слюна, деятельность мерцательного эпителия способствует защите слизистых оболочек.
Нормальная микрофлора организма препятствует колонизации организма посторонней микрофлорой (конкуренция за субстраты, различные формы антагонизма, в т.ч. выделение антибиотических веществ, изменение рН и др.).
Фагоцитоз и система комплемента- вторая линия защиты организма против микроорганизмов, преодолевших поверхностные барьеры. Клеточные факторы системы видовой резистентности- фагоциты, поглощающие и разрушающие патогенные микроорганизмы и другой генетически чужеродный материал. Представлены полиморфоядерными лейкоцитами или гранулоцитами- нейтрофилами, эозинофилами и базофилами (клетками миелопоэтического ряда), а также моноцитами и тканевыми макрофагами (клетками макрофагально- моноцитарной системы).
Значение фагоцитирующих клеток для защиты организма впервые доказал И.И.Мечников, разработавший фагоцитарную теорию иммунитета.
Стадии фагоцитоза.
Процесс фагоцитоза (поглощения твердофазного объекта) состоит из пяти стадий.
1.Активация (усиление энергетического метаболизма). Факторами активации и хемотаксиса являются бактериальные продукды (ЛПС, пептиды), компоненты комплемента (С3 и С5), цитокины и антитела.
2.Хемотаксис.
3.Адгезия.
4.Поглощение.
5.Исход фагоцитоза.
Адгезия связана с наличием ряда рецепторов на поверхности фагоцитов ( к Fc- фрагментам антител, компонентам комплемента, фибронектину), обеспечивающих прочность рецептор- опосредованных взаимодействий опсонинов, обволакивающих микроорганизмы и ограничивающих их подвижность (антитела, С3в, фибронектин).
Фагоциты обладают амебоподобными псевдоподиями. При поглощении образуется фагосома с поглощенным объектом (бактерией), к ней присоединяется и сливается содержащая литические ферменты лизосома, образуется фаголизосома.
Возможно три исхода фагоцитоза:
- завершенный фагоцитоз; - незавершенный фагоцитоз; - процессинг антигенов.
Завершенный фагоцитоз- полное переваривание микроорганизмов в клетке- фагоците.
Незавершенный фагоцитоз- выживание и даже размножение микроорганизмов в фагоците. Это характерно для факультативных и особенно - облигатных внутриклеточных паразитов. Механизмы персистирования в фагоцитах связаны с блокадой фагосомо- лизосомального слияния (вирус гриппа, микобактерии, токсоплазмы), резистентностью к действию лизосомальных ферментов (гонококки, стафилококки), способностью микробов быстро покидать фагосомы после поглощения и длительно пребывать в цитоплазме (риккетсии).
В процессе фагоцитоза происходит “окислительный взрыв” с образованием активных форм кислорода, что обеспечивает бактерицидный эффект.
К одной из важнейших функций макрофагов (наряду с хемотаксисом, фагоцитозом, секрецией биологически активных веществ) является переработка (процессинг) антигена и представление его иммунокомпетентным клеткам с участием белков главной системы гистосовместимости (МНС) класса 2.
Фагоцитоз- не только уничтожение чужеродного, но и представление антигена для запуска иммунных реакций и секреции медиаторов иммунных и воспалительных реакций. Система макрофагов- центральное звено не только естественной резистентности (видового иммунитета), но и играет важную роль в приобретенном иммунитете, кооперации клеток в иммунном ответе.
Воспаление как защитная реакция организма на различные повреждения тканей возникло на более высокой ступени эволюции, чем фагоцитоз и характерно для высокоорганизованных организмов, обладающих кровеносной и нервной системами.
Инфекционное воспаление сопровождается различными сосудистыми и клеточными (включая фагоцитоз) реакциями, а также запуском целого ряда медиаторов воспалительных реакций (гистамина, серотонина, кининов, белков острой фазы воспалеия, лейкотриенов и простагландинов, цитокинов, системы комплемента).
Многие бактериальные продукты активируют клетки макрофагально- моноцитарной системы и лимфоциты, отвечающие на них выделением биологически активных продуктов- цитокинов, в частности интерлейкинов. Их можно характеризовать как медиаторы клеточных иммунных реакций. В воспалительных реакциях основную роль имеет интерлейкин-1 (ИЛ-1), стимулирующий лихорадку, повышающий проницаемость сосудов и адгезивные свойства эндотелия, активирующий фагоциты.
Лихорадка. Повышение температуры тела- защитная реакция организма, ухудшающая условия для размножения многих микроорганизмов, активирует макрофаги, ускоряет кровоток и усиливает обменные процессы в организме.
Барьерные функции лимфоузлов. По выражению П.Ф.Здродовского (1969) лимфоузлы- своеобразный биологический фильтр для возбудителей, переносимых с лимфой. Здесь проникшие через кожу или слизистые и занесенные током лимфы микроорганизмы задерживаются и подвергаются действию макрофагов и активированных лимфоцитов.
Система комплемента- комплекс белков и гликопротеидов сыворотки крови человека и позвоночных животных (их более 20). Отдельные компоненты опосредуют процессы воспаления, опсонизацию чужеродных фрагментов для последующего фагоцитоза, участвуют наряду с макрофагами в непосредственном уничтожении микроорганизмов и других чужеродных клеток (лизис бактерий и вирусов). В условиях физиологической нормы компоненты системы комплемента находятся в неактивной форме. Известны три пути активации системы комплемента- классический, альтернативный и с использованием С1- шунта.
Классический путь- каскад протеазных реакций с компонента С1q до С9, реализуется при наличии антител к соответствующему антигену. С комплексом “антиген- антитела” взаимодействует компонент С1q, затем С4, следом- С2. Образуется комплекс “антиген- антитела-С1С4С2”, с ним соединяется С3 (центральный компонент системы) и запускается цепь активации с эффекторными функциями (опсонизация и лизис бактерий, активация системы макрофагов, воспаление).
Альтернативный путь реализуется при первичном контакте с возбудителем (когда еще нет антител). Он индуцируется ЛПС и другими микробными антигенами. С1, С4, С2 не участвуют, альтернативный и классический пути смыкаются на уровне С3.
Система интерферонов.
Интерфероны- синтезируемые различными клетками организма гликопротеиды широкого спектра биологической активности (прежде всего антивирусной), быстрый ответ организма на получение клетками неспецифического сигнала чужеродности. Существует целая система интерферонов, которые разделены на альфа, бета и гамма подтипы с выраженной гетерогенностью свойств. Противовирусное действие проявляется в способности подавлять внутриклеточное размножение ДНК- и РНК- вирусов (прежде всего в результате блокировки синтеза вирусных макромолекул). Индукцию синтеза интерферонов вызывают вирусы, бактерии, риккетсии, простейшие, синтетические соединения.
Киллерные клетки.
В обеспечении видового иммунитета существенную роль принадлежит Т- цитотоксическим лимфоцитам (Т- киллерам), а также главной системе гистосовместимости (подробнее- в следующих лекциях).
Т- киллеры по представлению антигенов главной системы гистосовместимости класса 1 распознают любые чужеродные антигены (включая мутантные, например- раковые клетки), атакуют и уничтожают их.
Клетки NK (natural killer- натуральные киллеры) имеют важное значение в поддержании генетического гомеостаза и противоопухолевой защите, их функции распознавания не зависят от представления антигенов МНС (major histocompatibility complex) класса 1.
Системы неспецифической резистентности и видового иммунитета способствуют поддержанию структурной и функциональной целостности организма и являются основой для формирования приобретенного (специфического) иммунитета. Стыкуясь на этом, более высоком уровне, системы видового и приобретенного иммунитета образуют единую и наиболее эффективную систему самозащиты организма от всего чужеродного.
Иммунная система
Иммунная система- совокупность органов, тканей и клеток, обеспечивающих клеточно- генетическое постоянство организма. Принципы антигенной (генетической) чистоты основываются на распознавании “своего- чужого” и в значительной степени обусловлены системой генов и гликопротеидов (продуктов их экспрессии)- главным комплексом гистосовместимости (MHC), у человека часто называемой системой HLA (human leucocyte antigens). На лейкоцитах человека четко экспрессированы белки МНС, с помощью исследования лейкоцитов типируют антигены МНС.
Органы иммунной системы.
Выделяют центральные (костный мозг- кроветворный орган, вилочковая железа или тимус, лимфоидная ткань кишечника) и периферические (селезенка, лимфатические узлы, скопления лимфоидной ткани в собственном слое слизистых оболочек кишечного типа) органы иммунитета.
Клетки- предшественники
иммунокомпетентных клеток продуцируются костным мозгом. Некоторые потомки
стволовых клеток становятся лимфоцитами. Лимфоциты подразделяют на два класса-
Т и В. Предшественники Т- лимфоцитов мигрируют в тимус, где созревают в клетки,
способные участвовать в иммунном ответе. У человека В- лимфоциты созревают в
костном мозге. У птиц незрелые В- клетки мигрируют в сумку (бурсу) Фабрициуса,
где достигают зрелости. Зрелые В- и Т- лимфоциты заселяют периферические
лимфоузлы. Таким образом, центральные органы иммунной системы осуществляют
образование и созревание иммунокомпетентных клеток, периферические органы
обеспечивают адекватный иммунный ответ на антигенную стимуляцию- “обработку”
антигена, его распознавание и клональную пролиферацию лимфоцитов- антиген-
зависимую дифференцировку.
Лекция № 11. Антигены, основные свойства. Антигены гистосовместимости. Процессинг антигенов.
Антигены- вещества различного происхождения, несущие признаки генетической чужеродности и вызывающие развитие иммунных реакций (гуморальных, клеточных, иммунологической толерантности, иммунологической памяти и др.).
Свойства антигенов, наряду с чужеродностью, определяет их иммуногенность- способность вызывать иммунный ответ и антигенность- способность (антигена) избирательно взаимодействовать со специфическими антителами или антиген- распознающими рецепторами лимфоцитов.
Антигенами могут быть белки, полисахариды и нуклеиновые кислоты в комбинации между собой или липидами. Антигенами являются любые структуры, несущие признаки генетической чужеродности и распознаваемые в этом качестве иммунной системой. Наибольшей иммуногенностью обладают белковые антигены, в том числе бактериальные экзотоксины, вирусная нейраминидаза.
Многообразие понятия “антиген”.
Антигены разделены на полные (иммуногенные), всегда проявляющие иммуногенные и антигенные свойства, и неполные (гаптены), не способные самостоятельно вызывать иммунный ответ.
Гаптены обладают антигенностью, что обусловливает их специфичность, способность избирательно взаимодействовать с антителами или рецепторами лимфоцитов, определяться иммунологическими реакциями. Гаптены могут стать иммуногенными при связывании с иммуногенным носителем (например, белком), т.е. становятся полными.
За специфичность антигена отвечает гаптенная часть, за иммуногенность- носитель (чаще белок).
Иммуногенность зависит от ряда причин (молекулярного веса, подвижности молекул антигена, формы, структуры, способности к изменению). Существенное значение имеет степень гетерогенности антигена, т.е. чужеродность для данного вида (макроорганизма), степени эволюционной дивергенции молекул, уникальности и необычности структуры. Чужеродность определяется также молекулярной массой, размерами и строением биополимера, его макромолекулярностью и жесткостью структуры. Белки и другие высокомолекулярные вещества с более высоким молекулярным весом наиболее иммуногенны. Большое значение имеет жесткость структуры, что связано с наличием ароматических колец в составе аминокислотных последовательностей. Последовательность аминокислот в полипептидных цепочках- генетически детерминированный признак.
Антигенность белков является проявлением их чужеродности, а ее специфичность зависит от аминокислотной последовательности белков, вторичной, третичной и четвертичной (т.е. от общей конформации белковой молекулы) структуры, от поверхностно расположенных детерминантных групп и концевых аминокислотных остатков. Коллоидное состояние и растворимость- обязательные свойства антигенов.
Специфичность антигенов зависит от особых участков молекул белков и полисахаридов, называемых эпитопами. Эпитопы или антигенные детерминанты- фрагменты молекул антигена, вызывающие иммунный ответ и определяющие его специфичность. Антигенные детерминанты избирательно реагируют с антителами или антиген- распознающими рецепторами клетки.
Структура многих антигенных детерминант известна. У белков это обычно фрагменты из 8- 20 выступающих на поверхности аминокислотных остатков, у полисахаридов- выступающие О- боковые дезоксисахаридные цепи в составе ЛПС, у вируса гриппа- гемагглютинин, у вируса иммунодефицита человека- мембранный гликопептид.
Эпитопы качественно могут отличаться, к каждому могут образовываться “свои” антитела. Антигены, содержащие одну антигенную детерминанту, называют моновалентными, ряд эпитопов- поливалентными. Полимерные антигены содержат в большом количестве идентичные эпитопы (флагеллины, ЛПС).
Основные типы антигенной специфичности (зависят от специфичности эпитопов).
1.Видовая- характерна для всех особей одного вида (общие эпитопы).
2.Групповая- внутри вида (изоантигены, которые характерны для отдельных групп). Пример- группы крови (АВО и др.).
3.Гетероспецифичность- наличие общих антигенных детерминант у организмов различных таксономических групп. Имеются перекрестно- реагирующие антигены у бактерий и тканей макроорганизма.
а. Антиген Форсмана- типичный перекрестно- реагирующий антиген, выявлен в
эритроцитах кошек, собак, овец, почке морской свинки.
б.Rh- система эритроцитов. У человека Rh- антигены агглютинируют антитела
к эритроцитам обезьян Macacus rhesus, т.е. являются перекрестными.
в. Известны общие антигенные детерминанты эритроцитов человека и палочки
чумы, вирусов оспы и гриппа.
г. Еще пример- белок А стрептококка и ткани миокарда (клапанный аппарат).
Подобная антигенная мимикрия обманывает иммунную систему, защищает от ее воздействия микроорганизмы. Наличие перекрестных антигенов способно блокировать системы, распознающие чужеродные структуры.
4.Патологическая. При различных патологических изменениях тканей происходят изменения химических соединений, что может изменять нормальную антигенную специфичность. Появляются “ожоговые”, “лучевые”, “раковые” антигены с измененной видовой специфичностью. Существует понятие аутоантигенов - веществ организма, к которым могут возникать иммунные реакции ( так называемые аутоиммунные реакции), направленные против определенных тканей организма. Чаще всего это относится к органам и тканям, в норме не подвергающихся воздействию иммунной системы в связи с наличием барьеров (мозг, хрусталик, паращитовидные железы и др.).
5.Стадиоспецифичность. Имеются антигены, характерные для определенных стадий развития, связанные с морфогенезом. Альфа- фетопротеин характерен для эмбрионального развития, синтез во взрослом состоянии резко увеличивается при раковых заболеваниях печени.
Антигенная специфичность и антигенное строение бактерий.
Для характеристики микроорганизмов выделяют родовую, видовую, групповую и типовую специфичность антигенов. Наиболее точная дифференциация осуществляется с использованием моноклональных антител (МКА), распознающих только одну антигенную детерминанту.
Обладая сложным химическим строением, бактериальная клетка представляет целый комплекс антигенов. Антигенными свойствами обладают жгутики, капсула, клеточная стенка, цитоплазматическая мембрана, рибосомы и другие компоненты цитоплазмы, токсины, ферменты.
Основными видами бактериальных антигенов являются:
- соматические или О- антигены (у грамотрицательных бактерий специфичность определяется дезоксисахарами полисахаридов ЛПС); - жгутиковые или Н- антигены (белковые); - поверхностные или капсульные К- антигены.
Выделяют протективные антигены, обеспечивающие защиту (протекцию) против соответствующих инфекций, что используется для создания вакцин.
Суперантигены (некоторые экзотоксины, например- стафилококковый) вызывают чрезмерно сильную иммунную реакцию, часто приводят к побочным реакциям, развитию иммунодефицита или аутоиммунных реакций.
Антигены гистосовместимости.
При пересадках органов возникает проблема совместимости тканей, связанная со степенью их генетического родства, реакциями отторжения чужеродных аллогенных и ксеногенных трансплантатов, т.е. проблемами трансплантационного иммунитета. Существует ряд тканевых антигенов. Трансплантационные антигены во многом определяют индивидуальную антигенную специфичность организма. Сопокупность генов, определяющих синтез трансплантационных антигенов, получила название главной системы гистосовместимости. У людей она часто называется системой HLA (Human leucocyte antigens), в связи с четким представительством на лейкоцитах трансплантационных антигенов. Гены этой системы расположены на коротком плече хромосомы С6. Система HLA- это система сильных антигенов. Спектр молекул МНС уникален для организма, что определяет его биологическую индивидуальность и позволяет различать “чужое- несовместимое”.
Семь генетических локусов системы разделены на три класса.
Гены первого класса контролизуют синтез антигенов класса 1, определяют тканевые антигены и контролируют гистосовместимость. Антигены класса 1 определяют индивидуальную антигенную специфичность, они представляют любые чужеродные антигены Т- цитотоксическим лимфоцитам. Антигены класса 1 представлены на поверхности всех ядросодержащих клеток. Молекулы МНС класса 1 взаимодействуют с молекулой CD8, экспрессируемой на мембране предшественников цитотоксических лимфоцитов (CD- claster difference).
Гены МНС класса 2 контролируют антигены класса 2. Они контролируют ответ к тимусзависимым антигенам. Антигены класса 2 экспрессированы преимущественно на мембране иммунокомпетентных клеток (прежде всего макрофагов и В- лимфоцитов, частично- активированных Т- лимфоцитов). К этой же группе генов (точнее- области HLA- D) относятся также гены Ir - силы иммунного ответа и гены Is - супрессии иммунного ответа. Антигены МНС класса 2 обеспечивают взаимодействие между макрофагами и В- лимфоцитами, участвуют во всех стадиях иммунного ответа- представлении антигена макрофагами Т- лимфоцитам, взаимодействии (кооперации) макрофагов, Т- и В- лимфоцитов, дифференцировке иммунокомпетентных клеток. Антигены класса 2 принимают участие в формировании противомикробного, противоопухолевого, трансплантационного и других видов иммунитета.
Структуры, с помощью которых белки МНС классов 1 и 2 связывают антигены (так называемые активные центры) по уровню специфичности уступают только активным центрам антител.
Гены МНС класса 3 кодируют отдельные компоненты системы комплемента.
Процессинг антигенов- это их судьба в организме. Одной из важнейших функций макрофагов является переработка антигена в иммуногенную форму (это собственно и есть процессинг антигена) и представление его иммунокомпетентным клеткам. В процессинге, наряду с макрофагами, участвуют В- лимфоциты, дендритные клетки, Т- лимфоциты. Под процессингом понимают такую переработку антигена, в результате которой пептидные фрагменты антигена (эпитопы), необходимые для передачи (представления), отбираются и связываются с белками МНС класса 2 (или класса 1). В таком комплексном виде антигенная информация передается лимфоцитам. Дендритные клетки имеют значение в фиксации и длительном хранении (депонировании) переработанного антигена.
Экзогенные антигены подвергаются эндоцитозу и расщеплению в антиген- представляющих (презентирующих) клетках. Фрагмент антигена, содержащий антигенную детерминанту, в комплексе с молекулой класса 2 МНС транспортируется к плазматической мембране антиген- представляющей клетки, встраивается в нее и представляется CD4 Т- лимфоцитам.
Эндогенные антигены - продукты собственных клеток организма. Это могут быть вирусные белки или аномальные белки опухолевых клеток. Их антигенные детерминанты представляются CD8 Т- лимфоцитам в комплексе с молекулой класса 1 МНС.
Лекция № 12. Гуморальный иммунитет. Иммуноглобулины. Роль антител в иммунном ответе. Реакция антиген- антитело, ее применение.
Основными формами иммунного ответа на попадание антигена в организм являются: биосинтез антител, образование клеток иммунной памяти, реакция гиперчувствительности немедленного типа, реакция гиперчувствительности замедленного типа, иммунологическая толерантность, идиотип- антиидиотипические отношения.
Для гуморального иммунитета характерна выработка специфических антител (иммуноглобулинов).
Антитела - специфические белки гамма- глобулиновой природы, образующиеся в организме в ответ на антигенную стимуляцию и способные специфически взаимодействовать с антигеном (in vivo, in vitro). В соответствии с международной классификацией совокупность сывороточных белков, обладающих свойствами антител, называют иммуноглобулинами.
Уникальность антител заключается в том, что они способны специфически взаимодействовать только с тем антигеном, который вызвал их образование.
Иммуноглобулины ( Ig ) разделены в зависимости от локализации на три группы:
- сывороточные (в крови);
- секреторные ( в секретах- содержимом желудочно- кишечного тракта, слезном секрете, слюне, особенно- в грудном молоке) обеспечивают местный иммунитет (иммунитет слизистых);
- поверхностные ( на поверхности иммунокомпетентных клеток, особенно В- лимфоцитов).
Любая молекула антител имеет сходное строение ( Y- образную форму) и состоит из двух тяжелых ( Н ) и двух легких ( L ) цепей, связанных дисульфидными мостиками. Каждая молекула антител имеет два одинаковых антигенсвязывающих фрагмента Fab ( fragment antigen binding ), определяющих антительную специфичность, и один Fc ( fragment constant ) фрагмент, который не связывает антиген, но обладает эффекторными биологическими функциями. Он взаимодействует со “своим” рецептором в мембране различных типов клеток ( макрофаг, тучная клетка, нейтрофил).
Концевые участки легких и тяжелых цепей молекулы иммуноглобулина вариабельны по составу ( аминокислотным последовательностям ) и обозначаются как VL и VH области. В их составе выделяют гипервариабельные участки, которые определяют структуру активного центра антител (антигенсвязывающий центр или паратоп). Именно с ним взаимодействует антигенная детерминанта (эпитоп) антигена. Антигенсвязывающий центр антител комплементарен эпитопу антигена по принципу “ключ - замок” и образован гипервариабельными областями L- и Н- цепей. Антитело свяжется антигеном (ключ попадет в замок) только в том случае, если детерминантная группа антигена полностью вместится в щель активного центра антител.
Легкие и тяжелые цепи состоят из отдельных блоков- доменов. В легких ( L ) цепях - два домена- один вариабельный ( V ) и один константный ( C ), в тяжелых ( H ) цепях- один V и 3 или 4 ( в зависимости от класса иммуноглобулина ) C домена.
Существуют легкие цепи двух типов- каппа и лямбда, они встречаются в различных пропорциях в составе различных (всех) классов иммуноглобулинов.
Выявлено пять классов тяжелых цепей- альфа ( с двумя подклассами), гамма ( с четырьмя подклассами), эксилон, мю и дельта. Соответственно обозначению тяжелой цепи обозначается и класс молекул иммуноглобулинов- А, G, E, M и D.
Именно константные области тяжелых цепей, различаясь по аминокислотному составу у различных классов иммуноглобулинов, в конечном результате и определяют специфические свойства иммуноглобулинов каждого класса.
Известно пять классов иммуноглобулинов, отличающихся по строению тяжелых цепей, молекулярной массе, физико- химическим и биологическим характеристикам: IgG, IgM, IgA, IgE, IgD. В составе IgG выделяют 4 подкласса ( IgG1, IgG2, IgG3, IgG4 ), в составе IgA- два подкласса (IgA1, IgA2 ).
Структурной единицей антител является мономер, состоящий из двух легких и двух тяжелых цепей. Мономерами являются IgG, IgA ( сывороточный), IgD и IgE. IgM- пентамер (полимерный Ig). У полимерных иммуноглобулинов имеется дополнительная j ( joint) полипептидная цепь, которая объединяет ( полимеризует) отдельные субъединицы (в составе пентамера IgM, ди- и тримера секреторного IgA).
Основные биологические характеристики антител.
1. Специфичность - способность взаимодействия с определенным (своим) антигеном (соответствие эпитопа антигена и активного центра антител).
2. Валентность- количество способных реагировать с антигеном активных центров ( это связано с молекулярной организацией- моно- или полимер). Иммуноглобулины могут быть двухвалентными ( IgG ) или поливалентными (пентамер IgM имеет 10 активных центров). Двух- и более валентные антитела навывают полными антителами. Неполные антитела имеют только один участвующий во взаимодействии с антигеном активный центр ( блокирующий эффект на иммунологические реакции, например, на агглютинационные тесты). Их выявляют в антиглобулиновой пробе Кумбса, реакции угнетения связывания комплемента.
3. Афинность - прочность связи между эпитопом антигена и активным центром антител, зависит от их пространственного соответствия.
4. Авидность - интегральная характеристика силы связи между антигеном и антителами, с учетом взаимодействия всех активных центров антител с эпитопами. Поскольку антигены часто поливалентны, связь между отдельными молекулами антигена осуществляется с помощью нескольких антител.
5. Гетерогенность - обусловлена антигенными свойствами антител, наличием у них трех видов антигенных детерминант:
- изотипические - принадлежность антител к определенному классу иммуноглобулинов;
- аллотипические- обусловлены аллельными различиями иммуноглобулинов, кодируемых соответствующими аллелями Ig гена;
- идиотипические- отражают индивидуальные особенности иммуноглобулина, определяемые характеристиками активных центров молекул антител. Даже тогда, когда антитела к конкретному антигену относятся к одному классу, субклассу и даже аллотипу, они характеризуются специфическими отличиями друг от друга (идиотипом). Это зависит от особенностей строения V- участков H- и L- цепей, множества различных вариантов их аминокислотных последовательностей.
Понятие о поликлональных и моноклональных антителах будет дано в следующих разделах.
Характеристика основных классов иммуноглобулинов.
Ig G. Мономеры, включают четыре субкласса. Концентрация в крови- от 8 до 17 г/л, период полураспада- около 3- 4 недель. Это основной класс иммуноглобулинов, защищающих организм от бактерий, токсинов и вирусов. В наибольшем количестве IgG- антитела вырабатываются на стадии выздоровления после инфекционного заболевания (поздние или 7S антитела), при вторичном иммунном ответе. IgG1 и IgG4 специфически (через Fab- фрагменты) связывают возбудителей (опсонизация), благодаря Fc- фрагментам IgG взаимодействуют с Fc- рецепторам фагоцитов, способствуя фагоцитозу и лизису микроорганизмов. IgG способны нейтрализовать бактериальные экзотоксины, связывать комплемент. Только IgG способны транспортироваться через плаценту от матери к плоду (проходить через плацентарный барьер) и обеспечивать защиту материнскими антителами плода и новорожденного. В отличие от IgM- антител, IgG- антитела относятся к категории поздних- появляются позже и более длительно выявляются в крови.
IgM. Молекула этого иммуноглобулина представляет собой полимерный Ig из пяти субъединиц, соединенных дисульфидными связями и дополнительной J- цепью, имеет 10 антиген- связывающих центров. Филогенетически это наиболее древний иммуноглобулин. IgM- наиболее ранний класс антител, образующихся при первичном попадании антигена в организм. Наличие IgM- антител к соответствующему возбудителю свидетельствует о свежем инфицировании (текущем инфекционном процессе). Антитела к антигенам грамотрицательных бактерий, жгутиковым антигенам- преимущественно IgM- антитела. IgM- основной класс иммуноглобулинов, синтезируемых у новорожденных и младенцев. IgM у новорожденных- это показатель внутриутробного заражения (краснуха, ЦМВ, токсоплазмоз и другие внутриутробные инфекции), поскольку материнские IgM через плаценту не проходят. Концентрация IgM в крови ниже, чем IgG- 0,5- 2,0 г/л, период полураспада- около недели. IgM способны агглютинировать бактерии, нейтрализовать вирусы, активировать комплемент, активизировать фагоцитоз, связывать эндотоксины грамотрицательных бактерий. IgM обладают большей, чем IgG авидностью (10 активных центров), аффинность (сродство к антигену) меньше, чем у IgG.
IgA. Выделяют сывороточные IgA (мономер) и секреторные IgA (IgAs). Сывороточные IgA составляют 1,4- 4,2 г/л. Секреторные IgAs находятся в слюне, пищеварительных соках, секрете слизистой носа, в молозиве. Они являются первой линией защиты слизистых, обеспечивая их местный иммунитет. IgAs состоят из Ig мономера, J-цепи и гликопротеина (секреторного компонента). Выделяют два изотипа- IgA1 преобладает в сыворотке, субкласс IgA2 - в экстраваскулярных секретах.
Секреторный компонент вырабатывается эпителиальными клетками слизистых оболочек и присоединяется к молекуле IgA в момент прохождения последней через эпителиальные клетки. Секреторный компонент повышает устойчивость молекул IgAs к действию протеолитических ферментов. Основная роль IgA- обеспечение местного иммунитета слизистых. Они препятствуют прикреплению бактерий к слизистым, обеспечивают транспорт полимерных иммунных комплексов с IgA, нейтрализуют энтеротоксин, активируют фагоцитоз и систему комплемента.
IgE. Представляет мономер, в сыворотке крови находится в низких концентра-
циях. Основная роль- своими Fc- фрагментами прикрепляется к тучным клеткам (мастоцитам) и базофилам и опосредует реакции гиперчувствительности немедленного типа. К IgE относятся “антитела аллергии”- реагины. Уровень IgE повышается при аллергических состояниях, гельминтозах. Антигенсвязывающие Fab- фрагменты молекулы IgE специфически взаимодействует с антигеном (аллергеном), сформировавшийся иммунный комплекс взаимодействует с рецепторами Fc- фрагментов IgE, встроенных в клеточную мембрану базофила или тучной клетки. Это является сигналом для выделения гистамина, других биологически активных веществ и развертывания острой аллергической реакции.
IgD. Мономеры IgD обнаруживают на поверхности развивающихся В- лимфоцитов, в сыворотке находятся в крайне низких концентрациях. Их биологическая роль точно не установлена. Полагают, что IgD участвуют в дифференциации Вклеток, способствуют развитию антиидиотипического ответа, участвуют в аутоиммунных процессах.
С целью определения концентраций иммуноглобулинов отдельных классов применяют несколько методов, чаще используют метод радиальной иммунодиффузии в геле (по Манчини)- разновидность реакции преципитации и ИФА.
Определение антител различных классов имеет важное значение для диагностики инфекционных заболеваний. Обнаружение антител к антигенам микроорганизмов в сыворотках крови- важный критерий при постановке диагноза- серологический метод диагностики. Антитела класса IgM появляются в остром периоде заболевания и относительно быстро исчезают, антитела класса IgG выявляются в более поздние сроки и более длительно (иногда- годами) сохраняются в сыворотках крови переболевших, их в этом случае называют анамнестическими антителами.
Выделяют понятия: титр антител, диагностический титр, исследования парных сывороток. Наибольшее значение имеет выявление IgM- антител и четырехкратное повышение титров антител (или сероконверсия- антитела выявляют во второй пробе при отрицательных результатах с первой сывороткой крови) при исследовании парных- взятых в динамике инфекционного процесса с интервалом в несколько дней- недель проб.
Реакции взаимодействия антител с возбудителями и их антигенами (реакция “антиген- антитело”) проявляется в виде ряда феноменов- агглютинации, преципитации, нейтрализации, лизиса, связывания комплемента, опсонизации, цитотоксичности и могут быть выявлены различными серологическими реакциями.
Динамика выработки антител. Первичный и вторичный иммунный ответ.
Первичный ответ- при первичном контакте с возбудителем (антигеном), вторичный- при повторном контакте. Основные отличия:
- продолжительность скрытого периода (больше- при первичном);
- скорость нарастания антител (быстрее- при вторичном);
- количество синтезируемых антител (больше- при повторном контакте);
- последовательность синтеза антител различных классов (при первичном более длительно преобладают IgM, при вторичном- быстро синтезируются и преобладают IgG- антитела).
Вторичный иммунный ответ обусловлен формированием клеток иммунной памяти. Пример вторичного иммунного ответа- встреча с возбудителем после вакцинации.
Роль антител в формировании иммунитета.
Антитела имеют важное значение в формировании приобретенного постинфекционного и поствакцинального иммунитета.
1. Связываясь с токсинами, антитела нейтрализуют их, обеспечивая антитоксический иммунитет.
2. Блокируя рецепторы вирусов, антитела препятствуют адсорбции вирусов на клетках, участвуют в противовирусном иммунитете.
3. Комплекс антиген- антитело запускает классический путь активации комплемента с его эффекторными функциями (лизис бактерий, опсонизация, воспаление, стимуляция макрофагов).
4. Антитела принимают участие в опсонизации бактерий, способствуя более эффективному фагоцитозу.
5. Антитела способствуют выведению из организма (с мочой, желчью) растворимых антигенов в виде циркулирующих иммунных комплексов.
IgG принадлежит наибольшая роль в
антитоксическом иммунитете, IgM- в антимикробном иммунитете (фагоцитоз
корпускулярных антигенов), особенно в отношении грамотрицательных бактерий,
IgA- в противовирусном иммунитете (нейтрализация вирусов), IgAs- в местном
иммунитете слизистых оболочек, IgE- в реакциях гиперчувствительности
немедленного типа.
Лекция № 13. Т- и В- лимфоциты. Рецепторы, субпопуляции. Кооперация клеток в иммунном ответе.
К клеткам иммунной системы относят лимфоциты, макрофаги и другие антиген- представляющие клетки (А- клетки, от англ. accessory- вспомогательный), а также так называемую третью популяцию клеток (т.е. клеток, не имеющих основных поверхностных маркеров Т- и В- лимфоцитов, А- клеток).
По функциональным свойствам все иммунокомпетентные клетки разделяют на эффекторные и регуляторные. Взаимодействие клеток в иммунном ответе осуществляется с помощью гуморальных медиаторов - цитокинов. Основные клетки иммунной системы- Т- и В- лимфоциты.
Лимфоциты.
В организме лимфоциты постоянно рециркулируют между зонами скопления лимфоидной ткани. Расположение лимфоцитов в лимфоидных органах и их миграция по кровеносному и лимфатическому руслу строго упорядочены и связаны с функциями различных субпопуляций.
Лимфоциты имеют общую морфологическую характеристику, однако их функции, поверхностные CD ( от claster differenciation) маркеры, индивидуальное (клональное) происхождение, различны.
По наличию поверхностных CD маркеров лимфоциты разделяют на функционально различные популяции и субпопуляции, прежде всего на Т- (тимусзависимые, прошедшие первичную дифференцировку в тимусе) лимфоциты и В - (bursa- зависимые, прошедшие созревание в сумке Фабрициуса у птиц или его аналогах у млекопитающих) лимфоциты.
Т- лимфоциты.
Локализация.
Обычно локализуются в так называемых Т- зависимых зонах периферических лимфоидных органов (периартикулярно в белой пульпе селезенки и паракортикальных зонах лимфоузлов).
Функции.
Т- лимфоциты распознают процессированный и представленный на поверхности антиген- представляющих ( А ) клеток антиген. Они отвечают за клеточный иммунитет, иммунные реакции клеточного типа. Отдельные субпопуляции помогают В- лимфоцитам реагировать на Т- зависимые антигены выработкой антител.
Происхождение и созревание.
Родоначальницей всех клеток крови, в том числе лимфоцитов, является единая стволовая клетка костного мозга. Она генерирует два типа клеток- предшественников- лимфоидную стволовую клетку и предшественника клеток красной крови, от которой происходят и клетки- предшественники лейкоцитов и макрофагов.
Образование и созревание иммунокомпетентных клеток осуществляется в центральных органах иммунитета (для Т- лимфоцитов- в тимусе). Клетки- предшественники Т- лимфоцитов попадают в тимус, где пре- Т- клетки (тимоциты) созревают, пролиферируют и проходят дифференцировку на отдельные субклассы в результате взаимодействия с эпителиальными и дендритными клетками стромы и воздействия гормоноподобных полипептидных факторов, секретируемых эпителиальными клетками тимуса ( альфа1- тимозин, тимопоэтин, тимулин и др.).
При дифференцировке Т- лимфоциты приобретают определенный набор мембранных CD- маркеров. Т-клетки разделяют на субпопуляции в соответствии с их функцией и профилем CD- маркеров.
Т- лимфоциты распознают антигены с помощью двух типов мембранных гликопротеинов- Т- клеточных рецепторов (семейство Ig- подобных молекул) и CD3, нековалентно связанных между собой. Их рецепторы, в отличие от антител и рецепторов В- лимфоцитов, не распознают свободно циркулирующие антигены. Они распознают пептидные фрагменты, представляемые им А- клетками через комплекс чужеродных веществ с соответствующим белком главной системы гистосовместимости 1 и 2 класса.
Выделяют три основные группы Т- лимфоцитов- помощники (активаторы), эффекторы, регуляторы.
Первая группа- помощники (активаторы), в состав которых входят Т- хелперы1, Т- хелперы2, индукторы Т- хелперов, индукторы Т- супрессоров.
1. Т- хелперы1 несут рецепторы CD4 (как и Т- хелперы2) и CD44, отвечают за созревание Т- цитотоксических лимфоцитов (Т- киллеров), активируют Т- хелперы2 и цитотоксическую функцию макрофагов, секретируют ИЛ-2, ИЛ-3 и другие цитокины.
2. Т- хелперы2 имеют общий для хелперов CD4 и специфический CD28 рецепторы, обеспечивают пролиферацию и дифференцировку В- лимфоцитов в антителпродуцирующие (плазматические) клетки, синтез антител, тормозят функцию Т- хелперов1, секретируют ИЛ-4, ИЛ-5 и ИЛ-6.
3. Индукторы Т- хелперов несут CD29, отвечают за экспрессию антигенов HLA класса 2 на макрофагах и других А- клетках.
4. Индукторы Т- супрессоров несут CD45 специфический рецептор, отвечают за секрецию ИЛ-1 макрофагами, активацию дифференцировки предшественников Т- супрессоров.
Вторая группа- Т- эффекторы. В нее входит только одна субпопуляция.
5. Т- цитотоксические лимфоциты (Т- киллеры). Имеют специфический рецептор CD8, лизируют клетки- мишени, несущие чужеродные антигены или измененные аутоантигены (трансплантант, опухоль, вирус и др.). ЦТЛ распознают чужеродный эпитоп вирусного или опухолевого антигена в комплексе с молекулой класса 1 HLA в плазматической мембране клетки- мишени.
Третья группа- Т-клетки- регуляторы. Представлена двумя основными субпопуляциями.
6. Т- супрессоры имеют важное значение в регуляции иммунитета, обеспечивая подавление функций Т- хелперов 1 и 2, В- лимфоцитов. Имеют рецепторы CD11, CD8. Группа функционально разнородна. Их активация происходит в результате непосредственной стимуляции антигеном без существенного участия главной системы гистосовместимости.
7. Т- контсупрессоры. Не имеют CD4, CD8, имеют рецептор к особому лейкину.
Способствуют подавлению функций Т- супрессоров, вырабатывают резистентность Т- хелперов к эффекту Т- супрессоров.
В- лимфоциты.
Существует несколько подтипов В- лимфоцитов. Основная функция В- клеток- эффекторное участие в гуморальных иммунных реакциях, дифференциация в результате антигенной стимуляции в плазматические клетки, продуцирующие антитела.
Образование В- клеток у плода происходит в печени, в дальнейшем- в костном мозге. Процесс созревания В- клеток осуществляется в две стадии- антиген - независимую и антиген - зависимую.
Антиген -независимая фаза. В- лимфоцит в процессе созревания проходит стадию пре- В- лимфоцита- активно пролиферирующей клетки, имеющей цитоплазменные H- цепи типа C мю (т.е. IgM). Следующая стадия- незрелый В- лимфоцит характеризуется появлением мембранного (рецепторного) IgM на поверхности. Конечная стадия антигеннезависимой дифференцировки- образование зрелого В- лимфоцита, который может иметь два мембранных рецептора с одинаковой антигенной специфичностью (изотипа) - IgM и IgD. Зрелые В- лимфоциты покидают костный мозг и заселяют селезенку, лимфоузлы и другие скопления лимфоидной ткани, где их развитие задерживается до встречи со “своим” антигеном, т.е. до осуществления антиген- зависимой дифференцировки.
Антиген- зависимая дифференцировка включает активацию, пролиферацию и дифференцировку В- клеток в плазматические клетки и В- клетки памяти. Активация осуществляется различными путями, что зависит от свойств антигенов и участия других клеток ( макрофагов, Т- хелперов). Большинство антигенов, индуцирующих синтез антител, для индукции иммунного ответа требуют участия Т- клеток- тимус- зависимые пнтигены. Тимус- независимые антигены (ЛПС, высокомолекулярные синтетические полимеры) способны стимулировать синтез антител без помощи Т- лимфоцитов.
В- лимфоцит с помощью своих иммуноглобулиновых рецепторов распознает и связывает антиген. Одновременно с В- клеткой антиген по представлению макрофага распознается Т- хелпером (Т- хелпером 2), который активируется и начинает синтезировать факторы роста и дифференцировки. Активированный этими факторами В- лимфоцит претерпевает ряд делений и одновременно дифференцируется в плазматические клетки, продуцирующие антитела.
Пути активации В- клеток и кооперации клеток в иммунном ответе на различные антигены и с участием популяций имеющих и не имеющих антиген Lyb5 популяций В- клеток отличаются. Активация В- лимфоцитов может осуществляться:
- Т- зависимым антигеном при участии белков МНС класса 2 Т- хелпера;
- Т- независимым антигеном, имеющим в составе митогенные компоненты;
- поликлональным активатором (ЛПС);
- анти- мю иммуноглобулинами;
- Т- независимым антигеном, не имеющим митогенного компонента.
Кооперация клеток в иммунном ответе.
В формировании иммунного ответа включаются все звенья иммунной системы- системы макрофагов, Т- и В- лимфоцитов, комплемента, интерферонов и главная система гистосовместимости.
В кратком виде можно выделить следующие этапы.
1. Поглощение и процессинг антигена макрофагом.
2. Представление процессированного антигена макрофагом с помощью белка главной системы гистосовместимости класса 2 Т- хелперам.
3. Узнавание антигена Т- хелперами и их активация.
4. Узнавание антигена и активация В- лимфоцитов.
5. Дифференциация В- лимфоцитов в плазматические клетки, синтез антител.
6. Взаимодействие антител с антигеном, активация систем комплемента и макрофагов, интерферонов.
7. Представление при участии белков МНС класса 1 чужеродных антигенов Т- киллерам, разрушение инфицированных чужеродными антигенами клеток Т- киллерами.
8. Индукция Т- и В- клеток иммунной памяти, способных специфически распознавать антиген и участвовать во вторичном иммунном ответе ( антигенстимулированные лимфоциты).
Клетки иммунной памяти. Поддержание долгоживущих и метаболически малоактивных клеток памяти, рециркулирующих в организме, является основой длительного сохранения приобретенного иммунитета. Состояние иммунной памяти обусловлено не только длительностью жизни Т- и В- клеток памяти, но и их антигенной стимуляцией. Длительное сохранение антигенов в организме обеспечивается дендритными клетками (депо антигенов), сохраняющими их на своей поверхности.
Дендритные клетки - популяции отросчатых клеток лимфоидной ткани костномозгового (моноцитарного) генеза, представляющая антигенные пептиды Т- лимфоцитам и сохраняющая антигены на своей поверхности. К ним относятся фолликулярные отросчатые клетки лимфоузлов и селезенки, клетки Лангерханса кожи и дыхательных путей, М- клетки лимфатических фолликулов пищеварительного тракта, дендритные эпителиальные клетки тимуса.
CD антигены.
Кластерная дифференциация поверхностных молекул (антигенов) клеток, прежде всего лейкоцитов, шагает далеко вперед. К настоящему времени CD антигены- не абстрактные маркеры, а функционально значимые для клетки рецепторы, домены и детерминанты, в том числе исходно не являющиеся специфическими для лейкоцитов.
Важнейшими дифференцировочными антигенами Т- лимфоцитов человека являются следующие.
1. CD2 - антиген, характерный для Т- лимфоцитов, тимоцитов, NK клеток. Он идентичен рецептору эритроцитов барана и обеспечивает образование розеток с ними (методика определения Т- клеток).
2. CD3 - необходимы для функционирования любых Т- клеточных рецепторов (ТКР). Молекулы CD3 имеют все субклассы Т- лимфоцитов. Взаимодействие ТКР- CD3 (она состоит из 5 субъединиц) с представляющей антиген молекулой МНС класса 1 или 2 определяет характер и реализацию иммунного ответа.
3. CD4. Эти рецепторы имеют Т- хелперы 1 и 2 и Т- индукторы. Являются корецептором (местом связывания) детерминант белковых молекул МНС класса 2. Является специфическим рецептором для оболочечных белков вируса иммунодефицита человека ВИЧ- 1 (gp120) и ВИЧ- 2.
4. CD8. Популяция CD8+ Т- лимфоцитов включает цитотоксические и супрессорные клетки. При контакте с клеткой- мишенью CD8 выступает в роли корецептора для белков HLA класса 1.
Дифференцировочные рецепторы В- лимфоцитов.
На поверхности В- лимфоцитов может находиться до 150 тысяч рецепторов, среди которых описано более 40 типов с различными функциями. Среди них - рецепторы к Fc- компоненту иммуноглобулинов, к С3 компоненту комплемента, антигенспецифические Ig рецепторы, рецепторы к различным факторам роста и дифференцировки.
Краткая характеристика методов оценки Т- и В- лимфоцитов.
Для выявления В- лимфоцитов используют метод розеткообразования с эритроцитами, обработанными антителами и комплементом (EAC- РОК), спонтанного розеткообразования с эритроцитами мыши, метод флюоресцирующих антител с моноклональными антителами (МКА) к рецепторам В- клеток (CD78, CD79a,b, мембранные Ig).
Для количественной оценки Т- лимфоцитов используют метод спонтанного розеткообразования с эритроцитами барана ( Е- РОК), для выявления субпопуляций ( например, Т- хелперов и Т- супрессоров) - иммунофлюоресцентный метод с МКА к CD рецепторам, для определения Т- киллеров- тесты цитотоксичности.
Функциональную активность Т- и В- клеток можно оценить в реакции бласттрансформации лимфоцитов (РБТЛ) на различные Т- и В- митогены.
Сенсибилизированные Т- лимфоциты, участвующие в реакциях гиперчувствительности замедленного типа (ГЗТ) можно определить по выделению одного из цитокинов - MIF (миграцию ингибирующего фактора) в реакции торможения миграции лейкоцитов (лимфоцитов) - РТМЛ. Подробнее о методах оценки иммунной системы- в лекциях по клинической иммунологии.
Одной из особенностей иммунокомпетентных клеток, особенно Т- лимфоцитов, является способность продуцировать большое количество растворимых веществ - цитокинов (интерлейкинов), осуществляющих регуляторные функции. Они обеспечивают согласованную работу всех систем и факторов иммунной системы, благодаря прямым и обратным связям между различными системами и субпопуляциями клеток обеспечивают устойчивую саморегуляцию иммунной системы. Их определение дает дополнительное представление о состоянии иммунной системы.
В целом гомеостаз организма обеспечивается согласованной работой (взаимодействием) иммунной, эндокринной и нервной систем.
Лекция № 14. Аллергия. ГНТ, ГЗТ. Особенности развития, методы диагностики. Иммунологическая толерантность.
Аллергические заболевания широко распространены, что связано с рядом отягощающих факторов - ухудшением экологической обстановки и широким распространением аллергенов, усилением антигенного давления на организм (в том числе- вакцинация), искусственным вскармливанием, наследственной предрасположенностью.
Аллергия ( allos + ergon, в переводе- другое действие) - состояние патологически повышенной чувствительности организма к повторному введению антигена. Антигены, вызывающие аллергические состояния, называют аллергенами. Аллергическими свойствами обладают различные чужеродные растительные и животные белки, а также гаптены в комплексе с белковым носителем.
Аллергические реакции - иммунопатологические реакции, связанные с высокой активностью клеточных и гуморальных факторов иммунной системы (иммунологической гиперреактивностью). Иммунные механизмы, обеспечивающие защиту организма, могут приводить к повреждению тканей, реализуясь в виде реакций гиперчувствительности.
Классификация Джелла и Кумбса выделяет 4 основных типа гиперчувствительности в зависимости от преобладающих механизмов, участвующих в их реализации.
По скорости проявления и механизму аллергические реакции можно разделить на две группы - аллергические реакции (или гиперчувствительность) немедленного типа (ГНТ) и замедленного типа (ГЗТ).
Аллергические реакции гуморального (немедленного) типа обусловлены главным образом функцией антител классов IgG и особенно IgE (реагинов). В них принимают участие тучные клетки, эозинофилы, базофилы, тромбоциты. ГНТ делят на три типа. По классификации Джелла и Кумбса к ГНТ относятся реакции гиперчувствительности 1, 2 и 3 типов, т.е. анафилактическая (атопическая), цитотоксическая и иммунных комплексов.
ГНТ характеризуется быстрым развитием после контакта с аллергеном (минуты), в ней участвуют антитела.
Тип 1. Анафилактические реакции - немедленного типа, атопические, реагиновые. Они вызываются взаимодействием поступающих извне аллергенов с антителами класса IgE, фиксированными на поверхности тучных клеток и базофилов. Реакция сопровождается активацией и дегрануляцией клеток- мишеней с высвобождением медиаторов аллергии (главным образом гистамина). Примеры реакций типа 1 - анафилактический шок, атопическая бронхиальная астма, поллиноз.
Тип 2. Цитотоксические реакции. В них участвуют цитотоксические антитела (IgM и IgG), которые связывают антиген на поверхности клеток, активируют систему комплемента и фагоцитоз, приводят к развитию антитело- зависимого клеточно- опосредованного цитолиза и повреждения тканей. Пример- аутоиммунная гемолитическая анемия.
Тип 3. Реакции иммунных комплексов. Комплексы антиген- антитела откладываются в тканях (фиксированные иммунные комплексы), активируют систему комплемента, привлекают к месту фиксации иммунных комплексов полиморфноядерные лейкоциты, приводят к развитию воспалительной реакции. Примеры- острый гломерулонефрит, феномен Артюса.
Гиперчувствительность замедленного типа (ГЗТ) - клеточно- опосредованная гиперчувствительность или гиперчувствительность типа 4, связанная с наличием сенсибилизированных лимфоцитов. Эффекторными клетками являются Т- клетки ГЗТ, имеющие CD4 рецепторы в отличие от CD8+ цитотоксических лимфоцитов. Сенсибилизацию Т- клеток ГЗТ могут вызывать агенты контактной аллергии (гаптены), антигены бактерий, вирусов, грибов, простейших. Близкие механизмы в организме вызывают антигены опухолей в противоопухолевом иммунитете, генетически чужеродные антигены донора- при трансплантационном иммунитете.
Т- клетки ГЗТ распознают чужеродные антигены и секретируют гамма- интерферон и различные лимфокины, стимулируя цитотоксичность макрофагов, усиливая Т- и В- иммунный ответ, вызывая возникновение воспалительного процесса.
Исторически ГЗТ выявлялась в кожных аллергических пробах (с туберкулином- туберкулиновая проба), выявляемых через 24 - 48 часов после внутрикожного введения антигена. Развитием ГЗТ на вводимый антиген отвечают только организмы с предшествующей сенсибилизацией этим антигеном.
Классический пример инфекционной ГЗТ - образование инфекционной гранулемы (при бруцеллезе, туберкулезе, брюшном тифе и др.). Гистологически ГЗТ характеризуется инфильтрацией очага вначале нейтрофилами, затем лимфоцитами и макрофагами. Сенсибилизированные Т- клетки ГЗТ распознают гомологичные эпитопы, представленные на мембране дендритных клеток, а также секретируют медиаторы, активирующие макрофаги и привлекающие в очаг другие клетки воспаления. Активированные макрофаги и другие участвующие в ГЗТ клетки выделяют ряд биологически активных веществ, вызывающих воспаление и уничтожающих бактерии, опухолевые и другие чужеродные клетки - цитокины (ИЛ-1, ИЛ-6, альфа- фактор некроза опухолей), активные метаболиты кислорода, протеазы, лизоцим и лактоферрин.
Методы лабораторной диагностики аллергии : выявление уровня сывороточных IgE, фиксированных на базофилах и тучных клетках антител класса Е (реагинов), циркулирующих и фиксированных (тканевых) иммунных комплесов, провокационные и кожные пробы с предполагаемыми аллергенами, выявление сенсибилизированных клеток тестами in vitro - реакция бласттрансформации лимфоцитов (РБТЛ), реакция торможения миграции лейкоцитов (РТМЛ), цитотоксические тесты.
Иммунологическая толерантность.
Иммунологическая толерантность- специфическое подавление иммунного ответа, вызванное предварительным введением антигена. Иммунологическая толерантность как форма иммунного ответа специфична.
Толерантность может проявляться в подавлении синтеза антител и гиперчувствительности замедленного типа (специфического гуморального и клеточного ответа) или отдельных видов и типов иммунного ответа. Толерантность может быть полной (нет иммунного ответа) или частичной (существенное снижение ответа).
Если на введение антигена организм отвечает подавлением только отдельных компонентов иммунного ответа, то это - иммунологическое отклонение (расщепленная толерантность). Наиболее часто выявляется специфическая ареактивность Т- клеток (обычно Т- хелперов) при сохранении функциональной активности В- клеток.
Естественная иммунологическая толерантность - иммунологическая ареактивность к собственным антигенам (аутоиммунная толерантность) возникает в эмбриональном периоде. Она предотвращает выработку антител и Т- лимфоцитов, способных разрушать собственные ткани.
Приобретенная иммунологическая толерантность - отсутствие специфической иммунной реакции к чужеродному антигену.
Иммунологическая толерантность представляет особую форму иммунного ответа, характеризующуюся запретом, налагаемым Т- и В- супрессорами на образование клеток- эффекторов против данного, в том числе собственного, антигена (А.И.Коротяев, С.А.Бабичев, 1998).
В основе индуцированной иммунологической толерантности лежат различные механизмы, среди которых принято выделять центральные и периферические.
Центральные механизмы связаны с непосредственным воздействием на иммунокомпетентные клетки. Основные механизмы:
- элиминация антигеном иммунокомпетентных клеток в тимусе и костном мозге (Т- и В- клеток соответственно);
- повышение активности супрессорных Т- и В- клеток, недостаточность контрсупрессоров;
- блокада эффекторных клеток;
- дефектность презентации антигенов, дисбаланс процессов пролиферации и дифференциации, кооперации клеток в иммунном ответе.
Периферические механизмы связаны с перегрузкой (истощением) иммунной системы антигеном, пассивным введением высокоаффинных антител, действием антиидиотипических антител, блокадой рецепторов антигеном, комплексом “антиген- антитела”, антиидиопатическими антителами.
Исторически иммунологическую толерантность рассматривают как защиту против аутоиммунных заболеваний. При нарушении толерантности к собственным антигенам могут развиваться аутоиммунные реакции, в том числе возникать такие аутоиммунные заболевания как ревматоидный артрит, системная красная волчанка и другие.
Основные механизмы отмены толерантности и развития аутоиммунных реакций
1. Изменения химической структуры аутоантигенов (например- изменение нормальной структуры антигенов клеточных мембран при вирусных инфекциях, появление ожоговых антигенов).
2. Отмена толерантности на перекрестно- реагирующие антигены микроорганизмов и эпитопы аутоантигена.
3. Появление новых антигенных детерминант в результате связывания чужеродных антигенных детерминант с клетками хозяина.
4. Нарушение гисто- гематических барьеров.
5. Действие суперантигенов.
6.
Нарушения регуляции иммунной системы ( уменьшение количества или
функциональная недостаточность супрессирующих лимфоцитов, экспрессия молекул
МНС класса 2 на клетках, в норме их не экспрессирующих- тиреоциты при
аутоиммунном тиреоидите).
Лекция № 15. Иммунный статус организма. Методы его оценки.
Состояние иммунной системы имеет важнейшее значение в обеспечении гомеостаза организма, защите от всего генетически чужеродного.
Иммунный статус - это совокупность лабораторных показателей, характеризующих количественную и функциональную активность клеток иммунной системы.
Цель иммунологической диагностики - выявление направленности значимых нарушений в иммунной системе и прогнозирование вероятных последствий различных воздействий на организм человека и популяцию в целом. Задачи ОИС:
1. Выявление дефектов в работе иммунной системе до клинического проявления иммунодефицитного состояния (донозологическая диагностика).
2. Определение нарушенного звена иммунитета у больных с различными расстройствами иммунной системы для подтверждения диагноза и проведения соответствующей иммунотерапии.
Иммунный статус определяет эффективность и согласованность работы всех систем и звеньев иммунитета - макрофагов, комплемента, интерферонов, Т- и В- лимфоцитов, главной системы гистосовместимости. Раздел медицины, изучающий патологию человека в аспекте нарушений функций иммунной системы, называется клинической иммунологией.
Для постановки диагноза иммунопатологического состояния проводят сбор иммунологического анамнеза и постановку иммунологических тестов. Могут также осуществляться тесты in vivo (кожные тесты), ретгенологическое исследование лимфоидных органов (тимуса).
При опросе определяют наиболее вероятный иммунопатологический синдром, среди которых основными являются шесть:
-инфекционный синдром; - аллергический синдром;
- аутоиммунный синдром;
- первичный иммунодефицит; - вторичный иммунодефицит;
- иммунопролиферативный синдром.
Для оценки общего иммунного статуса используют наиболее простые и достоверные показатели, отражающие суммарную эффективность работы всех систем иммунитета, для изучения уязвимого звена - специфичные для каждой системы дифференциальные тесты.
Изучение иммунного статуса проводится не менее чем в два этапа. Р.В.Петров с соавторами в 1984г. создали двухэтапный подход к оценке иммунного статуса, в соответствии с которым лабораторные иммунологические тесты разделены на тесты первого и второго уровня.
На первом этапе с помощью простых ориентировочных методов выявляют “грубые” дефекты фагоцитоза, клеточного и гуморального иммунитета. К тестам первого уровня относят:
✓ определение относительного и абсолютного количества лейкоцитов, нейтрофилов, моноцитов, лимфоцитов и тромбоцитов периферической крови;
✓ определение функциональной активности нейтрофилов (НСТ тест);
✓ тесты иммунофенотипирования для определения относительного и абсолютного количества Т- и В-лимфоцитов, натуральных киллеров;
✓ определение концентрации иммуноглобулинов основных классов (IgA,
IgM, IgG, IgE); ✓ определение гемолитической активности комплемента.
С помощью минимального набора тестов можно диагностировать первичные иммунодефициты: хроническую грануломатозную болезнь, Х-сцепленную агаммаглобулинемию, гипер–IgM–синдром, селективный дефицит IgA, синдром ВискоттаОлдрича, тяжелый комбинированный иммунодефицит.
С учетом анализа результатов тестов 1 уровня определяют дальнейшую тактику иммунологического исследования.
Более тщательный и глубокий анализ состояния иммунной системы проводят с помощью тестов второго уровня - аналитических методов. К ним можно отнести методы оценки функциональной активности Т- и В- лимфоцитов, фагоцитов, вспомогательных клеток, естественных киллеров, компонентов системы комплемента и многих других.
✓тесты иммунофенотипирования для определения относительного и абсолютного количества субпопуляций Т-, В-, NK-лимфоцитов;
✓активационные маркеры лимфоцитов;
✓оценка различных этапов фагоцитоза и рецепторного аппарата фагоцитарных клеток;
✓циркулирующие иммунные комплексы;
✓определение концентрации компонентов комплемента в сыворотке крови (С3, С4, С5, С1-ингибитор);
✓функциональная активность различных субпопуляций лимфоцитов;
✓оценка пролиферативной активности Т- и В-лимфоцитов; ✓ исследование интерферонового статуса; ✓ кожные пробы и т.д.
Набор полученных показателей при иммунологическом обследовании называется иммунограммой.
Иммунограмма – это способ оценки иммунного статуса, который включает комплекс лабораторных методов: развернутую лейкоформулу, определение показателей системы неспецифической резистентности, исследование клеточного и гуморального звеньев иммунитета.
Как известно, попадание в организм антигена вызывает, по крайней мере, два вида иммунного ответа – клеточный и гуморальный.
Под клеточным подразумевают способность CD4Т-лимфоцитов распознавать антиген, находящийся на АГ – представляющих клетках и реагировать на него синтезом ряда цитокинов, ведущих к образованию антиген-специфичных CD4 и CD8-лимфоцитов и активации клеток моноцитарно-макрофагального ряда.
Под гуморальным - подразумевают синтез специфических иммуноглобулинов – антител В – лимфоцитами, в результате их активации антигеном, находящимся на АГП-клетках и цитокинами, продуцируемыми CD4 - лимфоцитами.
К основным клеткам, принимающим участие в иммунном ответе относятся все типы лейкоцитов – лимфоциты, нейтрофилы сегментоядерные и палочкоядерные, моноциты, базофилы, эозинофилы. Однако, специфичность иммунной реакции организма определяют Т и В-лимфоциты, направляющие эффекторную функцию всего комплекса клеток в нужную сторону.
Методы исследования главных компонентов иммунной системы принято делить также на скрининговые и развернутые.
При оценке В- системы иммунитета к скрининговым тестам относятся: определение количества CD19+ и CD20+ клеток, IgG, IgM и IgA, а также IgG1,2,3,4, к развернутым- бласттрансформацию на митоген лаконоса и S.aureus, поверхностных маркеров В- лимфоцитов.
При оценке Т- системы иммунитета к скрининговым методам можно отнести кожные тесты на бактериальные антигены, определение поверхностных маркеров Т- лимфоцитов CD3, CD4, CD8, бласттрансформацию на фитогемагглютинин, к развернутым- изучение продукции цитокинов, активационных маркеров, Т- клеточных рецепторов.
При оценке фагоцитоза к скрининговым тестам относят определение количества нейтрофилов, изучение их морфологии и образования активных форм кислорода, к развернутым- определение киллинга микробов, лизосомальных ферментов, цитокинов.
Существующие методы оценки иммунного статуса постоянно совершенствуются, однако есть ряд общих правил, которых необходимо придерживаться при оценке иммунограмм:
- комплексный анализ, а не оценка одного показателя;
- анализ в комплексе с клиническими и анамнестическими данными;
- оценка резких сдвигов показателей (не менее 20% от нормы);
- анализ в динамике;
- анализ не только (и не столько) абсолютных данных, а соотношений показателей (особо- индекс Th/Ts);
- учет возможных индивидуальных особенностей (возраст, сопутствующие заболевания) и колебаний показателей (физиологических и патологических- прием пищи, физические нагрузки, время суток, действие стрессоров и т.д.);
- учет региональных норм;
- учет материально- технической базы лаборатории. На вооружении современной лаборатории- проточные цитометры и другое оборудование, которое позволяет наиболее точно и подробно определить иммунный статус (лазерная проточная цитофлюориметрия).
Методы исследования лимфоцитов можно разделить на изучение поверхностных маркеров и функциональные тесты.
Изучение поверхностных СД антигенов основывается на:
- методах иммунофлюоресценции (метод моноклональных антител); - иммуноферментном анализе.
Важнейшее значение имеет вычисление индекса CD4/CD8 (хелперно- супрессорного отношения). Необходимо учитывать, что нет единого метода для оценки Т- хелперов и Т- супрессоров. CD8+ несут Т- супрессоры и Т- киллеры, часть NK- клеток.
CD4+ несут Т- хелперы и Т- индукторы, моноциты, эффекторы- Т- клетки ГЗТ.
К функциональным тестам относят методы оценки пролиферативной активности лимфоцитов на Т- и В- митогены (РБТЛ- реакция бластной трансформации лимфоцитов), продукции антител, синтеза мононуклеарами цитокинов (особо!).
Следует особо подчеркнуть, что полноценный анализ иммунограммы возможен только в комплексе с клиническим состоянием и анамнезом пациента. Отсутствие характерных сдвигов в иммунограмме при выраженных клинических симптомах следует считать атипичной реакцией иммунной системы, что является отягощающим признаком заболевания. Полученные данные пациента сравниваются со средними значениями для данного аналита, полученными в регионе проживания пациента. Среднестатистические показатели различаются в зависимости от региона и подчиняются климатогеографическим условиям, экологической обстановке, условиям жизни. Необходимо также учитывать возраст пациента и циркадные ритмы.
Исследование показателей ИС имеет большое значение для диагностики и дифференциальной диагностики, особенно при первичных иммунодефицитах и лимфопролиферативных заболеваниях, для оценки тяжести, активности, длительности течения и прогноза различный заболеваний, оценки эффективности проводимого лечения.
Первичное иммунологическое обследование позволяет создать нормативные параметры иммунного статуса населения с учетом ведущих факторов, действующих в регионе: климатогеографических, национальных особенностей, экологической ситуации и эпидемиологической обстановки на момент обследования. Причем, необходимо иметь нормы региональные, но и для данной лаборатории, поскольку могут быть применены разные методы и их модификации, внесены различные по характеристикам реактивы и оборудование даже при использовании одних и тех же тестов.
Следует отметить, что нормальным является не только спокойное функционирование иммунной системы у здоровых людей, но и активная ее работа при большинстве благоприятно текущих воспалительных заболеваний. Из этого следует, что изменения показателей иммунограммы при благоприятном развитии воспалительного процесса на разных его этапах характеризуют нормальное функционирование иммунной системы, а значит, являются показателями «нормы» активного функционирования иммунной системы. Именно отталкиваясь от этих динамически изменяющихся нормативных показателей, характеризующих степень активации иммунной системы, а не узкой «нормы» иммунограммы только у клинически здорового человека, можно выявить недостаточность работы иммунной системы, и на основании этого правильно назначить препараты для ее нормализации при том или ином заболевании.
Пока клиницисты не располагают
данными о паспорте здоровья каждого человека, существенным подспорьем в их
работе могут служить результаты массовых обследований здоровых жителей
конкретных регионов, городов.
Лекция № 16. Иммунодефициты.
Иммунная система обладает особыми физиологическими механизмами функционирования (распознавание антигена, активация иммунокомпетентных клеток, их пролиферация, дифференцировка и иммунорегуляция). Если в одном или нескольких звеньях иммунной системы возникают дефекты, это приводит к иммунодефицитным состояниям.
По происхождению различают первичные (генетически обусловленные) и вторичные (возникающие в связи с инфекциями, инвазиями, опухолями, старением, ожогами, травмами и др.) иммунодефициты.
В зависимости от уровня дефекта выделяют:
- иммунодефициты, обусловленные преимущественным поражением В- звена; - иммунодефициты, обусловленные преимущественным поражением Т- звена; - комбинированные иммунодефициты.
Различают также гуморальные (самые частые), клеточные и клеточно- гуморальные иммунодефициты.
Дефицит лимфоцитов, макрофагов, плазмацитов, гранулоцитов - это клеточная форма иммунодефицита. Дефицит иммуноглобулинов (антител) - это гуморальный иммунодефицит.
Первичные (врожденные) иммунодефициты.
В основе первичных (врожденных) иммунодефицитов - генетический дефект, который может реализоваться на разных стадиях развития иммунокомпетентных клеток - стволовой клетки, этапах дифференциации Т- и В- клеток, при созревании плазматических клеток. Спектр хромосомных дефектов иммунитета достаточно широк - мутации, нарушения транскрипции и трансляции, генетически детерминированные дефекты мембран клеток. Особое значение придается аномалиям короткого плеча X - хромосомы, с которыми связан механизм иммунорегуляции. С областью HLA-D сцеплены гены Ir (силы иммунного ответа) и Is (супрессии иммунного ответа). Около трети первичных иммунодефицитов сцеплены с полом и передаются по наследству.
Проблема врожденных иммунодефицитов- это преимущественно проблема педиатрии, только в последние десятилетия после разработки методов диагностики и лечения первичных иммунодефицитов стало возможным продлевать жизнь этим больным (проблема оппортунистических инфекций, опухолей и т.д.). Иммунодефициты могут быть связаны с нарушением любого из звеньев иммунитета (т.е. не только Т- и В- лимфоцитов, но и макрофагов, комплемента, главной системы гистосовместимости, интерлейкинов и др. Наиболее часто выделяют иммунодефициты, обусловленные:
- нарушениями гуморального звена иммунитета (гипо- и агаммаглобулинемии и
др.;
- нарушениями функций тимуса и клеточного иммунитета;
- нарушениями в системе фагоцитоза;
- дефектами системы комплемента;
- тяжелыми комбинированными нарушениями.
Из числа первичных иммунодефицитов приведем наиболее яркие примеры.
1. Болезнь Брутона - наследственная , сцепленная с полом гипогаммаглобулинемия, обусловленная дефектом В- клеток. Болеют мальчики от клинически здоровой матери ( рецессивный тип наследственности, сцепленный с полом). Не вырабатываются антитела против бактериальных инфекций, необходима защита иммуноглобулинами и антибиотиками.
2. Швейцарская агаммаглобулинемия - тяжелый комбинированный Т- и В- иммунодефицит, связанный с нарушениями на уровне стволовой клетки. Снижено количество Т- клеток и иммуноглобулинов основных классов. Наследование аутосомно- рецессивное (болеют и мальчики и девочки) или рецессивное, сцепленное с полом. Отмечается гипоплазия тимуса и лимфоузлов, экзантемы, желудочно- кишечные расстройства и пневмонии. Дети редко доживают до трех лет, лечение- пересадкой костного мозга.
3. Синдром Луи- Бар (атаксия- телеангиэктазия). Связан с дефектом тимуса и мутациями в 7 и 14 хромосомах. Уменьшено количество Т- клеток (преимущественно Т- хелперов) и В- клеток. Клинически- нарушение координации движений, дебильность, телеангиэктазии, инфекции дыхательных путей (отсутствие IgA), опухоли лимфоидной ткани.
4. Синдром третьего и четвертого глоточных мешков (синдром Ди Джорджи). Наблюдается аплазия тимуса и паращитовидных желез. Синтез иммуноглобулинов осуществляется нормально. Клеточный иммунитет отсутствует, что делает таких больных восприимчивыми к вирусным инфекциям и грибковым поражениям. Ранний признак болезни - тетания, обусловленная дефицитом кальция.
5. Хронический гранулематоз - заболевание, связанное с нарушениями лизосомальных ферментов. Фагоцитоз осуществляется, однако киллинг и переваривание микроорганизмов не происходит, отсутствует способность восстанавливать нитросиний тетразолий (НСТ). У больных выявляют рецидивирующие стафилококковые абсцессы, отиты и другие гнойно- воспалительные заболевания. Болезнь обусловлена наследственным X - сцепленным аутосомным дефектом кислородного взрыва в нейтрофилах и моноцитах (макрофагах).
При всех формах первичных иммунодефицитов повышена частота злокачественных новообразований, особенно лимфоретикулярной системы. Опухоли иммунной системы подразделяются на В- клеточные, Т- клеточные и связанные с усиленной пролиферацией “нулевых” лимфоцитов (третьей популяции). Возникая в результате нарушения “надзорной” функции иммунитета, злокачественные опухоли сами становятся основой развития тяжелого вторичного иммунодефицита.
К общим проявлениям иммунодефицитов относятся:
- инфекционный синдром (гнойно- септические процессы связаны с нарушениями преимущественно гуморального иммунитета, оппортунистические вирусные, грибковые и протозойные заболевания - с дефектами клеточного иммунитета);
- желудочно- кишечные расстройства (нарушения всасывания, дефицит IgA, инфекции желудочно- кишечного тракта);
- опухоли иммунной системы;
- аллергический и аутоиммунный синдромы (атопии, аутоиммунные гемолитические анемии);
- частое сочетание с пороками развития (при врожденных иммунодефицитах);
- гематологические изменения (снижение количества лимфоцитов и нейтрофи-
лов, эозинофилия, анемия, тромбоцитопения).
Вторичные (приобретенные) иммунодефициты.
Вторичные или приобретенные иммунодефициты возникают вследствие какого- либо тяжелого заболевания (т.е. как правило при ранее нормальном иммунном статусе). К основным причинам возникновения вторичных иммунодефицитов можно отнести следующие.
1. Паразитарные и протозойные болезни (описторхоз, малярия, шисто- и трипаносомозы, трихинеллез и др.).
2. Вирусные инфекции - наиболее крупная группа инфекционных агентов, вызывающих иммунодефициты:
- внутриутробные инфекции (цитомегаловирусная инфекция, краснуха);
- острые инфекции (корь, грипп, краснуха, паротит, ветряная оспа, вирусные гепатиты);
- персистирующие (гепатит В и С, герпес);
- инфекции иммунной системы (ВИЧ, ЦМВ, вирус Эпштейн - Барр).
3. Бактериальные инфекции (туберкулез, сифилис, лепра).
4. Хирургические вмешательства, травмы.
5. Ожоги.
6. Нарушения обмена веществ (сахарный диабет) и истощение (голодание).
7. Заболевания органов выделения (уремия).
8. Опухоли.
9. Хронические соматические заболевания.
10. Действие лекарств, экологических и производственных факторов, радиации. Кратко дадим характеристику основных видов вторичных иммунодефицитов.
Дефекты иммунного статуса при паразитарных и протозойных заболеваниях связаны с рядом механизмов :
- угнетением функции макрофагов (малярия);
- выработкой лимфоцитотоксинов (описторхоз, трихинеллез);
- выработкой супрессивно действующих факторов (трипано- и шистосомозы); - различными нарушениями иммунорегуляции.
Иммунодефициты при бактериальных инфекциях. Часто наблюдается снижение Т- лимфоцитов и митогенной активности на фитогемагглютинин (ФГА)- лепра, туберкулез, сифилис, пневмококковые инфекции, коклюш, бруцеллез, скарлатина). При стрепто- и стафилококковых инфекциях подавление Т- звена иммунитета часто сочетается с повышением функции В- системы и формированием инфекционно- аллергических и аутоиммунных осложнений (заболеваний).
Иммунодефициты при вирусных инфекциях. Многие вирусы вызывают резкое угнетение Т- звена иммунитета ( вирусы кори, краснухи, гриппа, паротита). При кори и гриппе это нарушение сочетается с дефектами фагоцитоза, что еще более угнетает противомикробную защиту и способствует присоединению бактериальных осложнений. Однако наиболее существенные нарушения иммунной системы вызывают вирусы, непосредственно поражающие иммунную систему.
1. Вирус иммунодефицита человека (ВИЧ) вызывает заболевание, которое называют “синдром приобретенного иммунодефицита (СПИД)”. Этот вирус относится к ретровирусам и имеет тропизм к клеткам иммунной системы и некоторым другим клеткам, несущим CD4+ рецептор. CD4 является фактически рецептором для ВИЧ, благодаря которому РНК вируса попадает (инфицирует) клетки, формирует ДНК- копию, которая встраивается в ДНК (геном) клетки хозяина и получает возможность реплицироваться. Вирус оказывает на клетки цитопатический эффект, вызывая поражение Т- хелперов и других СД4+ клеток, снижение индекса CD4/CD8, глобальный дефект гуморального и клеточного иммунитета в сочетании с поликлональной активацией В- лимфоцитов, резкое ослабление противоинфекционной и противоопухолевой защиты. Парадокс- прогрессирование болезни (иммунодефицита) на фоне активного антительного ответа и ГЗТ на ВИЧ. На этом фоне присоединяются оппортунистические (СПИД- ассоциированные) вторичные инфекции и инвазии (ЦМВ, гепрес, пневмоцистоз, токсоплазмоз, микоплазмоз и др.).
2. Цитомегаловирус. ЦМВ инфекция приводит к резкому снижению CD4+ Т- лимфоцитов и гиперактивности CD8+ Т- клеток, угнетению клеточного иммунитета. ЦМВ относится к семейству герпес- вирусов , часто вызывающих персистентные инфекции и развитие вторичных иммунодефицитов.
3. Вирус Эпштейн- Барр вызывает инфекционный мононуклеоз. Рецептором для этого вируса является CD21- рецептор, поэтому поражаются преимущественно В- клетки. Эти же рецепторы имеются на дендритных клетках лимфоидных фолликулов, цервикальном эпителии. СД21 рецептор является местом присоединения С3d- компонента комплемента. Присоединение к рецептору вируса Эпштейн- Барр вызывает экспрессию на мембране В- лимфоцитов особого антигена, распознаваемого СД8+ лимфоцитами как чужеродного. В результате В- клетки становятся мишенью для собственных Т- клеток. В крови определяется атипичный Т- лимфоцитоз, бласттрансформация В- клеток, выработка гетерофильных антител. Формируется сложный иммунодефицит с элементами аутоагрессии.
Иммунодефициты при ожогах усугубляют опасность инфекционных осложнений. В первые дни преобладает снижение иммуноглобулинов основных классов (особенно IgG). В дальнейшем действие ожоговых антигенов приводит к В- клеточной стимуляции. Отмечено снижение ряда показателей Т- клеточного иммунитета в результате действия ожоговых токсинов- CD3+ и CD4+ клеток, ингибируется фагоцитоз, снижается активность комплемента. Прогностически неблагоприятен дисбаланс соотношения CD4/CD8.
Иммунодефициты, связанные с недостаточностью питания, голоданием, нарушениями обмена веществ.
При сахарном диабете возникает предрасположенность к бактериальным инфекциям, связанная с нарушением функций лейкоцитов- хемотаксиса, адгезивных и бактерицидных свойств.
Дефицит белка в организме также повышает восприимчивость к инфекциям. Угнетается первичный иммунный ответ (синтез IgM), фагоцитарная активность клеток, митогенная активность (по данным РБТЛ с ФГА).
Дефицит микроэлементов существенно сказывается на иммунной системе. Дефицит железа ведет к снижению активности железосодержащих ферментов, Т- звена, уровня миелопероксидазы и АФК. Дефицит цинка ведет к гипофункции тимуса со снижением CD4+ Т- лимфоцитов, ответа на митогены, активности NK клеток, фагоцитарного звена. Дефицит лития ведет к недостаточности Т- лимфоцитов, особенно CD8+ клеток. Существенно сказывается на иммунную систему дефицит меди, селена, кальция, магния. С дефицитом магния связаны нарушения синтеза антител, активации системы комплемента.
Лекарственные иммунодефициты связаны преимущественно с их иммунотоксическим действием. Достаточно часто отмечается активация Т- супрессоров, уменьшение количества В- клеток, снижение IgA. Существенное влияние на иммунный статус оказывают антибиотики, даже при коротких циклах применения, прежде всего - пенициллины, тетрациклины, стрептомицин, противотуберкулезные и антигрибковые препараты. Они вызывают :
- дефекты формирования первичного иммунного ответа (скорости образования клона плазматических клеток и антителообразования);
- снижение противовирусной защиты;
- снижение цитотоксической активности Т- лимфоцитов;
- уменьшение фагоцитарной активности нейтрофилов и макрофагов.
Около 70 тысяч химических соединений, связанных с производственной деятельностью и нарушениями экологической обстановки относятся к разряду токсических и оказывают разнообразное действие на иммунную систему.
Существенное влияние на иммунную систему оказывает стресс. Начальный период острого стресса характеризуется снижением противоопухолевого и противоинфекционного иммунитета, в дальнейшем могут присоединяться аутоиммунные и аллергические реакции. Хронический стресс неизбежно приводит к формированию вторичного иммунодефицита.
Существенное меняется иммунный статус при старении. Отмечается инволюция тимуса, снижается уровень тимического фактора. С возрастом снижается активность клеточного иммунитета, страдают этапы распознавания антигена, пролиферативная активность Т- клеток, изменяется CD4/CD8 индекс в сторону супрессорной активности, страдают надзорные функции противоопухолевой защиты.
В отличии от первичных иммунодефицитов, вторичные иммунодефициты в большинстве случаев не носят необратимого характера и функции иммунной системы могут восстанавливаться, если прекращается действие факторов, обословивших иммунодефицит.
Лекция № 17. Основы иммунотерапии и иммунопрофилактики.
Иммунотерапия- метод лечения, при котором осуществляется воздействие на иммунную систему : подавление иммунного ответа (иммуносупрессия), стимуляция ответа (иммуностимуляция), восстановление иммунодефицитов (иммунокоррекция). В прикладном, более узком смысле иммунотерапия использует специфические методы серотерапии (применение иммунных сывороток, иммуноглобулинов), вакцинотерапии (лечебные вакцины), иммунокоррекции (десенсибилизация и др.).
Иммунопрофилактика - способ предупреждения инфекционных заболеваний путем создания искусственного специфического иммунитета. Выделяют вакцинопрофилактику (создание активного иммунитета за счет вакцин, антигенов) и серопрофилактику (пассивный иммунитет за счет введения в организм специфических антител - иммуноглобулинов). Основную роль в специфической профилактике инфекционных заболеваний имеет вакцинопрофилактика.
Вариоляция - ранее применявшийся способ защиты от натуральной оспы с помощью втирания в кожу небольшого количества заразного материала от выздоравливающих от оспы людей известен с незапамятных времен. В России одной из первых этой процедуре подверглась Екатерина II. Однако способ вариоляции был очень опасным.
Вакцинация. Вакцинацией человечество обязано Э.Дженнеру, который в 1796г. показал, что прививка коровьей оспы - вакцинация (vaccinum - с лат. коровий) эффективна для профилактики натуральной оспы. С тех пор препараты, используемые для создания специфического активного иммунитета, называют вакцинами.
Существует ряд типов вакцин - живые, убитые, компонентные и субъединичные, рекомбинантные, синтетические олигопептидные, антиидиотипические и др.
1. Убитые (инактивированные) вакцины - это вакцинные препараты, не содержащие живых микроорганизмов. Вакцины могут содержать цельные микробы (корпускулы) - вакцины против чумы, гриппа, полиомиелитная вакцина Солка, а также отдельные компоненты (полисахаридная пневмококковая вакцина) или иммунологически активные фракции (вакцина против вируса гепатита В).
Различают вакцины, содержащие антигены одного возбудителя (моновалентные) или нескольких возбудителей (поливалентные).Убитые вакцины как правило менее иммуногенны, чем живые, реактогенны, могут вызывать сенсибилизацию организма.
2. Ослабленные (аттенуированные) вакцины. Эти вакцины имеют некоторые преимущества перед убитыми. Они полностью сохраняют антигенный набор микроорганизма и обеспечивают более длительное состояние специфической невосприимчивости. Живые вакцины применяют для профилактики полиомиелита, туляремии, бруцеллеза, кори, желтой лихорадки, эпидемического паротита. Недостатки - наличие не только нужных (протективных), но и вредных для организма антигенных комплексов ( в том числе перекрестно реагирующих с тканями человека), сенсибилизация организма, большая антигенная нагрузка на иммунную систему и др.
3. Компонентные (субъединичные) вакцины состоят из главных (мажорных) антигенных компонентов, способных обеспечить протективный иммунитет.
- компоненты структур клетки (антигены клеточной стенки, Н - и Vi - антигены, рибосомальные антигены);
- анатоксины - препараты, содержащие модифицированные химическим путем экзотоксины, лишенные токсических свойств, но сохранившие высокую антигенность и иммуногенность. Эти препараты обеспечивают выработку антитоксического иммунитета (антитоксических антител - антитоксинов). Наиболее широко используются дифтерийный и столбнячный анатоксины. АКДС - ассоциированная коклюшно- дифтерийно- столбнячная вакцина. Полученные химическим путем вакцинные препараты (пример- анатоксины, получаемые обработкой экзотоксинов формалином) называют химическими вакцинами;
- конъюгированные вакцины- комплекс малоиммуногенных полисахаридов и высокоиммуногенных анатоксинов- например, сочетание антигенов Haemophilus influenzae и обеспечивающего иммуногенность вакцины дифтерийного анатоксина;
- субъединичные вакцины. Вакцину против вируса гепатита В готовят из поверхностных белков (субъединиц) вирусных частиц (HBs антиген). В настоящее время эту вакцину получают на рекомбинантной основе- с помощью дрожжевых клеток с плазмидой, кодирующей HBs антиген.
4. Рекомбинантные вакцины. С помощью методов генной инженерии гены, контролирующие синтез наиболее значимых иммуногенных детерминант, встраивают в самореплицирующиеся генетические структуры (плазмиды, вирусы). Если носителем (вектором) является вирус осповакцины, то данная вакцина будет в организме индуцировать иммунитет не только против оспы, но и против того возбудителя, чей ген был встроен в его геном (если ген HBs антигена - против вируса гепатита В).
Если вектором является плазмида, то при размножении рекомбинантного клона микроорганизма (дрожжей, например) нарабатывается необходимый антиген, который и используется в дальнейшем для производства вакцин.
5. Синтетические олигопептидные вакцины. Принципы их конструирования включают синтез пептидных последовательностей, образующих эпитопы, распознаваемые нейтрализующими антителами.
6. Кассетные или экспозиционные вакцины. В качестве носителя используют белковую структуру, на поверхности которой экспонируют (располагают) введенные химическим или генно- инженерным путем соответствующие определенные антигенные детерминанты. В качестве носителей при создании искусственных вакцин могут использовать синтетические полимеры- полиэлектролиты.
7. Липосомальные вакцины. Они представляют собой комплексы, состоящие из антигенов и липофильных носителей (пример- фосфолипиды). Иммуногенные липосомы более эффективно стимулируют выработку антител, пролиферацию Т- лимфоцитов и секрецию ими ИЛ- 2.
8. Антиидиопатические вакцины. Антиидиотипические антитела содержат “внутренний” специфический портрет антигенной детерминанты. Получают моноклональные антиидиотипические антитела, содержащие “внутренний образ” протективного антигена. Для оптимальных результатов (защиты в отношении возбудителя) необходимо иметь набор МКА против различных антигенных детерминант возбудителя.
В настоящее время в нашей стране производится 7 анатоксинов, около 20 противовирусных и более 20 антибактериальных вакцин. Часть из них является ассоциированными - т.е. содержащими антигены различных возбудителей, или одного, но в различных вариантах (корпускулярные и химические).
Иммуномодулирующая терапия. Способы иммуномодуляции условно можно разделить на методы иммуностимуляции и иммунодепрессии.
Большинство иммунотропных препаратов подробно описано в фармацевтических справочниках. Однако при их применении необходимо придерживаться некоторых общих правил.
1. Решение о применении препаратов должно базироваться как на клинических проявлениях иммунодефицита, так и на данных лабораторных исследований.
2. Даже при положительном клиническом эффекте обязательно должно проводиться оценка иммунного статуса в динамике.
3. Необходимо строго придерживаться принятых схем и дозировок.
4. Результат действия может зависеть как от исходного состояния, так и от дозы препарата, т.е. на один и тот же препарат может быть как стимуляция, так и супрессия.
Иммуностимуляторы. Иммуностимулирующей активностью обладают препараты тимуса и их синтетические аналоги, левамизол (декарис), цитокины, препараты адамантанового ряда, некоторые соли, природные соединения, полиэлектролиты.
К стимуляторам Т- лимфоцитов относятся тактивин, тималин, тимоген, тимоптин, вилозен, декарис, диуцифон, нуклеинат натрия, цинка ацетат, спленин, к стимуляторам В- лимфоцитов - лиелопид, продигиозан, пирогенал. Стимуляторами фагоцитоза являются нуклеинат натрия, метилурацил (последний стимулирует также Т- и В- лимфоциты). К стимуляторам эндогенного интерферона относят дибазол и арбинол. Для заместительной терапии применяют иммуноглобулин для внутривенного введения, пентаглобулин (препарат IgM). Синтезирован ряд новых препаратов - различные цитокины, иммунофан, полиоксидоний. Определенным иммуностимулирующим действием обладают биогенные стимуляторы (адаптогены)- экстракт алоэ, ФИБС, стекловидное тело, сок каланхоэ, препараты женьшеня, пантокрина, радиолы розовой, элеуторококка, чабреца, чаги.
Иммунодепресанты. К препаратам с противовоспалительным и иммунодепрессивным действием относятся глюкокортикоидные гормоны. Большинство иммунодепрессантов является цитостатиками и часто применяются для химиотерапии злокачественных новообразований. Среди них выделяют антиметаболиты, алкилирующие препараты, антибиотики, алкалоиды и ингибиторы ферментов.
Антиметаболиты чаще всего влияют на обмен нуклеиновых кислот. К антогонистам пурина относятся меркаптопурин и азатиоприн (имуран).
К алкилирующим препаратам относят циклофосфамид, хлорбутин. Основной их мишенью являются белки и нуклеиновые кислоты, с которыми они ковалентно связываются. Нарушаются процессы репликации и трансляции, нарушаются процессы митоза клеток.
Антибиотики. Многие антибиотики оказывают влияние на обмен ДНК и РНК. В наибольшей степени это относится к продуктам деятельности актиномицет- актиномицинам С и Д , а также продукту жизнедеятельности грибов Trihoderma polysporium - циклоспорину. Актиномицин Д тормозит деление клеток и ДНК- зависимый синтез РНК. Актиномицин С является алкилирующим препаратом. Циклоспорин является активным иммунодепресантом, подавляющим клеточные иммунные реакции, в т.ч. реакции трансплантационного иммунитета, ГЗТ, Т- зависимое антителообразование. Механизм его действия связан с подавлением продукции Т- хелперами ИЛ-
2.
Применение иммунодепресантов, особенно цитостатиков, вызывает много осложнений: угнетение гемопоэза, противоинфекционной, противоопухолевой защиты.
Несмотря на обширный спектр иммуномодуляторов, подавляющее
число из них на практике используется редко. Причины- недостаточная
эффективность, побочные эффекты, токсичность, стоимость, недостаточная
изученность и др.
Лекция №1.Возбудители гнойно - воспалительных процессов. Стафилококки. Общая характеристика.
Подавляющее большинство гнойно - воспалительных заболеваний вызывают кокки, т.е. имеющие сферическую (шаровидную) форму микроорганизмы. Их делят на две большие группы - грамположительные и грамотрицательные. Внутри этих групп выделяют аэробные и факультативно - анаэробные кокки и анаэробные кокки.
Среди грамположительных аэробных и факультативно - анаэробных кокков наибольшее значение имеют микроорганизмы семейства Micrococcaceae (род Staphylococcus) и семейства Streptococcaceae (род Streptococcus), среди грамотрицательных аэробных и факультативно - анаэробных кокков - представители семейства Neisseriaceae (N.gonorrhoeae - гонококк и N.meningitidis - менингококк). Среди грамположительных анаэробных кокков наибольшее значение имеют пептококки и пептострептококки, среди грамотрицательных анаэробных кокков - вейлонеллы.
Представители семейства Micrococcaceae, способные вызывать заболевания у человека, относятся к родам Staphylococcus, Micrococcus и Stomatococcus.
Род Staphylococcus.
В состав рода входит более 20 видов, из которых наибольшее значение имеют S.aureus (золотистый стафилококк), S.epidermidis, S.saprophyticus.
Морфология. Грамположительные кокки, для которых характерно взаиморасположение скоплениями в виде гроздей винограда, т.к. они делятся во взаимоперпендикулярных плоскостях. Имеют микрокапсулу, спор не образуют, жгутиков не имеют.
Культуральные свойства. Факультативные анаэробы, хемоорганотрофы. Хорошо растут на простых питательных средах, в том числе на средах с 5- 10% NaCl. Температурный оптимум от +35 до +37о С, рН 6-8 (лучше слабощелочная реакция среды). На плотных средах образуют непрозрачные круглые (2-4 мм в диаметре) ровные колонии, окрашенные в цвет липохромного пигмента (кремовый, желтый, оранжевый). Кроме S- форм колоний могут образовывать R- формы. На жидких средах дают равномерное помутнение, затем выпадает рыхлый осадок.
Биохимические свойства. Обладают высокой биохимической активностью, образуют различные ферменты, во многом определяющие патогенность. Каталаза- положительны, оксидаза- отрицательны. Углеводы ферментируют до кислоты без газа, разжижают желатин с образованием воронки, образуют сероводород. По наличию коагулазы их делят на две группы - коагулаза- положительные и коагулаза- отрицательные. Среди патогенных видов коагулаза - положителен лишь S.aureus, остальные - отрицательны.
Антигенная структура очень сложная (более 50 типов антигенов). По специфичности антигены подразделяют на родовые (общие для стафилококков), перекрестнореагирующие (с изоантигенами эритроцитов, кожи и почки человека), видо- и типоспецифические. Видоспецифическими антигенами являются тейхоевые кислоты, белок А золотистого стафилококка. Антигенными свойствами обладают токсины.
Факторы патогенности. К ним относят микрокапсулу, компоненты клеточной стенки (тейхоевые кислоты, белок А), ферменты и токсины.
1. Факторами адгезии являются высокие гидрофобные свойства поверхностных структур.
2. Компоненты клеточной стенки стимулируют развитие воспалительных реакций, основное значение в них имеют нейтрофилы.
3. Разнообразные ферменты стафилококков играют роль факторов агрессии и защиты. Главным фактором является плазмокоагулаза, свертывающая сыворотку (плазму) крови и образующая тромбиноподобное вещество, обвалакивающее стафилококки и препятствующее действию защитных реакций организма. Кроме нее - фибролизин, ДНК- аза, лецитиназа, фосфатаза.
4. Стафилококки синтезируют обширный комплекс экзотоксинов.
Мембраноповреждающие токсины могут повреждать эритроциты (гемолизины), лейкоциты, макрофаги, тромбоциты и др. Выделяют несколько типов, отличающихся по антигенной структуре, спектру лизируемых клеток, скорости действия.
Эксфолиативные токсины оказывают дерматонекротическое действие (пузырчатка новорожденных).
Экзотоксин, вызывающий синдром токсического шока. Высокосорбционные тампоны вызывали тяжелый эндотоксический шок у женщин.
Энтеротоксины, с которыми связаны пищевые интоксикации. Энтеротоксины - термостабильные белки со свойствами суперантигенов. Они вызывают избыточный синтез интерлейкина 2, который и обусловливает интоксикацию. Интоксикации чаще связаны с употреблением инфицированных стафилококками молочных продуктов.
5. Ряд экзотоксинов и других структур стафилококков обладают аллергизирующим действием, обусловливая эффект развития ГЗТ. Наличие перекрестно - реагирующих антигенов способствует развитию аутоиммунных процессов.
6. Факторы, угнетающие фагоцитоз - капсула, белок А, тейхоевые кислоты, пептидогликан, токсины.
Генетика. Для стафилококков характерна высокая изменчивость, связанная с мутациями и рекомбинациями. У них могут быть различные плазмиды. Особенно распространены штаммы с плазмидами устойчивости к различным антибиотикам (прежде всего - к пенициллинам). Выделяют фаготипы и проводят фаготипирование стафилококков. Для типирования используют наборы умеренных фагов.
Особые свойства возбудителя.
1. Способность поражать практически любую ткань и орган.
2. Очень высокая устойчивость среди неспорообразующих бактерий к факторам внешней среды.
3. Постоянное пребывание на кожных покровах и сообщающихся с внешней средой слизистых оболочках.
4. Суперантигенные свойства.
5. Высокая изменчивость и антибиотикорезистентность, что имеет важное значение для эпидемического процесса.
Эпидемиологические особенности. Стафилококковые инфекции могут носить характер эндогенной инфекции (повреждение органов и тканей с проникновением возбудителя) или экзогенный характер, обусловленный различными путями заражения - алиментарным (при стафилококковых отравлениях), контактно - бытовым, воздушно - капельным и воздушно - пылевым.
Существенное значение имеет носительство патогенных стафилококков, чаще всего - на слизистой носа и зева, на втором месте - кожа. В условиях медицинских учреждений (родильные дома, хирургические стационары) и в закрытых коллективах особую опасность представляют резидентные (постоянные) носители, у которых наблюдается колонизация слизистых носа и зева и длительная персистенция стафилококков. Длительное носительство в определенной мере связано с дефицитом местного секреторного иммунитета (секреторного IgA в первую очередь) и хроническими очагами воспаления в организме.
Особенности клиники и патогенеза. Только инфекции, вызываемые золотистым стафилококком, включают более 100 нозологических форм. Стафилококковые инфекции можно разделить на локальные и системные (генерализованные). Среди первых - фурункулы, панариции, мастит, гнойные осложнения раневых поверхностей. Среди вторых - сепсис (септицемия - “гнилокровие” с размножением возбудителя в крови, септикопиемия - сепсис с гнойными очагами - метастазами), стафилококковые пневмонии, осложнения после родов и операций, приводящих к синдрому токсического шока, остеомиелиты и др.
Постинфекционный иммунитет при стафилококковых инфекциях можно разделить на клеточный и гуморальный, антибактериальный и антитоксический. В связи с очень разнообразной и изменчивой антигенной структурой перекрестного иммунитета у стафилококков нет. В защите важную роль имеют антитоксины, антибактериальные антитела, антитела против ферментов патогенности, Т- лимфоциты, фагоциты (тканевые макрофаги при локализованных формах).
Лабораторная диагностика. Применяют бактериологические, микроскопические и серологические методы. Основным является бактериологический метод.
Материал для исследования - кровь, гной, слизь из носа и зева, отделяемое ран, мокрота (при пневмонии), испражнения (при колите), подозрительные продукты, рвотные массы и промывные воды желудка, испражнения - при пищевых интоксикациях.
Из серологических тестов чаще применяют РПГА и ИФА для выявления антител к видоспецифическим антигенам (тейхоевым кислотам).
Для определения энтеротоксигенности чаще применяют биологический метод, определение энтеротоксина в реакции преципитации в агаре, определение РНК- азы (коррелирует с выработкой энтеротоксина). Из видоспецифических антигенов отдельно следует сказать о белке А золотистого стафилококка, имеющего важное значение в патогенезе и лабораторной диагностике. Уникальным свойством белка А является его способность связывать Fc- фрагменты иммуноглобулинов (субклассов 1,2 и 4 IgG). Белок А широко используют в ИФА для выявления специфических IgG- антител (конъюгаты “белок А - пероксидаза”), стафилококковый реагент - для дифференциации титров IgG- и IgM- антител в серологических реакциях по снижению титров IgG- антител после обработки проб сыворотки крови стафилококковым реагентом.
Профилактика и лечение - это наиболее сложные вопросы стафилококковых инфекций. Для проведения адекватной антимикробной терапии необходимо определение чувствительности культур к антибиотикам (прежде всего - к бета- лактамовым), в тяжелых и затяжных случаях применяют донорский антистафилококковый иммуноглобулин. Определенный эффект может оказать фаготерапия. Для создания иммунитета применяют стафилококковый анатоксин, создающий антитоксический иммунитет. Современные вакцины недостаточно эффективны, поскольку иммунитет при стафилококковых инфекциях типоспецифический (к определенному серовару).
Лекция №2. Стрептококки. Гонококки, менингококки.
Стрептококки.
В семейство Streptococcaceae входит семь родов, из которых для человека наибольшее значение имеют стрептококки (род Streptococcus) и энтерококки (род Enterococcus). Наиболее значимые виды - S.pyogenes (стрептококки группы А), S.agalactiae (стрептококки группы В), S.pneumoniae (пневмококк), S.viridans (зеленящие стрептококки, биогруппа mutans), Enterococcus faecalis.
Морфология. Стрептококки (от греч. streptos - цепочка и coccus - зерно) - грамположительные цитохромнегативные бактерии шаровидной или овоидной формы, растущие чаще в виде цепочек, преимущественно неподвижные, не имеют спор. Патогенные виды образуют капсулу (у пневмококка имеет диагностическое значение).
Факультативные (большинство) или строгие анаэробы.
Культуральные свойства. Стрептококки плохо растут на простых питательных средах. Обычно используют среды с кровью или сывороткой крови. Чаще применяют сахарный бульон и кровяной агар, содержащий 5% дефибринированной крови. На бульоне рост придонно - пристеночный в виде крошковатого осадка, бульон чаще прозрачен. На плотных средах чаще образуют очень мелкие колонии. Оптимум температуры +37о С, рН - 7,2-7,6. На плотных средах стрептококки группы А образуют колонии трех типов:
- мукоидные (напоминают капельку воды) - характерны для вирулентных штаммов, имеющих капсулу;
- шероховатые - плоские, с неровной поверхностью и фестончатыми краями - характерны для вирулентных штаммов, имеющих М- антигены; - гладкие - характерны для маловирулентных штаммов.
Предпочитают газовую смесь с 5% СО2. Способны образовывать L- формы.
Для дифференциации стрептококков используют различные признаки: рост при +10о и 45о С, рост на среде с 6,5% NaCl, рост на среде с рН 9,6, рост на среде с 40% желчи, рост в молоке с 0,1% метиленовым синим, рост после прогревания в течение 30 мин. при 60о С. Наиболее распространенный S.pyogenes относится к 1 группе (все признаки отрицательны), энтерококки (3 группа) - все признаки положительны.
Существует ряд классификаций стрептококков. Наиболее проста классификация, основанная на особенностях роста этих микроорганизмов на агаре с кровью барана (по отношению к эритроцитам).
Бета - гемолитические стрептококки при росте на кровяном агаре образуют вокруг колонии четкую зону гемолиза, альфа - гемолитические - частичный гемолиз и позеленение среды (превращение окси- в метгемоглобин), гамма- гемолитические - на кровяном агаре гемолиза незаметно. Альфа - гемолитические стрептококки за зеленый цвет среды называют S.viridans (зеленящими).
Антигенная структура. Серологическая классификация имеет практическое значение для дифференциации имеющих сложное антигенное строение стрептококков. В основе классификации - группоспецифические полисахаридные антигены клеточной стенки. Выделяют 20 серогрупп, обозначенных заглавными латинскими буквами. Наибольшее значение имеют стрептококки серогрупп А,В и D.
У стрептококков серогруппы А имеются типоспецифические антигены - белки М, Т и R. По М- антигену гемолитические стрептококки серогруппы А подразделены на серовары (около 100).
Стрептококки имеют перекрестно - реагирующие антигены с антигенами клеток базального слоя эпителия кожи, эпителиальных клеток корковой и медуллярной зон тимуса. В клеточной стенке стрептококков обнаружен также антиген (рецептор II), способный взаимодействовать с Fс- фрагментом IgG.
Факторы патогенности стрептококков.
1. Белок М- главный фактор. Определяет адгезивные свойства, угнетает фагоцитоз, определяет типоспецифичность, обладает свойствами суперантигена. Антитела к М- белку обладают протективными свойствами.
2. Капсула - маскирует стрептококки за счет гиалуроновой кислоты, аналогичной гиалуроновой кислоте в тканях хозяина.
3. С5а - пептидаза - расщепляет С5а - компонент комплемента, чем снижает хемоатрактивную активность фагоцитов.
4. Стрептококки вызывают выраженную воспалительную реакцию, в значительной степени обусловленную секрецией более 20 растворимых факторов - ферментов (стрептолизины S и О, гиалуронидаза, ДНК- азы, стрептокиназа, протеазы) и эритрогенных токсинов.
Эритрогенин - скарлатинозный токсин, обусловливающий за счет иммунных механизмов образование ярко красной скарлатинозной сыпи. Выделяют три серологических типа этого токсина (А,В и С). Токсин обладает пирогенным, аллергенным, иммуносупрессивным и митогенным действием.
Генетика. Мутации и рекомбинации менее выражены, чем у стафилококков. Способны синтезировать бактериоцины. Фаги для дифференциации не применяют.
Эпидемиологические особенности. Основными источниками являются больные острыми стрептококковыми инфекциями (ангина, пневмония, скарлатина), а также реконвалесценты. Механизм заражения - воздушно - капельный, реже - контактный, очень редко - алиментарный.
Клинико - патогенетические особенности. Стрептококки - обитатели слизистых верхних дыхательных путей, пищеварительного и моче - полового трактов, вызывают различные заболевания эндо- и экзогенного характера. Выделяют локальные (тонзиллит, кариес, ангины, отиты и др.) и генерализованные инфекции (ревматизм, рожистое воспаление, скарлатина, сепсис, пневмония, стрептодермии и др.). Развитие тех или иных форм зависит от ряда условий, в т.ч. от входных ворот, различных факторов патогенности, состояния иммунной системы (особую роль играют антитоксины и типоспецифические М- антитела).
Особое положение в роде Streptococcus занимает вид S.pneumoniae (пневмококк) - этиологический агент крупозной пневмонии, острых и хронических воспалительных заболеваний легких. От остальных стрептококков отличается морфологией (чаще диплококки в форме пламени свечи, плоскими концами друг к другу, обладают выраженной капсулой), антигенной специфичностью (имеют 83 серовара по капсульному полисахаридному антигену), высокой чувствительностью к желчи и оптохину, вызывают альфа - гемолиз. Главный фактор патогенности - полисахаридная капсула.
Скарлатину вызывают различные серотипы бета - гемолитических стрептококков, обладающих М- антигеном и продуцирующих эритрогенин (токсигенные стрептококки серогруппы А). При отсутствии антитоксического иммунитета возникает скарлатина, при наличии - ангина.
Лабораторная диагностика. Основной метод диагностики - бактериологический. Материал для исследования - кровь, гной, слизь из зева, налет с миндалин, отделяемое ран. Решающим при исследовании выделенных культур является определение серогруппы (вида). Группоспецифические антигены определяют в реакции преципитации, латекс - агглютинации, коагглютинации, ИФА и в МФА с моноклональными антителами (МКА). Серологические методы чаще используют для диагностики ревматизма и гломерулонефрита стрептококковой этиологии - определяют антитела к стрептолизину О и стрептодорназе.
Грамотрицательные аэробные и факультативно - анаэробные кокки.
Наибольшее значение имеет род Neisseria, из видов - N.gonorrhoeae - гонококк и N.meningitidis - менингококк, остальные виды нейссерий - комменсалы.
Морфология. Нейссерии - диплококки, напоминающие кофейные зерна или бобы, прилегающие друг к другу уплощенными сторонами. Для них характерно наличие капсулы, а также пилей и ворсинок, облегчающих адгезию патогенных нейссерий к эпителию.
Культуральные особенности. Для культивирования патогенные нейссерии требуют среды с кровью, сывороткой крови или асцитической жидкостью человека. Каждый вид избирательно ферментирует углеводы. Оптимальная температура +37о С, рН- 7,2-7,4. Для культивирования пригодны кровяной и шоколадный агар с добавлением крахмала, нужна повышенная концентрация CO2, селективные компоненты, подавляющие рост сопутствующей микрофлоры. N.gonorrheae (гонококк).
Гонококк - возбудитель гонореи - венерического заболевания с воспалительными проявлениями в моче- половых путях. Субстрат для колонизации - эпителий уретры, прямой кишки, конъюнктивы глаза, глотки, шейки матки, маточных труб и яичника.
Диплококки, хорошо окрашиваемые метиленовым синим и другими анилиновыми красителями, плеоморфные (полиморфизм). Очень прихотливы к условиям культивирования и питательным средам. Из углеводов ферментируют только глюкозу.
Антигенная структура очень изменчива - характерны фазовые вариации (исчезновение антигенных детерминант) и антигенные вариации (изменение антигенных детерминант). Основную антигенную нагрузку несут детерминанты пилей и поверхностных белков. С высокой антигенной изменчивостью связано отсутствие иммунной защиты против повторного заражения. Наибольшее антигенное родство - с менингококками.
Факторы патогенности. Основными факторами являются пили, с помощью которых гонококки осуществляют адгезию и колонизацию эпителиальных клеток слизистой оболочки моче- половых путей, и липополисахарид (эндотоксин, освобождающийся при разрушении гонококков). Гонококки синтезируют IgAI- протеазу, расщепляющую IgA.
Генетика. Характерна генетическая изменчивость, даже на протяжении жизни одной микробной популяции. Передача информации осуществляется преимущественно конъюгацией. Выявлены F- и R- плазмиды, в т.ч. плазмиды, несущие ген бета - лактамазы.
Лабораторная диагностика. Бактериоскопическая диагностика включает окраску по Граму и метиленовым синим. Типичные признаки гонококка - грамотрицательная окраска, бобовидные диплококки, внутриклеточная локализация. При антибиотикотерапии, хронической гонорее и некоторых других случаях как морфологические признаки, так и отношение к окраске по Граму может изменяться. Более достоверна люминесцентная диагностика с использованием прямого и непрямого иммунофлюоресцентного анализа.
Посев производят на специальные среды (КДС- МПА из мяса кролика или бычьего сердца с сывороткой, асцит- агар, кровяной агар). Характерные признаки гонококка при бактериологической диагностике - грамотрицательные диплококки, колонии которых на плотных средах обладают оксидазной активностью. Ферментируют глюкозу, но не мальтозу или сахарозу. N.meningitidis (менингококки).
Менингококк - возбудитель менингококковой инфекции - строгого антропоноза с воздушно - капельной передачей возбудителя. Основной источник - носители. Природный резервуар - носоглотка человека. Морфологические, культуральные и биохимические свойства аналогичны гонококку. Отличия - ферментируют не только глюкозу, но и мальтозу, продуцируют гемолизин. Обладают капсулой, имеющей большие размеры и другое строение, чем у гонококка.
Антигенный состав. Имеют четыре основные антигенные системы.
1. Капсульные группоспецифические полисахаридные антигены. Штаммы серогруппы А наиболее часто вызывают эпидемические вспышки.
2. Белковые антигены наружной мембраны. По этим антигенам менингококки серогрупп В и С подразделены на классы и серотипы.
3. Родо- и видоспецифические антигены.
4. Липополисахаридные антигены (8 типов). Имеют высокую токсичность, вызывают пирогенное действие.
Факторы патогенности. Факторы адгезии и колонизация - пили и белки наружной мембраны. Факторы инвазивности - гиалуронидаза и другие продуцируемые ферменты (нейраминидаза, протеазы, фибринолизин). Большое значение имеют капсульные полисахаридные антигены, защищающие микроорганизмы от фагоцитоза.
Иммунитет стойкий, антимикробный.
Лабораторная диагностика основана на бактериоскопии, выделении культуры и ее биохимической идентификации, серологических методах диагностики. Посев материала производят на твердые и полужидкие питательные среды, содержащие кровь, асцитическую жидкость, сыворотку крови.
Оксидаза- позитивные культуры рассматривают как принадлежащие к роду Neisseria. Для менингококка характерна ферментация глюкозы и мальтозы. Принадлежность к серогруппе определяют в реакции агглютинации (РА). Для обнаружения антигенов может также применяться реакция коагглютинации, латекс- агглютинации, ИФА, для серодиагностики - РНГА с группоспецифическими полисахаридными антигенами.
Лекция №3. Клостридии.
Общая характеристика патогенных анаэробных бактерий.
Облигатные анаэробы - микроорганизмы, живущие только в условиях крайне низкого содержания кислорода - в почве, иле водоемов, кишечниках позвоночных и человека. У теплокровных анаэробы составляют основную массу нормальной кишечной микрофлоры и определяют ряд важнейших функций организма.
Разнородную группу анаэробных грамположительных бактерий дифференцируют прежде всего по способности к спорообразованию и морфологическим особенностям. В патологии человека наибольшее значение имеют анаэробные спорообразующие бактерии рода Clostridium. Среди неспорообразующих грамположительных анаэробов медицинское значение имеют семейство лактобацилл (Lactobacillaceae) и род бифидобактерий (Bifidibacterium), в патологии - неспорообразующие грамотрицательные анаэробы родов Bacteroides (бактероиды) и Fusobacterium (фузобактерии).
Род Clostridium.
Это подвижные крупные палочки (большинство видов), образуют овальные или круглые эндоспоры, придающие клостридиям (греч. kloster - веретено) ветеренообразную форму. Спора в диаметре больше диаметра (поперечника) вегетативной клетки. Грамположительны, хемоорганотрофы. Строгие анаэробы.
По экологическим и патогенным свойствам можно выделить три группы клостридий: - сапрофиты, вызывающие бродильные (сахаролитические) процессы;
- сапрофиты, вызывающие процессы гниения (протеолиза);
- патогенные виды - по биохимическим свойствам могут вызывать процессы гниения и брожения. Патогенные виды клостридий условно разделить на три группы - возбудителей травматических (раневых) клостридиозов - газовой гангрены, столбняка, возбудителей энтеральных клостридиозов (токсикоинфекций) - ботулизма, псевдомембранозного колита и виды, вызывающие патологические процессы только в ассоциациях между собой или с другими микроорганизмами.
Современная систематика выделяет пять групп клостридий по расположению спор, способности гидролизовать желатин и наличию особых условий для роста. Все патогенные для человека виды ферментируют желатин, отличаясь расположением спор - терминальное (в виде тенисных ракеток) у четвертой группы (C.tetani - возбудитель столбняка), субтерминальное (веретено) у второй группы (остальные возбудители: C.botulinum- возбудитель ботулизма, C.perfringens и другие возбудители газовой гангрены, C.difficile - возбудитель псевдомембранозного колита).
Микробиология газовой гангрены.
Газовая гангрена - анаэробная поликлостридиальная (т.е. вызываемая различными видами клостридий) раневая (травматическая) инфекция. Основное значение имеет C.perfringens, реже - C.novyi, а также другие виды клостридий в стойких ассоциациях между собой, аэробными гноеродными кокками и гнилостными анаэробными бактериями.
C.perfringens - нормальный обитатель кишечников человека и животных, в почву попадает с испражнениями. Является возбудителем раневой инфекции - вызывает заболевание при попадании возбудителя в анаэробных условиях в раны. Обладает высокой инвазивностью и токсигенностью. Инвазивность связана с выработкой гиалуронидазы и других ферментов, оказывающих разрушающее действие на мышечную и соединительную ткани. Главный фактор патогенности - экзотоксин, оказывающий гемо-, некро-, нейро-, лейкотоксическое и летальное воздействие. В соответствии с антигенной специфичностью экзотоксинов выделяют серотипы возбудителя. Наряду с газовой гангреной C.perfringens вызывает пищевые токсикоинфекции (в их основе - действие энтеротоксинов и некротоксинов).
Особенности патогенеза. В отличии от гнойных заболеваний, вызываемых аэробами, при анаэробной инфекции преобладает не воспаление, а некроз, отек, газообразование в тканях, отравление токсинами и продуктами распада тканей. Клостридии - некропаразиты, активно создающие анаэробные условия и вызывающие некроз тканей, т.е. условия для своего размножения. Некроз мышечной и соединительной тканей - следствие некротоксического действия токсинов и ферментов, газообразование в тканях - результат ферментативной активности клостридий. Общее действие токсинов (общая интоксикация) проявляется преимущественно в нейротоксическом воздействии.
Иммунитет - преимущественно антитоксический.
Лабораторная диагностика включает бактериоскопию отделяемого ран, выделение и идентификацию возбудителя, выявление и идентификацию токсина в биопробах с использованием реакции нейтрализации специфическими антитоксическими антителами.
Особенностью всех спорообразующих бактерий является устойчивость к нагреванию, что учитывается при лабораторной диагностике - исследуют прогретую и непрогретую пробы. При исследовании на клостридии одну (непрогретую) пробу засевают на жидкие среды (казеиновые или мясные) и плотные дифференциально - диагностические среды (Вильсон - Блера, Виллиса - Хоббса). Другие части исходного материала прогревают при +80 и +100о С в течение до 20 минут и засевают в жидкие накопительные среды. Выросшие культуры грамположительных палочек пересевают на плотные дифференциально - диагностические среды.
Среда Виллиса - Хоббса содержит кроме питательного агара, лактозу, индикатор, яичный желток и обезжиренное молоко. Дифференциация видов клостридий осуществляется по изменению цвета индикатора в красный (ферментация лактозы) и наличию вокруг колоний зоны (ореола) опалесценции (лецитиназная активность).
Колонии C.perfringens окрашены в цвет индикатора и имеют ореол опалесценции.
Профилактика и лечение. В основе предупреждения газовой гангрены - своевременная и правильная хирургическая обработка ран. При тяжелых ранениях вводят антитоксические сыворотки против основных видов клостридий по 10 тысяч МЕ, в лечебных целях - по 50 тысяч МЕ. Clostridium difficile.
Это представитель нормальной микрофлоры кишечника, редко вызывает газовую гангрену, является возбудителем псевдомембранозного энтероколита - результата нерациональной антибиотикотерапии и развития дисбактериоза, обусловленного доминированием этого возбудителя. При применении антибиотиков (особенно клиндомицина, ампициллина и цефалоспоринов) и цитостатиков возникает глубокий дисбаланс микрофлоры кишечника и колонизация его C.difficile. Токсический компонент патогенности этого вида клостридий состоит из энтеротоксина и цитотоксина. Возбудитель проявляет высокую резистентность к антибиотикам широкого спектра, что способствует массированной колонизации кишечника и секреции большого количества токсинов, вызывающих изменения кишечной стенки. Псевдомембранозный колит - госпитальная инфекция, доминирующая среди кишечных поражений, однако бактериологически трудно выявляемая (сложности выделения и изучения анаэробов).
Микробиология столбняка.
Столбняк - острая раневая инфекция, характеризующаяся поражением нейротоксином двигательных клеток спинного и головного мозга, которое проявляется в виде судорог поперечно - полосатой мускулатуры. Болеют люди и сельскохозяйственные животные. Почва, особенно загрязненная испражнениями человека и животных, является постоянным источником заражения столбняком.
Возбудитель - C.tetani - крупная спорообразующая грамположительная палочка. Споры располагаются терминально (вид барабанной палочки), подвижна за счет жгутиков - перитрихов. Обязательный анаэроб. Споры обладают очень высокой устойчивостью.
Антигенные свойства. Возбудитель имеет О- и Н- антигены.
Факторы патогенности. Главный фактор - сильнейший экзотоксин. Выделяют две его основные фракции - тетаноспазмин (нейротоксин) и тетанолизин (гемолизин). Нейротоксин в центральную нервную систему проникает в области мионевральных синапсов, передается от нейрона к нейрону в области синапсов, накапливается в двигательных зонах спинного и головного могза, блокирует синаптическую передачу. Смерть наступает от паралича дыхательного центра, асфиксии (поражение мышц гортани, диафрагмы, межреберных мышц) или паралича сердца.
Лабораторная диагностика. Микробиологическая диагностика включает бактериоскопию исходных материалов, посев для выделения возбудителя и его идентификацию, обнаружение столбнячного токсина.
Исследованию подлежит материал от больного (особенно в местах проникновения возбудителя в организм - из ран), трупа (кровь, кусочки печени и селезенки), перевязочный и шовный хирургический материал, пробы почвы, пыли и воздуха.
Выделение возбудителя проводят по стандартной для анаэробов схеме, используя различные плотные и жидкие (среда Китта - Тароцци) среды, идентификацию - на основе морфологических, культуральных, биохимических и токсигенных свойств.
Наиболее простой и эффективный метод микробиологической диагностики - биопроба на белых мышах. Одну группу заражают исследуемым материалом, вторую (контрольную) - после смешивания проб с антитоксической столбнячной сывороткой. При наличии столбнячного токсина опытная группа мышей погибает, контрольная - остается живой.
Лечение и экстренная профилактика. Используют донорский противостолбнячный иммуноглобулин (антитоксин), антитоксическую сыворотку (350МЕ/кг), антибиотики (пенициллины, цефалоспорины). Для создания вакцинального иммунитета используют столбнячный анатоксин, чаще в составе АКДС вакцины (анатоксины столбняка, дифтерии и убитые коклюшные палочки).
Микробиология ботулизма.
Ботулизм - тяжелая пищевая токсикоинфекция, связанная с употреблением продуктов, зараженных C.botulinum, и характеризующаяся специфическим поражением центральной нервной системы. Свое название получила от лат. botulus - колбаса.
Свойства возбудителя. Крупные полиморфные грамположительные палочки, подвижные, имеют перитрихиальные жгутики. Споры овальные, располагаются субтерминально (тенисная ракетка). Образуют восемь типов токсинов, отличающихся по антигенной специфичности, и соответственно выделяют 8 типов возбудителя. Среди важнейших характеристик - наличие или отсутствие протеолитических свойств (гидролиз казеина, продукция сероводорода).
Токсин оказывает нейротоксическое действие. Токсин попадает в организм с пищей, хотя вероятно может накапливаться при размножении возбудителя в тканях организма. Токсин термолабильный, хотя для полной инактивации необходимо кипячение до 20 мин. Токсин быстро всасывается в желудочно - кишечном тракте, проникает в кровь, избирательно действует на ядра продолговатого мозга и ганглиозные клетки спинного мозга. Развиваются нервно - паралитические явления - нарушения глотания, афония, дисфагия, офтальмо - плегический синдром (косоглазие, двоение в глазах, опущение век), параличи и парезы глоточных и гортанных мышц, остановка дыхания и сердечной деятельности.
Лабораторная диагностика. Принципы - общие для клостридий. Выделяют и идентифицируют возбудитель, однако наибольшее практическое значение имеет обнаружение ботулотоксина и определение его серотипа.
Для выделения возбудителя пробы сеют на плотные среды и накопительную среду Китта - Тароцци (часть материала предварительно прогревают при +85о С).
Для изучения токсина проводят биопробы на белых мышах (одна группа - опытная и четыре контрольные со смесью материала и соответствующей антисыворотки - типа А,В,С и Е). Погибают все партии, кроме одной (с гомологичной типу токсина антисывороткой). Можно также определять токсин в РНГА с антительным диагностикумом.
Лечение и профилактика. В основе - раннее применение антитоксических сывороток (поливалентных или при установлении типа - гомологичных). В основе профилактики - санитарно - гигиенический режим при обработке пищевых продуктов. Особенно опасны грибные консервы домашнего приготовления и другие продукты, хранящиеся в анаэробных условиях.
Лекция №4. Коринебактерии. Бордетеллы.
Род Corynebacterium.
Возбудитель дифтерии - Corynebacterium diphtheriae и большая группа близких по морфологическим и биохимическим свойствам микроорганизмов рода коринебактерий называют коринеформными бактериями или дифтероидами.Они представлены грамположительными неподвижными палочками, чаще с утолщениями на концах, напоминающими булаву (coryne - булава). Дифтероиды широко распространены в почве, воздухе, пищевых продуктах (молоке). Среди них можно выделить три экологические группы:
- патогены человека и животных;
- патогены растений;
- непатогенные коринебактерии.
Многие виды дифтероидов являются нормальными обитателями кожи, слизистых зева, носоглотки, глаз, дыхательных путей, уретры и половых органов.
Дифтерия.
Дифтерия - острое инфекционное заболевание преимущественно детского возраста, которое характеризуется интоксикацией организма дифтерийным токсином и характерным фибринозным (дифтеритическим) воспалением в месте локализации возбудителя (phther - пленка).
Морфологические и тинкториальные свойства. C.diphtheriae - тонкие полиморфные палочки с булавовидными концами, часто содержащие волютиновые включения, выявляемые окраской метиленовым синим или по Нейссеру. При последнем палочки окрашены в желто - соломенный цвет, зерна волютина (полиметафосфата) - в темно - коричневый цвет. В культурах палочки находятся под углом друг к другу (особенности деления) , образуя различные фигуры - растопыренных пальцев, V, Y, L и т.д. Имеют микрокапсулу, а также фимбрии, облегчающие адгезию к эпителию слизистых оболочек.
Культуральные свойства. На простых средах коренебактерии дифтерии не растут. Они требуют сред с кровью или сывороткой крови (среды Леффлера, Ру), на которых рост отмечается уже через 10-12 часов, за это время сопутствующая (контаминирующая пробы) микрофлора полностью развиться не успевает.
Наиболее оптимальны теллуритовая среда и теллурит - шоколадный агар Маклеода. Высокие концентрации теллурита калия в этих средах подавляют рост посторонней флоры. Коринебактерии дифтерии восстанавливают теллурит до металлического теллура, что придает их колониям темно - серый или черный цвет.
У этого возбудителя выделяют биотипы - gravis, mitis, intermedius, отличающиеся по морфологии, антигенным и биохимическим свойствам, тяжести заболеваний у человека. Тип gravis чаще вызывает вспышки и более тяжелое течение, для него характерны крупные с неровными краями и радиальной исчерченностью колонии в виде маргаритки (R- формы). Тип mitis вызывает преимущественно легкие спорадические заболевания, образует на плотных средах мелкие гладкие колонии с ровными краями (S- формы). Тип intermedius занимает промежуточное положение, образует на плотных средах переходные по характеристикам RS- формы, однако еще более мелкие. На жидких средах вызывают помутнение сред, образуют крошковидный осадок.
Биохимические свойства. Коринебактерии дифтерии ферментируют глюкозу, мальтозу. Отсутствие активности в отношении сахарозы и мочевины - важный дифференциальный признак среди дифтероидов. Обладают цистеназной активностью (расщепляют цистеин) - проба Пизу.
Антигенная структура. Выделяют О- и К- антигены. Полисахаридные компоненты О- антигенов клеточной стенки обладают межродовыми свойствами, обусловливая неспецифические перекрестные реакции с микобактериями, актиномицетами (нокардиями).
Поверхностные К- антигены - капсульные белки, обладают видовой специфичностью и иммуногенностью. Выделяют 11 серотипов. Серотипы 1-5 и 7 относятся к биовару gravis. Серотипирование культур проводят в РА с диагностическими сыворотками к соответствующим сероварам и полигрупповой агглютинирующей сывороткой.
В серологической диагностике у людей чаще применяют РПГА, более чувствительную, чем РА. В настоящее время применяют также ИФА. Многие штаммы коринебактерий дифтерии (особенно нетоксигенные) обладают спонтанной агглютинабельностью и полиагглютинабельностью.
Факторы патогенности. Токсигенные штаммы возбудителя дифтерии продуцируют сильный экзотоксин (термолабильный высокотоксичный иммуногенный белок). Нетоксигенные штаммы не вызывают заболевания.
Токсин вызывает необратимое блокирование удлинения полипептидной цепи, т.е. любого белкового синтеза. Поражаются в основном определенные системы: симпатико - адреналовая, сердце и кровеносные сосуды, периферические нервы. Отмечаются структурные и функциональные нарушения миокарда, демиелинизация нервных волокон, приводящая к параличам и парезам.
Способность к токсинообразованию проявляют лишь лизогенные штаммы, инфицированные бактериофагом (бета - фагом), несущим ген tox, который кодирует структуру токсина (т.е. несущие гены умеренного профага в своей хромосоме). Фаготипирование применяют для дифференциации штаммов коринебактерий дифтерии.
Эпидемиология. Резервуар - человек (больной, реконвалесцент, бактерионоситель). Основной путь передачи - воздушно - капельный, сезонность - осенне - зимняя. Возбудитель хорошо сохраняется при низких температурах, высушенном состоянии (слюна, слизь, пыль).
Клинико - патогенетические особенности. Возбудитель в месте внедрения вызывает фибринозное воспаление с образованием плотно спаянной с тканями фибринозной пленки. Существенное значение в вызываемой патологии имеет действие экзотоксина (описано в разделе “факторы патогенности” ). По локализации выделяют дифтерию ротоглотки (наиболее часто), дыхательных путей, носа и редкой локализации (глаза, наружные половые органы, кожа, раневая поверхность). Дифтерия зева может быть причиной крупа и асфиксии.
Лабораторная диагностика. Основной метод диагностики - бактериологический. Применяют для выявления больных, бактерионосителей, контактных. Стерильными тампонами берут материал для микроскопии и посевов - слизь из зева и носа, пленки с миндалин и других мест, подозрительных на наличие дифтеритических поражений.
Возбудитель выделяют посевом на элективные теллуритовые среды и кровяной агар. На слизистой оболочке глаза часто выявляют C.xerosis (возможная причина хронических конъюнктивитов), в носоглотке - C.pseudodiphtheriticum (палочка Хофманна), выявляют и другие дифтероиды.
Для дифференциации возбудителя дифтерии от дифтероидов используют такие показатели, как способность восстанавливать теллурит и образовывать темные колонии, пробу Пизу, ферментацию углеводов (глюкоза, мальтоза, сахароза) и мочевины, способность расти в анаэробных условиях (характерно для возбудителя дифтерии).
Обязательным этапом является определение токсигенности культуры. Наиболее распространенные методы - биопробы на морских свинках, реакция преципитации в агаре. Используют также ИФА с антитоксином, генетические зонды и ПЦР для выявления фрагмента А гена tox.
Лечение. Используют антитоксическую противодифтерийную сыворотку, антибиотики и сульфаниламидные препараты.
Постинфекционный иммунитет - стойкий, преимущественно антитоксический. Для количественного определения уровня антитоксического иммунитета ранее применялась проба Шика (внутрикожное введение токсина), сейчас - РПГА с эритроцитарным диагностикумом, получаемым сенсибилизацией эритроцитов дифтерийным анатоксином.
Профилактика. В основе - массовая иммунизация населения. Используют различные препараты, содержащие дифтерийный анатоксин - АКДС, АДС, АДС- М, АД и АД- М.
Род Bordetella.
Бордетеллы - мелкие коккобациллы, имеют форму овоидной палочки, грамотрицательны, неустойчивы во внешней среде. Основное значение имеют три вида.
1. B.pertussis - возбудитель коклюша - острого инфекционного заболевания, сопровождающегося воспалением гортани, трахеи и бронхов и приступообразным кашлем.
2. B.parapertussis - возбудитель паракоклюша.
3. B.bronchiseptica - возбудитель коклюшеподобного заболевания собак, кошек и кроликов, может вызывать у людей респираторные заболевания по типу ОРВИ (относительно редко).
Культуральные особенности. Бордетеллы требовательны к средам, особенно возбудитель коклюша. Для первичной изоляции этих гемофильных микроорганизмов используют картофельно - глицериновый агар с добавлением крови (среду Борде - Жангу) или казеиново - угольный агар (КУА). На кровяных средах возбудитель коклюша вызывает образование мелких колоний металлического оттенка с потемнением сред, на КУА - буровато - коричневое окрашивание колоний. Основные формы колоний - гладкие (S) - так называемая Iфаза (вирулентные культуры) через промежуточные формы переходят в шероховатые (R) - фаза IV, что сопровождается изменением культуральных, биохимических и антигенных свойств, потерей вирулентности.
Антигенный состав. Имеются общие (родовые) и специфические (видовые) антигены. Видоспецифическими являются соматические О- антигены (агглютиногены), выявляемые в РА.
Факторы патогенности. Главный фактор - термолабильный экзотоксин белковой природы, обладающий тропизмом к нервной и сосудистой системе. Имеется термостабильный эндотоксин, обладающий токсическими и сенсибилизирующими свойствами. Трахеальный цитотоксин вызывает повреждение мерцательного эпителия.
Бактерии продуцируют гиалуронидазу, лецитиназу, плазмокоагулазу.
Эпидемиология. Источник инфекции при коклюше и паракоклюше - больные типичными и стертыми формами инфекции. Механизм заражения - воздушно - капельный. Возбудитель, попавший на слизистую оболочку дыхательных путей, размножается, выделяет экзо- и эндотоксины, некротизирует слизистую, раздражают кашлевые рецепторы и кашлевой центр продолговатого мозга, вызывает спазматические приступы кашля.
Лабораторная диагностика. Посев проводят методом “кашлевых пластинок” или материал берут заднеглоточным тампоном с посевом на агар Борде - Жангу и КУА. Идентификация выделенных культур проводится по морфологическим и культуральным свойствам, а также в РА со специфическими сыворотками. Для экспресс - диагностики и идентификации применяют метод флюоресцирующих антител.
Для серологической диагностики используют различные методы - РА, РСК, РПГА с исследованием парных сывороток, взятых в динамике инфекционного процесса.
Лечение. В основе лечения - антибактериальная терапия антибиотиками (гентамицин, ампициллин и др.).
Иммунитет и иммунопрофилактика. Постинфекционный иммунитет стойкий, видоспецифический, обусловлен гуморальными и клеточными факторами.
Для вакцинации используют убитые бордетеллы I фазы. В вакцине АКДС имеется коклюшный компонент (20 млрд микробных тел/ мл).
Лекция №5. Микобактерии.
Род Mycobacterium.
Микобактерии - кислотоустойчивые неподвижные грамположительные палочковидные (прямые или изогнутые) бактерии, способные образовывать нитевидные и мицелиальные структуры. Для них характерно высокое содержание липидов и восков в клеточных стенках, что обеспечивает устойчивость к спиртам, кислотам, щелочам, дезинфицирующим средствам, высушиванию и действию солнечных лучей, плохую окрашиваемость красителями, высокую гидрофобность, патогенность.
Наряду с кислотоустойчивостью, важной характеристикой микобактерий является медленный рост на питательных средах, особенно микобактерий туберкулеза. Еще одна особенность микобактерий - образование пигментов, часть видов образует пигмент в темноте.
Род микобактерий может насчитывать до 200 паразитических и сапрофитических видов, из них хорошо изучено и идентифицировано около 30 видов. Микобактерии широко распространены в почве и воде, их выявляют у широкого круга тепло- и холоднокровных животных.
Среди патогенных микобактерий наибольшее значение имеют основной возбудитель туберкулеза человека - M.tuberculosis (палочка Коха), M.bovis - возбудитель туберкулеза крупного рогатого скота и M.leprae - возбудитель проказы (лепры). Заболевания у людей могут вызывать также M.avium - возбудитель туберкулеза птиц и около 20 других потенциально патогенных видов, способных вызывать у человека атипичные формы поражений (микобактериозы).
Mycobacterium tuberculosis (палочка Коха).
Морфологические свойства типичны для микобактерий. Это тонкие прямые или слегка изогнутые палочки с зернистыми образованиями в цитоплазме, могут встречаться кокковидные структуры, L - формы. Кислотоустойчивы (высокое содержание липидов и миколовой кислоты в клеточной стенке). Имеют кислотолабильные гранулы (зерна Муха) в цитоплазме. Грамположительны, плохо красятся анилиновыми красителями, по Цилю - Нильсену они окрашиваются в ярко - красный цвет.
Культуральные свойства. Растут в аэробных и факультативно - анаэробных условиях. Растут очень медленно - в течении нескольких недель. Микобактерии нуждаются в белке и глицерине, факторах роста. Наиболее часто используют плотные яичные среды Левенштайна - Йенсена, Финна II, синтетические и полусинтетические жидкие среды.
На плотных средах рост отмечается на 15-40 сутки в виде сухого морщинистого налета кремового цвета (R- формы), колонии по виду напоминают цветную капусту. На жидких средах отмечается рост в виде поверхностной пленки.
Палочка Коха устойчива во внешней среде, в высохших биосубстратах сохраняется до нескольких недель.
Факторы патогенности. Патогенные свойства туберкулезной палочки и биологические реакции, которыми отвечает макроорганизм на внедрение возбудителя, связано с особенностями его химического состава, высоким содержанием липидов и их составом (наличие жирных кислот - фтиоидной, миколовой, туберкулостеариновой и др., фосфатидов и других фракций).
Главный фактор - токсичный гликолипид - “корд - фактор”, легко выявляемый при культивировании на жидких средах. Он обеспечивает сближенное расположение микобактерий в виде кос, жгута, корда. Корд - фактор оказывает токсическое действие на ткани, а также блокирует окислительное фосфорилирование в митохондриях макрофагов (защищает от фагоцитоза). С химическим составом микобактерий связаны еще две их важнейшие характеристики:
- незавершенный фагоцитоз этого внутриклеточного паразита (механизмы - блокада фагосомо - лизосомального слияния, устойчивость к действию лизосомальных ферментов);
- способность вызывать выраженную реакцию ГЗТ, выявляемую с помощью туберкулиновой пробы - “ГЗТ туберкулинового типа”.
Антигенная структура. Микобактерии туберкулеза имеют сложный и мозаичный набор антигенов. В антигенном отношении M.tuberculosis имеет наибольшее сходство с M.bovis и M.microti. Имеются перекрестно - реагирующие антигены с коринебактериями, актимомицетами. Для идентификации микобактерий антигенные свойства практически не используют.
Эпидемиология. Основными путями заражения являются воздушно - капельный и воздушно - пылевой. Основным источником заражения является больной туберкулезом человек. Особую роль имеет скученность проживания, в России наибольшую значимость имеют места заключения, лагеря беженцев, лица без определенного места жительства и другиен социально ущербные группы населения. В относительно небольшом проценте случаев туберкулез обусловлен заражением от животных (чаще - через молоко) М.bovis.
Патогенетические особенности.
В течение жизни человек неоднократно контактирует с микобактериями туберкулеза, однако туберкулезный патологический процесс развивается далеко не у всех инфицированнных. Это зависит от многих факторов и прежде всего - резистентности организма.
Наиболее часто заражение происходит через дыхательные пути. Попавшие в организм микобактерии захватываются альвеолярными и легочными макрофагами. В месте попадания может развиться первичный аффект (бронхопневмонический фокус). Далее возбудитель транспортируется в регионарные лимфоузлы, вызывая воспалительную реакцию - лимфангоит и лимфаденит. Первичный аффект, лимфангоит и лимфаденит - первичный комплекс (первичный очаг туберкулеза), характеризующийся образованием по ходу лимфатических путей и узлов гранулем в виде бугорков (бугорчатка или туберкулез).
Образование гранулем представляет собой клеточную реакцию ГЗТ на ряд химических компонентов микобактерий. В центре гранулемы в очаге некроза (казеозного распада) находятся микобактерии. Очаг окружен гигантскими многоядерными клетками Пирогова - Лангханса, их окружают эпителиоидные клетки а по периферии - лимфоциты, плазматические и мононуклеарные клетки.
Исходы первичного очага:
- при достаточной резистентности организма размножение возбудителя в гранулемах прекращается, очаг окружается соединительнотканной капсулой и обезизвествляется (откладываются соли кальция). Этот процесс определяется формированием нестерильного инфекционного иммунитета к возбудителю туберкулеза. Нестерильность - способность микобактерий длительно сохраняться в первичном очаге и ждать свой час (иногда через несколько десятилетий);
- при недостаточной резистентности - усиленный казеозный распад очага, казеозная пневмония, тяжелая первичная легочная чахотка и генерализованный туберкулез (диссеминированный или милиарный туберкулез с гранулемами в различных органах).
Вторичный туберкулез.Вторичный туберкулезный процесс - реактивация возбудителя в результате ослабления резистентности наблюдается при стрессах, нарушениях питания и у лиц пожилого возраста. Возникают очаги казеозного распада в легких с образованием полостей, поражение бронхов, мелких кровеносных сосудов.
Иммунитет. В основе нестерильного инфекционного и вакцинального иммунитета при туберкулезе - клеточный иммунитет в виде гиперчувствительности замедленного типа (ГЗТ), опосредуемой Т- лимфоцитами и макрофагами. Т- лимфоциты при участии белков главной системы гистосовместимости класса I распознают клетки, инфицированные микобактериями туберкулеза, атакуют и разрушают их. Антибактериальные антитела связываются с различными антигенами возбудителя, образуют циркулирующие иммунные комплексы (ЦИК) и способствуют их удалению из организма.
Аллергическая перестройка (ГЗТ) к туберкулезной палочке свидетельствует о формировании приобретенного иммунитета и может быть выявлена с помощью туберкулиновой пробы. Эта проба является достаточно специфичной. Старый туберкулин Коха представляет концентрированный фильтрат стерилизованных компонентов микобактерий. Очищенный препарат PPD (новый туберкулин Коха, содержащий туберкулопротеины) используют преимущественно для постановки внутрикожной пробы Манту. С помощью этой пробы проводят отбор лиц, подлежащих ревакцинации. Положительный результат пробы Манту нельзя рассматривать как обязательный признак активного процесса (это на самом деле показатель ГЗТ), а отрицательный - не всегда свидетельствует об его отсутствии (анергия, иммунодефициты).
Иммунопрофилактика включает внутрикожное введение аттенуированного штамма Mycobacterium bovis, известного как бацилла Кальметта - Жерена (БЦЖ). В России вакцинацию проводят новорожденным (на 5-7 дни жизни), ревакцинацию - в 712-17-22 лет и более старших возрастах при отрицательной пробе Манту (т.е. отсутствии клеточного нестерильного = вакцинального или инфекционного иммунитета - ГЗТ).
Лабораторная диагностика. Применяют микроскопические, бактериологические, биологические, аллергологические, серологические и молекулярно - генетические методы.
Микроскопическая диагностика включает микроскопию нативного материала, использование методов накопления, люминесцентную диагностику. Микроскопия нативного патологического материала (мокрота, отделяемое свищей, промывные воды из бронхов, моча) в мазках, окрашенных по Цилю - Нильсену, позволяет выявлять красные кислотоустойчивые палочки при концентрации микобактерий не менее нескольких сот тысяч / мл. Методы накопления (например, флотации) повышают чувствительность микроскопии до нескольких тысяч микробных тел / мл. Люминесцентная микроскопия с использованием акридинового оранжевого или аурамина - родамина - наиболее чувствительный и эффективный метод бактериоскопии, чувствительность - 500-1000 микобактерий / мл. Позволяет выявлять микобактерии с измененными культуральными и тинкториальными свойствами.
Бактериологический метод (посев на питательные среды) позволяет обнаружить микобактерии при концентрации 200-300 / мл. Наиболее эффективен до или в начале лечения, в конце лечения уступает по эффективности люминесцентному методу. Недостаток - длительность получения результатов - от 2 до 12 недель. Достоинство - возможность оценки вирулентности культуры, определение чувствительности к лекарственным препаратам. Разработаны ускоренные методы выделения. По методу Прайса материал помещают на предметное стекло, обрабатывают серной кислотой, отмывают физиологическим раствором и вносят в питательную среду с цитратной кровью. Стекло вынимают через 3-4 суток и окрашивают по Цилю - Нильсену.
Золотой стандарт - биологическая проба на морских свинках, позволяет определять до 10 микобактерий в мл. Распространение резистентных и измененных микобактерий снизило чувствительность метода. Метод требует соблюдения режимных условий и применяется в крупных специализированных лабораториях.
Аллергологические методы - это широко используемые кожные пробы с туберкулином и методы аллергодиагностики in vitro (РТМЛ, ППН - показатель повреждения нейтрофилов и др.).
Серологические методы многочисленны (РСК, РА, РПГА), однако в связи с недостаточной специфичностью используют мало.
Наиболее совершенны генетические методы, в практических лабораториях их используют пока недостаточно.
Среди методов идентификации микобактерий наибольшую практическую ценность имеют два подхода:
- методы дифференциации M.tuberculosis и M.bovis от прочих микобактерий; - методы дифференциации M.tuberculosis и M.bovis.
Существует ряд методов дифференциации микобактерий двух основных видов от остальных. Из них наиболее простым и доступным является оценка роста на яичной среде, содержащей салициловый натрий в концентрациях 0,5 и 1,0 мг/мл. На этих средах, в отличии от других микобактерий, M.tuberculosis и M.bovis не растут.
Для дифференциации M.tuberculosis от всех других видов микобактерий , в том числе от M.bovis, используют ниациновый тест (определение синтезируемой M.tuberculosis в больших количествах никотиновой кислоты, выявляемой с помощью цианистых или роданистых соединений по ярко- желтому окрашиванию). У микобактерий туберкулеза также отмечается положительный тест восстановления нитратов. Учитывают скорость роста и характер пигментообразования. Используют цитохимические методы, позволяющие выявлять корд - фактор (вирулентность) по прочности связи красителей - нейтрального красного или нильского голубого при обработке щелочью.
Лекция №6. Семейство Enterobacteriaceae. Род Salmonella.
Общая характеристика семейства энтеробактерий.
Бактерии этого семейства являются наиболее частыми возбудителями кишечных инфекций. Их объединяет ряд общих признаков. Это короткие, не образующие спор, палочки с закругленными концами, подвижные (перитрихи) или неподвижные, некоторые имеют капсулы. Аэробы или факультативные анаэробы. Характерна отрицательная окраска по Граму. Хорошо растут на обычных питательных средах с мясном экстрактом. На большинстве плотных сред энтеробактерии образуют круглые выпуклые блестящие S- (гладкие) колонии, а также часто обусловленные потерей капсулы плоские, неровные и зернистые R- (шероховатые) формы. Для них характерна ферментация глюкозы (и других углеводов) с образованием кислоты и газа. По отношению к лактозе их делят на лактоза- ферментирующие и лактоза - неферментирующие. Каталаза - положительны, восстанавливают нитраты в нитриты.
Семейство энтеробактерий включает более 20 родов, объединяющих более 100 видов бактерий, обитающих в почве, на растениях, входящих в состав микробных биоценозов кишечников животных и человека. Наибольшее значение для человека имеют рода Escherichia, Salmonella, Shigella, Yersinia, Proteus, Klebsiella и др. Для дифференциации родов используют в основном биохимические признаки, для классификации внутри родов и видов - изучение антигенной структуры (О-, Н- и К- антигенов).
О- антиген представлен липополисахаридами (ЛПС) наружной мембраны. Штаммы, лишенные О- антигена, образуют R- колонии и обычно авирулентны.
Н- антиген - термолабильные белки, имеются только у подвижных (имеющих жгутики) видов.
К- антиген - термостабильные полисахариды капсулы и наружной оболочки.
В патогенезе поражений, вызываемых энтеробактериями, имеют значение ЛПС (эндотоксин, освобождающийся при разрушении бактерий), различные энтеротоксины, факторы инвазивности и адгезии (жгутики и др.), ферменты патогенности.
Род Salmonella.
Сальмонеллы - большая группа энтеробактерий, среди которых различные серотипы - возбудители брюшного тифа, паратифов А, В и С и наиболее распространенных пищевых токсикоинфекций - сальмонеллезов. По признаку патогенности для человека сальмонеллы разделяют на патогенные для человека- антропонозы (вызывают брюшной тиф и паратифы А и В) и патогенные для человека и животных - зоонозы (вызывают сальмонеллезы). Несмотря на значительные различия сальмонелл по антигенным характеристикам, биохимическим свойствам, вызываемым ими заболеваниям, по современной, но недостаточно удобной и совершенной классификации выделяют два вида - S.bongori и S.enteritica. Последний разделен на подвиды, из которых наибольшее значение имеют подвиды choleraesuis и salamae. Подвид choleraesuis включает наибольшую часть известных сероваров сальмонелл (около 1400 из примерно 2400).
Морфология. Прямые грамотрицательные палочки размером 2-4 x 0,5 мкм. Подвижны благодаря наличию перитрихиально расположенных жгутиков.
Культуральные и биохимические свойства. Факультативные анаэробы, хорошо растут на простых питательных средах. Оптимум рН- 7,2-7,4, температуры - +37. Метаболизм - окислительный и бродильный. Сальмонеллы ферментируют глюкозу и другие углеводы с образованием кислоты и газа (серотип Salmonella typhi газообразования не вызывает). Обычно не ферментируют лактозу (на средах с этим углеводом - безцветные колонии), сахарозу. Оксидаза- отрицательны, каталаза - положительны. Реакция Фогеса - Проскауэра отрицательна.
На основании биохимических (ферментативных) свойств сальмонеллы разделены на четыре группы. Характерные признаки сальмонелл - образование сероводорода, отсутствие продукции индола и аэробность. Для выделения используют дифференциально - диагностические среды (висмут - сульфит агар, среды Эндо, Плоскирева, SS агар) и среды обогащения (селенитовый бульон, желчный бульон, среда Раппопорта). S- формы образуют мелкие (от 1 до 4 мм) прозрачные колонии (на среде Эндо - розоватые, на среде Плоскирева - безцветные, на висмут - сульфит агаре - черные, с металлическим блеском). На жидких средах S- формы дают равномерное помутнение, R- формы - осадок.
Антигенная структура. Выделяют О-, Н- и К- антигены. К группе К- антигенов относят Vi- антигены (антигены вирулентности). Благодаря более поверхностному расположению (чем О- антигены) Vi- антиген может препятствовать агглютинации культур сальмонелл О- специфической сывороткой (экранирование). Для дифференциации сальмонелл применяют схему (серологическую классификацию) Кауфманна - Уайта.
В соответствии со структурой О- антигенов сальмонеллы подразделяют на О- группы (67 серогрупп), в каждую из которых входят серологические типы, отличающиеся строением Н- антигенов. Принадлежность сальмонелл к определенному серовару устанавливают при изучении антигенной структуры в соответствии со схемой Кауфманна - Уайта. Примеры: серотип S.paratyphi A относится к серогруппе А,
S.paratyphi В - к серогруппе В, S.paratyphi С - к группе С, S.typhi - к серогруппе D.
Факторы патогенности.
1.Факторы адгезии и колонизации.
2. Способность к внутриклеточному паразитированию, препятствовать фагоцитозу, размножаться в клетках лимфоидной ткани выражены у возбудителей брюшного тифа, паратифов А и В, способствуя хроническому носительству.
3.Эндотоксин (ЛПС).
4. Термолабильные и термостабильные энтеротоксины.
5. Цитотоксины.
6. Существенное значение имеют плазмиды вирулентности и R- плазмиды.
7. Vi - антиген ингибирует действие сывороточных и фагоцитарных бактериоцидных факторов.
Основными факторами патогенности сальмонелл является их способность проникать в макрофаги и размножаться в лимфоидных образованиях собственно слизистого слоя тонкого кишечника (пейеровы бляшки, солитарные фолликулы), а также продукция эндотоксина.
Патогенез поражений. Различия клинических форм заболеваний, вызываемых сальмонеллами, зависит от вирулентности и дозы возбудителя и состояния иммунной системы организма. Обычная доза, вызывающая клинические проявления - 106 - 109 бактерий, меньшая доза достаточна при иммунодефицитах, гипохлоргидрии и других заболеваниях желудочно - кишечного тракта.
Выделяют следующие основные формы сальмонеллезной инфекции:
- гастроинтестинальную;
- генерализованную (тифоподобный и септикопиемический варианты); - бактерионосительство (острое, хроническое, транзиторное).
Существенные патогенетические особенности инфекционного процесса, вызываемого серотипами S.typhi, S.paratyphi A,B, являются основанием для выделения тифо- паратифозных заболеваний в самостоятельную нозологическую группу. Каждой фазе патогенеза соответствует клинический период заболевания и своя тактика лабораторного обследования. Основные фазы - внедрения возбудителя (соответствует инкубационному периоду), первичной локализации возбудителя (продромальный период), бактеремии (первая неделя заболевания), вторичной локализации сальмонелл (разгар заболевания - 2-3 недели), выделительно- аллергическая (реконвалесценция - 4 неделя заболевания).
Проникшие через рот сальмонеллы попадают в эпителиальные клетки двенадцатиперсной и тонкой кишки посредством эндоцитоза. Они легко проникают в эпителиальные клетки, но не размножаются здесь, а проходят и размножаются в лимфатическом аппарате тонкого кишечника. Сальмонеллы размножаются преимущественно в lamina propria (первичная локализация), что сопровождается местной воспалительной реакцией слизистой оболочки, притоком жидкости в очаг поражения и развитием диарейного синдрома (гастроэнтерит). Энтеротоксины повышают уровень циклического аденомонофосфата (цАМФ), происходит повышение уровня гистамина и других биологически активных веществ, проницаемости сосудов. Наблюдаются водно - электролитные нарушения, развиваются гипоксия и ацидоз, которые усугубляют патологический процесс с преобладанием сосудистых растройств. Происходит разрушение части сальмонелл с выделением эндотоксина, сенсибилизация (ГЗТ) лимфатического аппарата тонкого кишечника.
Из слизистой оболочки сальмонеллы могут попадать в лимфо- и далее в кровоток, вызывая бактеремию. В большинстве случаев она носит транзиторный характер, т.к. сальмонеллы элиминируются фагоцитами.
В отличие от других сальмонелл, возбудители брюшного тифа и паратифов, проникнув в кровоток, способны выживать и размножаться в фагоцитах. Они могут размножаться в мезентериальных лимфоузлах, печени и селезенке и вызывать генерализацию процесса. После гибели фагоцитов сальмонеллы вновь поступают в кровь. При этом Vi- антиген ингибирует бактерицидные факторы.
При гибели сальмонелл освобождается эндотоксин, угнетающий деятельность центральной нервной системы (тиф - от греч. typhos - туман, спутанное сознание) и вызывающий длительную лихорадку. Действие эндотоксина может вызвать миокардит, миокардиодистрофию, инфекционно - токсический шок.
В результате бактеремии происходит генерализованное инфицирование желчного пузыря, почек, печени, костного мозга, твердых мозговых оболочек (вторичная локализация сальмонелл). Происходит вторичная инвазия эпителия кишечника, особенно пейеровых бляшек. В сенсибилизированной сальмонеллами стенке развивается аллергическое воспаление с образованием основного грозного осложнения - брюшнотифозных язв. Наблюдается длительное носительство сальмонелл в желчном пузыре с выделением возбудителя с испражнениями, пиелонефриты, кровотечения и перфорации кишечника при поражении пейеровых бляшек. Затем происходит формирование постинфекционного иммунитета, элиминация возбудителя и заживление язв или формирование бактерионосительства (в Западной Сибири часто на фоне хронического описторхоза).
Возбудителями сальмонеллезов являются другие серотипы сальмонелл, патогенные для человека и животных (S.typhimurium, S.enteritidis, S.heldelberg, S. newport и другие). В основе патогенеза сальмонеллезов - действие самого возбудителя (его взаимодействия с организмом хозяина) и эндотоксина, накапливающегося в пищевых продуктах, инфицированных сальмонеллами. В классическом варианте сальмонеллезная токсикоинфекция - гастроэнтерит. Однако при прорыве лимфатического барьера кишечника могут развиваться генерализованные и внекишечные формы сальмонеллезов (менингит, плеврит, эндокардит, артрит, абсцессы печени и селезенки, пиелонефрит и др.). Увеличение генерализованных и внекишечных форм сальмонеллезов связано с увеличением количества иммунодефицитных состояний, что имеет особое значение при ВИЧ- инфекции.
Отдельную проблему представляют госпитальные штаммы сальмонелл (чаще отдельные фаговары S.typhimurium), вызывающие вспышки внутрибольничных инфекций преимущественно среди новорожденных и ослабленных детей. Они передаются преимущественно контактно- бытовым путем от больных детей и бактерионосителей, обладают высокой инвазивной активностью, часто вызывая бактеремию и сепсис. Эпидемические штаммы характеризуются множественной лекарственной устойчивостью (R- плазмиды), высокой резистентностью, в том числе к действию высоких температур.
Эпидемиологические особенности. Характерно повсеместное распространение.
Основные резервуары сальмонелл - человек (возбудители брюшного тифа и паратифа А) и различные животные (остальные серотипы сальмонелл). Основные возбудители отличаются полипатогенностью. Основные источники заражения - мясные и молочные продукты, яйца, птице- и рыбопродукты. Основные пути передачи - пищевой и водный, реже - контактный. Характерна чрезвычайная множественность резервуаров и возможных источников инфекции. Основное значение имеют сельскохозяйственные животные и птицы.
Лабораторная диагностика. Основной метод - бактериологический. Исходя из патогенеза оптимальными сроками бактериологических исследований при гастроинтестинальных формах являются первые дни, при генерализованных формах - конец второй - начало третьей недели заболевания. При исследовании различных материалов (испражнения, кровь, моча, желчь, рвотные массы, пищевые остатки) наибольшая частота положительных результатов отмечается при исследовании испражнений, для возбудителя брюшного тифа и паратифов - крови (гемокультура).
Исследования проводят по стандартной схеме. Исследуемый материал засевают на плотные дифференциально - диагностические среды - высокоселективные (висмут- сульфит агар, агар с бриллиантовым зеленым), среднеселективные (среда Плоскирева, слабощелочной агар), низкоселективные (агары Эндо и Левина) и в среды обогащения. Для посева крови используют среду Рапопорт. На висмут- сульфит агаре колонии сальмонелл приобретают черный (реже- зеленоватый) цвет.Выросшие колонии пересевают на среды для первичной (среды Ресселя) и биохимической (сероводород, мочевина, глюкоза, лактоза) идентификации. Для предварительной идентификации используют О1- сальмонеллезный фаг, к которому чувствительно до 98% сальмонелл.
Для идентификации культур в РА используют поливалентные и моновалентные О-,
Н- и Vi- антисыворотки. Сначала используют поливалентные адсорбированные О- и Н- сыворотки, а затем- соответствующие моновалентные О- и Н- сыворотки. Для идентификации возбудителей брюшного тифа и паратифов используют антитела к антигену О2 (S.paratyphi A), O4 (S.paratyphi B), O9 (S.typhi). Если культура не агглютинируется О- сывороткой, ее нужно исследовать с Vi- сывороткой. Для быстрого выявления сальмонелл используют поливалентные люминесцентные сыворотки.
Серологические исследования проводят для диагностики, а также выявления и дифференциации различных форм носительства. Применяют РА (реакцию Видаля) с О- и Н- диагностикумами и РПГА с применением поливалентных эритроцитарных диагностикумов, содержащих полисахаридные антигены серогрупп А,В,С,Д и Е и Vi- антиген.
Лечение - антибиотики (левомицетин и др.). Часто выявляют резистентные к антибиотикам штаммы. Необходимо определять антибиотикорезистентность выделенных культур.
Специфическая профилактика может применяться преимущественно в отношении брюшного тифа. Применяют химическую сорбированную брюшнотифозную моновакцину. Вакцинацию в настоящее время применяют преимущественно по эпидемическим показаниям.
Лекция №7. Эшерихии и шигеллы.
Род Escherichia.
Эшерихии - наиболее распространенные аэробные бактерии кишечника, способные при определенным условиях вызывать обширную группу заболеваний человека, как кишечной (диарея), так и внекишечной (бактеремия, инфекции мочевыводящих путей и др.) локализации. Основной вид - E.coli (кишечная палочка) - самый распространенный возбудитель инфекционных заболеваний, вызываемых энтеробактериями. Этот возбудитель является показателем фекального загрязнения, особенно воды. Коли - титр и коли - индекс часто использовали как санитарные показатели. Эшерихии входят в состав микрофлоры толстого кишечника млекопитающих, птиц, пресмыкающихся и рыб.
Культуральные свойства. На жидких средах E.coli дает диффузное помутнение, на плотных средах образует S- и R- формы колоний. На основной для эшерихий среде Эндо лактозоферментирующие кишечные палочки образуют интенсивно красные колонии с металлическим блеском, не ферментирующие - бледно- розовые или бесцветные колонии с более темным центром, на среде Плоскирева - красные с желтоватым оттенком, на среде Левина - темно- синие с металлическим блеском.
Биохимические свойства. Кишечная палочка в большинстве случаев ферментирует углеводы (глюкозу, лактозу, маннит, арабинозу, галактозу и др.) с образованием кислоты и газа, образует индол, но не образует сероводород, не разжижает желатин.
Антигенная структура. Какие - либо существенные морфологические различия между патогенными и непатогенными кишечными палочками не обнаружены. Их дифференциация основана на изучении антигенных свойств. Среди поверхностных антигенов выделяют полисахаридные О- антигены, жгутиковые Н- антигены и капсульные полисахаридные К- антигены. Известно более 170 вариантов О- антигенов (это соответствует принадлежности возбудителя к определенной серогруппе) и 57 - Н- антигенов (принадлежность к серовару). В состав диареегенных (вызывающих диарею) кишечных палочек входят 43 О- группы и 57 ОН- вариантов.
Основные факторы патогенности диареегенных E.coli.
1. Факторы адгезии, колонизации и инвазии, связанные с пилями, фимбриальными структурами, белками наружной мембраны. Они кодируются плазмидными генами и способствуют колонизации нижних отделов тонкой кишки.
2. Экзотоксины: цитотонины (стимулируют гиперсекрецию клетками кишечника жидкости, нарушают водно - солевой обмен и способствуют развитию диареи) и энтероцитотоксины (действуют на клетки стенки кишечника и эндотелия капилляров).
3. Эндотоксин (липополисахарид).
В зависимости от наличия различных факторов патогенности диареегенные кишечные палочки разделены на пять основных типов: энтеротоксигенные, энтероинвазивные, энтеропатогенные, энтерогеморрагические, энтероадгезивные.
4. Для патогенных кишечных палочек характерна выработка бактериоцинов (колицинов).
Энтеротоксигенные E.coli имеют высокомолекулярный термолабильный токсин, схожий по действию с холерным, вызывают холероподобную диарею (гастроэнтериты у детей младшего возраста, диарею путешественников и др.).
Энтероинвазивные кишечные палочки способны проникать и размножаться в клетках эпителия кишечника. Вызывают профузную диарею с примесью крови и большим количеством лейкоцитов (показатель инвазивного процесса) в испражнениях. Клинически напоминает дизентерию. Штаммы имеют некоторое сходство с шигеллами (неподвижные, не ферментируют лактозу, обладают высокими энтероинвазивными свойствами).
Энтеропатогенные E.coli - основные возбудители диареи у детей. В основе поражений - адгезия бактерий к эпителию кишечника с повреждением микроворсинок. Характерна водянистая диарея и выраженное обезвоживание.
Энтерогеморрагические кишечные палочки вызывают диарею с примесью крови (геморрагический колит), гемолитико - уремический синдром (гемолитическая анемия в сочетании с почечной недостаточностью). Наиболее частый серотип энтерогеморрагических кишечных палочек - О157: Н7.
Энтероадгезивные E.coli не образуют цитотоксины, слабо изучены.
Эпидемиология. Основной механизм распространения диареегенных кишечных палочек - фекально - оральный. Заражение может происходить через пищу, воду, при уходе за животными. Поскольку эшерихии обитают в кишечниках многих видов животных, конкретный источник заражения установить сложно. Контактный путь заражения может быть в закрытых заведениях. Энтеропатогенные и энтероинвазивные E.coli - наиболее частые причины внутрибольничных вспышек эшерихиозов.
Лабораторная диагностика. Основным подходом является выделение чистой культуры на дифференциально - диагностических средах и ее идентификация по антигенным свойствам. Ставят РА с набором поливалентных ОК (к О- и К- антигенам) сывороток, затем - адсорбированных О- сывороток и прогретыми при 100 градусах Цельсия (для разрушения К- антигенов) культурами.
Биохимическая дифференциация имеет дополнительное значение. Идентификация диареегенных типов возможна при выявлении специфических маркеров (энтерогеморрагические кишечные палочки не ферментируют сорбит, а серовар О157: Н7 не проявляет бета - глюкуронидазной активности).
Род Shigella.
Шигеллы - кишечные патогены человека и приматов, которые вызывают бактериальную дизентерию или шигеллезы. В соответствии с антигенной структурой О- антигена и биохимическими свойствами известные серотипы шигелл разделяют на четыре вида или серогруппы - S.dysenteriae (серогруппа А), S.flexneri (серогруппа В), S.boydii (серогруппа С) и S.sonnei (серогруппа Д).
По морфологическим признакам шигеллы не отличаются от остальных энтеробактерий. Это неподвидные факультативно - анаэробные грамотрицательные палочки.
Биохимические свойства. Шигеллы по сравнению с другими кишечными бактериями биохимически малоактивны. Не образуют сероводород на трехсахарно - железном агаре, не ферментируют мочевину.
Наименьшей ферментативной активностью обладают штаммы S.dysenteriae (серогруппа А), ферментирующие только глюкозу без газообразования, в отличие от других шигелл этот вид является маннит - отрицательным.
Шигеллы Флекснера ферментируют маннит, образуют индол, но не ферментируют лактозу, дульцит и ксилозу. Серотип Ньюкасл разделен на три биохимических типа.
Для шигелл Флекснера более характерен водный путь передачи.
Шигеллы Бойда (серогруппа С) имеют близкую биохимическую активность, однако ферментируют дульцит, ксилозу и арабинозу. Имеют ряд серотипов, каждый из которых имеет свой главный типовой антиген.
Шигеллы Зонне (серогруппа Д) способны медленно ферментировать лактозу и сахарозу, имеют биохимические типы и фаготипы. Основной путь передачи - пищевой (чаще через молоко и молочные продукты).
Антигенная структура. У шигелл имеются О- и К- антигены. О- антигены имеют эпитопы различной специфичности - от общих для семейства энтеробактерий до типоспецифических. В классификации учитывают только термостабильные групповые (четыре группы или вида - А,В,С и Д) и типоспецифические (деление на серотипы). К термолабильным антигенам относятся К- антигены (они имеются в группах А и С) и фимбриальные антигены (у шигелл Флекснера они близки в антигенном отношении E.coli). Определение антигенной структуры необходимо для окончательной идентификации.
Эпидемиология. Шигеллы достаточно устойчивы во внешней среде. Источник инфекции - человек с различными формами клинического проявления шигеллезов. Механизм заражения - фекально - оральный. Для различных видов шигелл характерны преобладающие пути передачи (контактно- бытовой - для S.dysenteriae, пищевой - для S.sonnei, водный - для S.flexneri). Для эпидемического процесса характерна изменение структуры циркулирующих популяций возбудителей - смена ведущих видов, биоваров, сероваров, что связано как с изменениями популяционного иммунитета, так и с изменениями свойств возбудителя, особенно с приобретением различных плазмид (R, F, Col и др.). Инфицирующая доза - порядка 200 - 300 шигелл. Более легкое течение имеет дизентерия, вызванная шигеллами Зонне.
Факторы патогенности и патогенез поражений. Главная биологическая характеристика шигелл - способность внедряться в эпителиальные клетки, размножаться в них и вызывать их гибель. Формирование очага в слизистой нисходящего отдела толстого кишечника (сигмовидная и прямая кишки) носит циклический характер: адгезия, колонизация, внедрение шигелл в цитоплазму энтероцитов, размножение, разрушение и отторжение эпителиальных клеток, выход шигелл в просвет кишечника, снова адгезия и т.д.
Роль факторов адгезии и колонизации выполняют пили, белки наружной мембраны, ЛПС, ферменты - нейраминидаза, муциназа, гиалуронидаза (разрушают слизь).
Шигеллы имеют целый ряд факторов инвазии и устойчивости к действию механизмов защиты (К- антиген, ЛПС и др.), контролизуемых хромосомными генами шигелл и плазмидами.
Шигеллы имеют различные токсины. Они имеют эндотоксин и шигаподобные цитотоксины (SLT-1, SLT-2). Цитотоксины обусловливают разрушение клеток, энтеротоксин - диарею, эндотоксин - общую интоксикацию. Токсин Шига вызывает нарушение синтеза белка, всасывания ионов натрия и воды, приток жидкости в очаг воспаления.
Наиболее типичные признаки дизентерии - понос, тенезмы (болезненные спазмы прямой кишки) и частые позывы, общая интоксикация. Характер стула определяется степенью поражения толстого кишечника.
Постинфекционный иммунитет - прочный, типоспецифический.
Лабораторная диагностика. Основной метод диагностики - бактериологический. Производят посев испражнений на дифференциально - диагностические среды Эндо и Плоскирева для получения изолированных колоний. Чистые культуры изучают по биохимическим свойствам, идентификацию проводят в РА с поли- и моновалентными сыворотками. Если выделенная культура обладает биохимическими свойствами шигелл, но не агглютинирует сыворотки к О- антигенам, ее нужно прокипятить 30 минут для разрушения термолабильных К- антигенов, часто препятствующих агглютинации шигелл серогрупп А и С (т.е. имеющих К- антигены), и снова исследовать в РА.
Для серологической диагностики используют РПГА с групповыми эритроцитарными диагностикумами.
Лекция №8. Представители родов Vibrio, Campylobacter, Helicobacter.
Род Vibrio.
Семейство Vibrionaceae объединяет подвижные, изогнутые палочковидные бактерии, обладающие полярными жгутиками. Эволюционно происходят из водных бактерий, широко распространены в пресной и морской воде, у беспозвоночных и позвоночных хозяев. Патогенные для человека виды относят к родам Vibrio, Aeromonas и Plesiomonas.
Для рода Vibrio характерны короткие прямые или изогнутые грамотрицательные палочки, подвижные, не образующие спор и капсул, хорошо растущие на обычных средах. Они ферментируют углеводы с образованием кислоты без газа, чувствительны к вибриостатику О/129. Можно культивировать при температуре от плюс 18 до 37о С, рН 8,6-9,0.
От других родов семейства представители рода Vibrio дифференцируют по биохимическим тестам. Род насчитывает более 25 видов, из них основное значение имеет Vibrio cholerae- возбудитель холеры, а также V.parahaemolyticum, V.vulnificus.
Vibrio cholerae.
Морфология. Холерный вибрион имеет один полярный жгутик, часто напоминает запятую (запятая Коха). Важный диагностический признак - подвижность (определяют микроскопией по методу висячей или раздавленной капли). Морфологически изменчивы. Хорошо окрашиваются водным фуксином Пффейфера и карболовым фуксином Циля.
Культуральные свойства. Факультативный анаэроб. Холерный вибрион неприхотлив к питательным средам. Хорошо размножается на 1% щелочной (рН 8,6-9,0) пептонной воде, опережая бактерии кишечной группы (среда обогащения), образует нежную голубоватую пленку и муть. Для подавления роста протея и некоторых других микроорганизмов используют пептонную воду с добавлением теллурита калия.
На плотных средах холерный вибрион образует гладкие стекловидные, прозрачные с голубоватым оттенком дисковидные колонии вязкой консистенции. Используют щелочной агар, желчно - солевой агар, щелочной агар с кровью, наилучшей является TCBS - агар (агар с тиосульфатом, цитратом, солями желчных кислот и сахарозой).
Биохимические свойства. Холерный вибрион сбраживает с образованием кислоты без газа многие углеводы (глюкозу, сахарозу, маннозу, маннит, лактозу, левулезу, гликоген, крахмал). По отношению к трем сахарам (триада Хейберга) - сахарозе, маннозе и арабинозе вибрионы делят на восемь биохимических групп, холерный вибрион относится к первой группе (разлагает сахарозу и маннозу).
Холерный вибрион разлагает желатин, казеин, свертывает молоко и разлагает белковые препараты до индола и аммиака.
Вид V.cholerae делят на биотипы, серогруппы и серовары.. Основные биотипы - классический (V.cholerae asiaticae) и Эль - Тор (V.cholerae eltor). Серогруппы выделяют по структуре О- антигенов, в основной О1 группе холерных вибрионов выделяют серовары Огава, Инаба и Хикоджима.
Антигенная структура. У холерных вибрионов выделяют термостабильные О- антигены и термолабильные Н- антигены. По структуре О- антигенов выделено 139 серогрупп, биотипы Эль - Тор и классический объединены в 01 группу (типируются 01 - антисывороткой). Изоляты Эль - Тор отличаются гемолитическими свойствами (вызывают гемолиз эритроцитов барана), способностью агглютинировать куриные эритроциты, резистентности к полимиксину, чувствительности к фагам.
О- антиген 01 группы неоднороден и включает общий А- компонент и два типоспецифических - В и С. Соответственно их наличию серовар Огава имеет сочетание АВ, Инаба - АС, Хикоджима - АВС.
Холерный вибрион может переходить из S- в R- форму, не агглютинироваться О- сывороткой. В связи с антигенной структурой для идентификации V.cholerae используют О- сыворотку, OR- сыворотку ( для выявления OR- и R- диссоциантов), типоспецифические сыворотки Инаба (С) и Огава (В). В 90- е годы выявлен новый серовар V.cholerae 0139, не агглютинирующийся вышеуказанными сыворотками, по остальным свойствам мало отличающийся от холерного вибриона 01 группы.
Не типируемые основной 01 сывороткой (т.е. не относящиеся к 01 группе) вибрионы называются неагглютинирующими (НАГ) вибрионами- холероподобными или парахолерными. Они имеют общий с холерным вибрионом Н- антиген, но отличаются по О- антигену.
По Н- антигену выделяют группы А и В, холерные вибрионы входят в группу А. Вирионы группы В (биохимически отличающиеся от холерных) имеют неоднородную структуру О- антигена и подразделяются на шесть серологических подгрупп.
Факторы патогенности холерного вибриона.
1. Подвижность (жгутики) и хемотаксис.
2. Ферменты способствуют адгезии и колонизации, взаимодействию с эпителиальными клетками- муциназа (разжижает слизь), нейтаминидаза (взаимодействие с микроворсинками, создание посадочной площадки), лецитиназа и другие.
3. Эндотоксин - термостабильный липополисахарид, схожий по структуре и свойствам с другими эндотоксинами грамотрицательных бактерий.
4. Экзотоксин - холероген - главный фактор патогенности, термолабильный белок. Синтез холерогена - важнейшее, генетически детерминированное свойство холерного вибриона. Молекула холерогена состоит из двух фрагментов А и В. Собственно токсическую функцию выполняет пептид А1 фрагмента А. Молекула холерогена распознает рецептор энтероцита, проникает в мембрану клетки, активирует аденилатциклазную систему, накапливающийся циклический АМФ вызывает гиперсекрецию жидкости, Na+, HCO3-, K+, Cl- из энтероцитов. Это приводит к характерной для холеры диарее, обезвоживанию и обессоливанию организма.
5. У многих вибрионов, в т.ч. не относящихся к 01 группе, имеются различные энтеротоксины.
6. В патогенезе проявлений холеры имеет значение также фактор, повышающий проницаемость капилляров.
Некоторые особенности эпидемиологии. Холера - кишечная инфекция. Основной источник - человек (больной или вибрионоситель), загрязненная вода. Способ заражения - фекально - оральный. Индивидуальная восприимчивость к холере чрезвычайно вариабельна. Характерно большое количество скрытых (стертых) форм, вибрионосительство. Обнаружение возбудителя в воде напрямую связано с наличием больных или бактерионосителей. Холерные вибрионы 01 группы могут длительно находиться в водных экосистемах в виде некультивируемых форм.
Лабораторная диагностика. Холера относится к группе особо опасных инфекций, культивирование ее возбудителя требует соблюдения особого режима биологической безопасности. Основной метод диагностики - бактериологический, включает выделение и идентификацию возбудителя.
Материал для исследования - испражнения и рвотные массы, секционные материалы от погибших, пробы воды и смывы с объектов окружающей среды, пищевые остатки.
Для посева используют жидкие среды обогащения, щелочной МПА, элективные и дифференциально - диагностические среды (лучше TCBS). В качестве транспортной среды наиболее удобна 1% пептонная вода. Подозрительные стекловидные прозрачные колонии пересевают для получения чистой культуры, которую идентифицируют по морфологическим, культуральным, биохимическим свойствам, подвижности, антигенным свойствам, фаготипируют.
Для ускоренной диагностики применяют иммунолюминесцентный метод, биохимическую идентификацию с набором индикаторных дисков, для обнаружения холерных вибрионов в первичных материалах - РНГА с антительным диагностикумом, для выявления некультивируемых форм - ПЦР, для определения вирулентности и синтеза холерогена - биопробы на кроликах - сосунках, ИФА, ДНК- зонды (выявление фрагмента хромосомы, несущего оперон холерогена).
Специфическая профилактика. Имеются различные вакцины - убитая из сероваров Инаба и Огава, анатоксин холерогена, химическая бивалентная вакцина. Вакцины применяют только по эпидпоказаниям (низкая иммуногенность). Может проводиться антибиотикопрофилактика (превентивная терапия) тетрациклином и другими антибиотиками.
V.parahaemolyticus (парагемолитический вибрион) является галофилом, встречается в морской воде (Японского, Черного, Каспийского и других южных морей). При употреблении морских продуктов, не подвергнутых достаточной термической обработке, этот возбудитель вызывает у людей пищевые токсикоинфекции и дизентерия - подобные заболевания. Вызывает гемолиз на кровяном агаре с повышенной концентрацией хлористого натрия (на 7% NaCl - штаммы с энтеропатогенными свойствами).
V.vulnificus - наиболее патогенный вид для человека из нехолерных вибрионов. Выявляют в морской воде и ее обитателях. Вызывает раневые инфекции, септицемии и другие заболевания. Ферментирует сахарозу и лактозу, на TCBS - агаре образует желтые колонии.
Представители родов Campylobacter и Helicobacter.
Грамотрицательные микроаэрофилы этих двух родов - мелкие, подвижные, не образующие спор, изогнутые (S- образные или похожие на крыло чайки) палочковидные бактерии. Виды, составляющие род Helicobacter, в том числе H.pylori - возбудитель язвенной болезни человека, выделены из рода Campylobacter. Род Campylobacter
Этот род включает 13 видов спиральных бактерий, имеющих один и более завитков. Кампилобактеры - подвижные, обладающие полярными жгутиками (или жгутиком) и винтообразным движением, микроорганизмы. Они не ферментируют и не окисляют углеводы, капрофилы и микроаэрофилы , т.е. требуют повышенной концентрации СО2 и пониженной - О2.
Род включает виды, патогенные для человека и теплокровных животных. Заболевания, ими вызываемые - кампилобактериозы - острые кишечные заболевания, протекающие с поражением желудочно - кишечного тракта. Кампилобактеры часто выделяют из кишечника, ротовой полости и органов мочеполовой системы теплокровных.
Выделяют группу C.jejuni (включает этот вид, а также C.coli, C.lari) или термофильных кампилобактерий. Они отличаются высоким температурным оптимумом роста (+420С).
Среди мезофильных кампилобактерий с оптимумом роста при +370С существенную роль в патологии человека имеет C.fetus (вызывает артриты, менингиты, васкулиты, аборты), а также имеется ряд условно - патогенных видов (C.concisus и C.sputorum - в ротовой полости, C.fennelliae, C.cinaedi и C.hyointestinalis - в толстом кишечнике).
Культуральные свойства. Для кампилобактеров нужны специальные газовые смеси для создания микроаэрофильных условий, рН - 7,0-7,2, мезофильные условия (+420С - для термофилов, +370С - для остальных). Используют специальные питательные среды (мясные, печеночные, кровяные) с добавлением селективных антибиотиков. Для получения более чистых проб для посевов (чаще исследуют копрофильтраты!) можно использовать фильтрацию через мембранные фильтры с диаметром пор 0,65 мкм. На плотных средах образуют два типа колоний - “расползающиеся” с неровными краями или блестящие выпуклые с ровными краями, колонии мелкие.
Биохимические свойства. Инертны к углеводам, редуцируют нитраты, оксидаза - положительны, энергию получают за счет аминокислот и трикарбоновых кислот. Дифференциация видов по биохимическим свойствам основана на гидролизе гиппурата (C.jejuni и C.coli), чувствительности к налидиксовой кислоте (C.jejuni и C.lari), образовании сероводорода и др.
Антигенная структура. Кампилобактеры имеют О- , Н- и К- антигены. Основное значение для серотипирования имеют термостабильные О- антигены.
Эпидемиологические особенности. Кампилобактеры распространены у различных видов млекопитающих и птиц. Основной путь передачи - пищевой. Характерна преимущественно летняя сезонность.
Основные патогенетические факторы. Кампилобактеры характеризуются высокой адгезивной и инвазивной активностью, быстрой колонизацией верхних отделов тонкого кишечника. Важнейшими факторами адгезии являются жгутики и специфические поверхностные адгезины. У этих бактерий имеется эндотоксин, термолабильный энтеротоксин.
Клинические проявления - энтероколит.
Лабораторная диагностика. Микроскопический метод - окраска 1% водным раствором основного фуксина 10- 20 секунд - выявляют S- образные короткие цепочки, “крылья чайки”. Основной метод - бактериологический - посев испражнений. Культуры идентифицируют по совокупности признаков.
Helicobacter pylori.
Язвенная болезнь - заболевание, которое характеризуется наличием язвенного дефекта в слизистой желудка или двенадцатиперсной кишке. Открытие H.pylori обусловило переворот в представлениях об этиологии, патогенезе, лечении и профилактике язвенной болезни. Язвенная болезнь практически в 100% ассоциирована с хеликобактером пилори. Существенную роль имеют также стрессовые факторы и психологические особенности больных, а также генетические факторы в развитии болезни.
Морфологические и культуральные свойства - аналогичны Campylobacter. Предпочитает шоколадный агар.
Биохимические свойства. Имеет уреазу, оксидаза- и каталаза- положителен.
Антигенные свойства. Имеет О- и Н- антигены.
Патогенез поражений. Хеликобактеры проникают через слой слизи (чаще в антральной части желудка и двенадцатиперсной кишке), прикрепляются к эпителиальным клеткам, проникают в крипты и железы слизистой оболочки. Антигены бактерий (в первую очередь ЛПС) стимулируют миграцию нейтрофилов и вызывают острое воспаление. Хеликобактеры локализуются в области межклеточных ходов, что обусловлено хемотаксисом на мочевину и гемин (разрушение гемоглобина эритроцитов в микроциркуляторном русле). Под действием уреазы хеликобактеров мочевина расщепляется до аммиака, с действием которого связано повреждение слизистой оболочки желудка и двенадцатиперсной кишки. Многие ферменты (муциназа, фосфолипаза и др.) также могут стимулировать нарушения целостности слизистых.
К факторами патогенности H.pylori в первую очередь относят факторы колонизации (адгезию, подвижность), факторы персистенции и факторы, вызывающие заболевание. Ключевые факторы в тропизме и патогенности H.pylori включают механизмы адгезии и секреции бактериальных токсинов. Представлены доказательства ведущей роли Lewis B антигена как рецептора для адгезии. Кроме них имеют значение муцины желудка и сульфатиды слизистой желудка. Выявлен белок Bab A возбудителя (адгезин), который позволяет микроорганизму связываться с групповым антигеном крови Lewis B, присутствующим на поверхности клеток эпителия желудка. Другими факторами патогенности являются cag A (цитотоксин- ассоциированный ген) и vac A (вакуолизирующий цитотоксин). Штаммы, экспрессирующие эти маркеры вирулентности, относятся к штаммам первого типа, которые ассоциированы с повышенным ульцерогенным и воспалительным потенциалом, в отличии от штаммов второго типа, не имеющих этих факторов.
Присутствие всех трех факторов (Bab A, cag A, vacA) является существенным для проявления патогенных свойств H.pylori (триплет - позитивные штаммы). Повреждающее действие на слизистую может быть связано как с прямым действием бактериальных токсинов, так и с опосредованным влиянием через иммунную систему. Длительная персистенция возбудителя связана с рядом механизмов, позволяющих преодолевать защитные барьеры слизистой, и способностью формирования кокковых, не обладающих патогенным потенциалом, форм.
H.pylori не всегда приводит к развитию язвенной болезни, однако при язвенной болезни этот возбудитель выявляют постоянно. Факторы, определяющие ульцерогенность H.pylori, усиленно изучаются.
Лабораторная диагностика должна быть комплексной, основанной на нескольких тестах. Методы выявления могут быть инвазивными (связанными с необходимостью взятия биоптата слизистой) и неинвазивными (косвенными).
Основные методы выявления H.pylori в биоптатах слизистой.
1.Микроскопические методы (окраска гематоксилином - эозином, акридиновым оранжевым, по Граму, водным фуксином, инпрегнация серебром; фазово - контрастная микроскопия с определением подвижности).
2.Определение уреазной активности.
3.Выделение и идентификация возбудителя на плотных средах (чаще кровяных). Посевы осуществляют на кровяной агар, кровяной агар с амфотеррицином, эритрит - агар с амфотеррицином. Культивируют 5-7 суток при 37о С в микроаэрофильных, аэробных и анаэробных условиях. Принадлежность определяют по морфологии микроорганизмов и их колоний, винтообразной подвижности, росту в микроаэрофильных условиях и отсутствию роста в аэробных и анаэробных условиях и при температурах +25 и +42о С, по наличию оксидазной, каталазной и уреазной активности.
4.Выявление антигенов возбудителя в ИФА.
5.ПЦР- диагностика - наиболее чувствительный и специфичный тест.
Среди неинвазивных методов - “дыхательный тест”, ИФА для выявления IgG и IgA - антител.
Лечение комплексное, с применением методов санации (эррадиации возбудителя). Применяют Де- НОЛ (коллоидный субцитрат висмута), ампициллин, трихопол (метронидазол) и др.
Лекция №9. Возбудители сибирской язвы и бруцеллеза.
Эти возбудители вызывают инфекции, относящиеся к группе особо опасных (из бактериальных инфекций к их числу относят чуму, холеру, сибирскую язву, туляремию, сап и бруцеллез).
Сибирская язва.
Возбудитель сибирской язвы - Bacillus anthracis относится к роду Bacillus семейства Bacillaceae (к бациллам).
Морфология. Крупная грамположительная палочка, часто с закругленными концами. В отличии от других бацилл - неподвижна, хорошо окрашивается анилиновыми красителями. В клинических материалах расположены парами или в виде коротких цепочек, окруженных общей капсулой (образуется только в организме человека и животных или на специальных средах с кровью, сывороткой крови). На средах возбудитель образует длинные цепочки в виде “бамбуковой трости” (с уточщениями на концах и сочленениями клеток). На агаре, содержащем пенициллин, происходит разрушение клеточных стенок, образуются шаровидные протопласты в виде цепочек (“жемчужное ожерелье”). Возбудитель сибирской язвы образует эндоспоры, которые располагаются центрально, их диаметр не превышает диаметра бактериальной клетки. Споры образуются только вне организма, при наличии (доступе) кислорода и определенной температуре (от +12 до +43о С, оптимум при 30-35о С). Споры проявляют очень высокую устойчивость во внешней среде (десятилетия). Сибирская язва - прежде всего почвенная инфекция.
Культуральные свойства. Возбудитель растет в аэробных и факультативно - анаэробных условиях. Температурный оптимум +37о С, рН -7,2-7,6. Растет на простых питательных средах, в т.ч. на картофеле, настое соломы, экстрактах злаковых и бобовых культур. Дает характерный рост при посеве уколом в желатин (“перевернутая елочка”). Вирулентные R- формы на плотных средах образуют шероховатые серовато - белые колонии волокнистой структуры (“голова медузы” или “львиная грива”). На жидких средах образуется осадок в виде комочка ваты. Возбудитель сибирской язвы может образовывать также гладкие (S), слизистые (М) или смешанные (SM) колонии, особенно в микроаэрофильных условиях. В S- форме возбудитель утрачивает вирулентность.
Биохимические свойства. B.anthracis биохимически высоко активна. Она ферментирует с образованием кислоты без газа глюкозу, сахарозу, мальтозу, трегалозу, образует сероводород, свертывает и пептонизирует молоко.
Антигенная структура. Выделяют три основных группы антигенов - капсульный антиген, токсин (кодируются плазмидами, при их отсутствии штаммы авирулентны), соматические антигены.
Капсульные антигены отличаются по химической структуре от К- антигенов других бактерий, полипептидной природы, образуются преимущественно в организме хозяина.
Соматические антигены - полисахариды клеточной стенки, термостабильны, долго сохраняются во внешней среде, трупах. Выявляют их в реакции термопреципитации Асколи.
Токсин включает протективный антиген (индуцирует синтез защитных антител), летальный фактор, отечный фактор.
Факторы патогенности - капсула и токсин.
Краткая эпидемиологическая характеристика. Сибирская язва - зоонозная инфекция. Основной источник для человека - травоядные животные. Их заражение происходит преимущественно алиментарным путем, споры длительно сохраняются в почве и заглатываются животными преимущественно с кормами, травой). Особую опасность представляют сибиреязвенные скотомогильники (в них споры длительно сохраняются, при их разрывании, размывании и других процессах попадают на поверхность почвы и растения). Человек заражается при контакте с инфицированным материалом (уход за больными животными, разделка и употребление в пищу инфицированных мясных продуктов, контакт со шкурами сибиреязвенных животных и др.).
Основные формы клинического проявления зависят от входных ворот инфекции - кожная (карбункул), кишечная, легочная, септическая. Характерна высокая летальность (меньше при кожной форме).
Лабораторная диагностика. Материал для исследования от больных зависит от клинической формы. При кожной форме исследуют содержимое пузырьков, отделяемое карбункула или язвы, при кишечной - испражнения и мочу, при легочной - мокроту, при септической - кровь. Исследованию подлежат объекты внешней среды, материал от животных, пищевые продукты.
Бактериоскопический метод используют для обнаружения грамположительных палочек, окруженных капсулой, в материалах от человека и животных, спор - из объектов внешней среды. Чаще применяют метод флюоресцирующих антител (МФА), позволяющий выявлять капсульные антигены и споры.
Основной метод - бактериологический применяется в лабораториях особо опасных инфекций по стандартной схеме с посевом на простые питательные среды (МПА, дрожжевая среда, среда ГКИ), определением подвижности, окраской по Граму и изучением биохимических особенностей. В дифференциации от других представителей рода Bacillus существенное значение имеет биологическая проба. Белые мыши погибают в пределах двух суток, морские свинки и кролики - в течение четырех суток. Определяют также лизабельность бактериофагами, чувствительность к пенициллину (жемчужное ожерелье). Для ретроспективной диагностики используют серологические тесты, аллергическая проба с антраксином, для выявления соматического антигена - реакция Асколи, которая может быть результативна при отрицательных результатах бактериологических исследований.
Лечение. Применяют противосибиреязвенный иммуноглобулин, антибиотики (пенициллины, тетрациклины и др.).
Профилактика. Применяют живую споровую безкапсульную вакцину СТИ, протективный антиген.
Род Brucella. Бруцеллез.
Род объединяет мелкие неподвижные палочки или коккобациллы, обладающие значительным полиморфизмом. Аэробы, оптимум температуры около +37о С, рН - 6,6- 7,4. Факультативные внутриклеточные паразиты, хорошо окрашиваемые анилиновыми красителями.
Патогенные для человека четыре вида - B.melitensis (распространена преимущественно среди мелкого рогатого скота, вызывает наиболее тяжелые поражения у человека), B.abortus (связана с крупным рогатым скотом), B.suis, B.canis. Основным хозяином B.suis 4 биовара являются северные олени, этот возбудитель часто называют B.rangiferis. От грызунов выделены бруцеллы вида B.neotomae, от овец - B.ovis. B.melitensis разделена на 3 биовара, B.abortus - на 9, B.suis - на 5 биоваров. Бруцеллы - возбудители зоонозной инфекции человека и животных - бруцеллеза.
Возбудитель легко диссоциирует, переходя из S- в R- форму.
Культуральные свойства. Лучше растут на обогащенных средах сложного состава с добавлением крови или сыворотки крови, глюкозы, глицерина. Используют печеночный агар Хеддльсона, кровяной агар, мясо - пептонный бульон. Колонии возбудителя в S- форме мелкие, выпуклые, гладкие, с перламутровым оттенком, при диссоциации образуют широховатые R- формы колоний. Характерен медленный рост бруцелл в первых генерациях - колонии образуются через 2-4 недели. Рост бруцелл на жидких средах сопровождается равномерным помутнением сред.
Антигенная структура. Бруцеллы имеют общий соматический родоспецифический антиген, поэтому бруцеллы разных видов дают перекрестную агглютинацию. Два главных поверхностных антигена - А (преобладает у B.abortus) и М (преобладает у B.melitensis) встречаются в различных количественных соотношениях у различных видов бруцелл. Для их идентификации используют соответствующие антисыворотки.
Бруцеллы имеют поверхностный L- антиген (сходен с Vi- антигенами сальмонелл). Шероховатые формы содержат специфический R- антиген, для его идентификации используют специфические антисыворотки, применяемые при серотипировании. Колонии B.canis, B.ovis и B.suis 5 биотипа всегда имеют R- форму. Многие антигенные фракции бруцелл обладают выраженным аллергизирующим действием. У бруцелл имеются перекрестнореагирующие антигены с возбудителем туляремии,
Bordetella bronchiseptica и Y.enterocolitica серотипа 09.
Биохимические свойства. Бруцеллы ферментируют углеводы, однако при дифференциации на виды и биотипы используют ряд дополнительных признаков, в т.ч. способность расти на средах в присутствии обладающих бактериостатическим действием на отдельные виды бруцелл красителей (основной фуксин, тионин, сафранин), выделять сероводород, образовывать ферменты (уреазу, фосфатазу, каталазу), окислять различные аминокислоты.
Эпидемиологические особенности. Бруцеллез - зооноз преимущественно сельскохозяйственных и домашних животных. Человек инфицируется от животных или при контакте с инфицированным сырьем животного происхождения. Бруцеллез может носить профессиональный характер (уход за инфицированными животными) или быть связан с употреблением недостаточно термически обработанных молочных или мясных продуктов. Возбудитель может внедряться в организм человека через поврежденную кожу, слизистые дыхательных путей (аэрогенно) и желудочно - кишечного тракта (алиментарным путем), при заносе возбудителя на конъюнктиву глаза. Пути заражения - контактный, алиментарный и аспирационный. Бруцеллы обладают относительно высокой устойчивостью во внешней среде.
Патогенез и факторы патогенности. Патогенность бруцелл связана с наличием эндотоксина, гиалуронидазы и других ферментов, наличием низкомолекулярных продуктов, способствующих подавлению фагоцитоза и окислительного взрыва в макрофагах, наличием аллергизирующих субстанций.
По лимфатическим путям бруцеллы попадают в регионарные лимфоузлы, где размножаются в макрофагах, частично подвергаются внутриклеточному уничтожению. Продукты жизнедеятельности бруцелл ингибируют фагосомо - лизосомальное слияние. Морфологически в лимфоузлах формируется “первичный бруцеллезный комплекс” с формированием гранулем Новицкого с преобладанием крупных эпителиоидных клеток. Из разрушенных макрофагов в лимфоузлах бруцеллы попадают в кровь, распространяются по организму и поражают лимфатическую систему, печень, селезенку, нарушаются функции опорно - двигательного аппарата, нервной и половой систем.
Болезнь может иметь длительное хроническое течение (по сути - хрониосепсис). В патогенезе бруцеллеза имеет значение цикличность процессов, связанных с повторными проникновениями бруцелл в кровь из очагов с развитием местной воспалительной и общей реакций, на формирование которых существенное влияние оказывает специфическая сенсибилизация (реакция ГЗТ). Существенное значение в патогенезе, кроме аллергической перестройки и интоксикации, имеет преимущественное вовлечение в инфекционный процесс тех или иных органов и тканей.
Лабораторная диагностика осуществляется с использованием бактериологических методов, биопробы, серологических реакций, аллергической пробы Бюрне, генетических методов.
Материалом для бактериологических исследований служат кровь, костный мозг, грудное молоко (у кормящих), моча, околосуставная жидкость. Чаще выделяют гемо- и миелокультуры. Чистые культуры изучают по биохимическим свойствам, определяют их способность расти на средах с добавлением бактериостатических красителей, типируют в РА.
На практике чаще используют различные серологические методы диагностики:
- развернутую реакцию агглютинации Райта - основной метод и ускоренные микрометоды на стекле - агглютинации (Хеддельсона), роз- бенгал, латекс - агглютинации;
- реакцию Кумбса - для выявления неполных антител с применением антиглобулиновой сыворотки;
- РНИФ, РПГА с антигенным эритроцитарным диагностикумом, ИФА.
При отрицательных результатах бактериологических и серологических исследований применяют аллергическую кожную пробу с бруцеллином (пробу Бюрне).
Для выделения бруцелл из объектов внешней среды и от животных широко используют биопробу, для серодиагностики у животных - РСК, для исследования молока - кольцевую пробу.
Специфическая профилактика. В очагах козье - овечьего бруцеллеза применяют живую бруцеллезную вакцину ЖБВ. Разработана химическая бруцеллезная вакцина, которая отличается от живой вакцины более низкой реактогенностью.
Лекция N 10. Франциселлы и иерсинии
Род Francisella
Представители этого рода входят в семейство Brucellacea.Основной вид - F.tularensis - возбудитель туляремии - природноочаговой инфекции, резервуаром которой являются многие виды преимущественно мелких диких позвоночных животных (представителей четырех основных семейств - мышевидных, заячьих, беличьих и тушканчиковых). На территории России основными носителями являются мышевидные грызуны - водяные крысы, ондатры, различные виды полевок. Помимо F. tularensis к этому роду относится F. novicida, патогенность которой для человека не доказана.
Морфология.
Франциселлы - мелкие кокковидные или элипсоидные полиморфные палочки, неподвижные, грамотрицательные, не образующие спор.
Культуральные свойства.
Строгие аэробы, оптимум температуры около +37 градусов Цельсия, pH- близкая к нейтральной. Культивируют на агаровых и желточных средах сложного состава с добавлением цистеина, глюкозы, крови. Рост медленный. Образуют мелкие колонии, напоминающие капельки росы, круглые с ровным краем, выпуклые, блестящие, с голубоватым отливом.
Биохимические свойства.
Слабо ферментируют до кислоты без газа некоторые углеводы (глюкозу, мальтозу, левулезу, маннозу), образуют сероводород. Туляремийный микроб по вирулентности для кроликов и биохимическим особенностям, а также географическому распространению разделен на подвиды (эколого - географические расы):
- голарктическую (не ферментирует глицерин, цитруллин, маловирулентен для кроликов и человека, распространен в Евразии и Америке); туляремийный микроб этого подвида более приспособлен к водным экосистемам, способен распространяться водным путем, передаваться через комаров;
- неарктическую (ферментирует глицерин, не ферментирует цитруллин, более вирулентен для кроликов и человека, распространен в Северной Америке);
- среднеазиатскую (ферментирует глицерин и цитруллин, мало вирулентен). Среднеазиатский подвид по свойствам занимает промежуточное положение между первыми двумя, приближаясь к исходной предковой форме возбудителя.
Голарктический подвид включает три биовара - 1 (Erys - эритромицинчувствительный), 2 (Eryr - резистентный) и японский (var.japonica). Биовар 2 известен только в Евразии, где совпадает с распространением водяной крысы.
Антигенные свойства.
F.tularensis в S (вирулентной) форме имеет два основных антигенных комплекса - О антиген (обнаруживает сходство с О - антигенами бруцелл) и Vi (капсульный) антиген. Диссоциация S→ R приводит к утрате капсулы, вирулентности и иммуногенности.
Эколого - эпидемиологические особенности.
На территории России выделено 7 основных ландшафтных типов природных очагов туляремии : пойменно - болотный, луго - полевой, степной, лесной, предгорно - ручьевой, тундровый и тугайный (пойменно - пустынный) со своими основными хозяевами возбудителя и эколого - эпидемиологическими особенностями. Человек очень чувствителен к туляремийному микробу, минимальная инфицирующая доза - одна микробная клетка. Животные по чувствительности к этому микроорганизму разделены на четыре группы. Особое значение в условиях Западной Сибири имеют водяные крысы и ондатры. Заражение человека может происходить путем контакта с грызунами или инфицированными ими предметами, алиментарным путем (инфицированные грызунами вода и пищевые продукты), воздушно - пылевым путем, трансмиссивно (иксодовые клещи и другие кровососы). Н.Г.Олсуфьев выделяет две экологические формы возбудителя - “сухопутную”, характеризующуюся передачей через иксодовых клещей (все три подвида), и “водную”, связанную с околоводными видами грызунов и другими организмами - гидробионтами, с преимущественной передачей через воду и укусы комаров (голарктический подвид).
Клинико - патогенетические особенности.
Возбудитель туляремии является внутриклеточным паразитом. Его вирулентность обусловлена :
- капсулой, угнетающей фагоцитоз;
- нейраминидазой, способствующей адгезии;
- эндотоксином (интоксикация);
- аллергенными свойствами клеточной стенки;
- способностью размножаться в фагоцитах и подавлять их киллерный эффект;
- наличием рецепторов к Fc- фрагменту IgG, подавлять активность систем комплемента и макрофагов.
Франциселлы проникают в организм через кожу и слизистые глаз, рта, дыхательных путей, желудочно - кишечного тракта. Г.П.Руднев (1970) предложил в патогенезе туляремии выделить следующие стадии:
1. Внедрения и первичной адаптации возбудителя.
2. Лимфогенного заноса.
3. Первичных регионарно - очаговых (туляремийный бубон) и общих реакций.
4. Гематогенных матастазов и генерализации.
5. Вторичной полиочаговости.
6. Реактивно - аллергических изменений.
7. Обратного метаморфоза и выздоровления.
В ряде случаев процесс может ограничиваться первыми тремя фазами.
Основными клиническими формами туляремии являются язвенно - бубонная (ульцерогландулярная), глазо - бубонная (окулогландулярная), легочная, абдоминальная, генерализованная, другие формы (в т.ч. ангинозно - гландулярная), неуточненная (международная статистическая классификация болезней, 10 пересмотр. ВОЗ,
1995). Лабораторная диагностика.
Бактериологические методы диагностики туляремии для человека имеют дополнительное значение и не всегда эффективны, что определяется биологическими особенностями возбудителя и особенностями инфекции у человека (малая концентрация возбудителя в органах и тканях).
Биопроба является намного более эффективным методом диагностики. Материал от больного (пунктат бубона, выделения с конъюнктивы, пленка с миндалин, мокрота и др.) используют для заражения лабораторных животных (чаще белых мышей), из органов павших животных делают высевы на питательные среды, культуру идентифицируют по совокупности следующих признаков:
а) морфология клеток и грамотрицательная окраска ;
б) рост на желточной среде и специальных средах и отсутствие роста - на простых
мясо - пептонных средах;
в) специфическое свечение в реакции иммунофлюоресценции (МФА);
г) агглютинация культуры туляремийной сывороткой;
д) способность вызывать гибель белых мышей и морских свинок с характерными
патологоанатомическими изменениями в органах и выделением чистой культуры.
Бактериологические методы и биопробы могут выполняться только специализированными лабораториями, имеющими разрешение на работу с возбудителем туляремии (2 группа патогенности). В качестве метода выявления туляремийного микроба может использоваться МФА, реакция нейтрализация антител - РНАТ, в качестве дополнительного - ПЦР.
Наибольшее значение в лабораторной диагностике туляремии имеют серологические методы - РА, РПГА. Обязательно исследование парных сывороток крови. Дополнительными серологическими методами являются ИФА, РНИФ.
Аллергодиагностика (проба с тулярином - туляремийным аллергеном) чаще используется для оценки естественного и вакцинального иммунитета. ГЗТ развивается на первой неделе болезни, а также после вакцинации и сохраняется несколько лет. У больных накожные и внутрикожные туляриновые пробы не рекомендуются в связи с возможностью ухудшения состояния больного. Могут применяться методы аллергодиагностики in vitro - реакция лейкоцитолиза, РТМЛ и др.
Специфическая профилактика.
На неблагополучных по туляремии территориях применяют живую туляремийную вакцину. Иммунитет длительный, проверяется с помощью пробы с тулярином. С помощью этой пробы отбирают контингенты на вакцинацию и ревакцинацию.
Род Yersinia.
Род включает 11 видов. Y.pestis вызывает чуму, Y.pseudotuberculesis - псевдотуберкулез, Y.enterocolitica - (кишечный) иерсиниоз, ряд видов не патогенны или условно- патогенны для человека.
Морфология.
Чаще имеют овоидную (кокко-бациллярную) форму, окрашиваются биполярно, склонны к полиморфизму. Большинство видов подвижны при температуре ниже +30 градусов Цельсия (имеют перитрихиальные жгутики), грамотрицательны, имеют капсульное вещество. Y.pestis неподвижны, имеют капсулу.
Культуральные и биохимические свойства.
Факультативные анаэробы. Температурный оптимум от +25 до + 28 градусов Цельсия, pH - близкая к нейтральной. Хорошо культивируются на простых питательных средах. Ферментируют большинство углеводов без образования газа. Иерсинии способны менять свой метаболизм в зависимости от температуры и размножаться при низких температурах (психрофильные свойства). Вирулентные штаммы образуют шероховатые (R) колонии, переходные (RS) и сероватые слизистые гладкие (S) формы.
При изучении колоний чумного микроба выделяют два типа колоний - молодые и зрелые. Молодые микроколонии в неровными краями (стадия “битого стекла”) в дальнейшем сливаются, образуя нежные плоские образования с фестончатыми краями (стадия “кружевных платочков”). Зрелые колонии крупные, с бурым зернистым центром и неровными краями (“ромашки”). Многие штаммы способны восстанавливать красители с обесцвечиванием сред (метиленовый синий, индиго и др.). На скошенном агаре черед двое суток при +28 С образуют серовато - белый налет, врастающий в среду, на бульоне - нежную поверхностную пленку и хлопклвидный осадок. Температура +37С - селективная для образования капсулы у Y.pestis.
Культуры Y.pseudotuberculosis и Y.enterocolitica не имеют стадии “битого стекла”, вначале мелкие блестящие, выпуклые, затем может отмечаться сливной рост с образованием выпуклых бугристых колоний, схожих с колониями Y.pestis. Растут на универсальных питательных средах (среда Эндо, агар Мак Конки, среда Серова и др.) в сочетании с методами накопления в холодовых условиях.
Антигенная структура.
Все виды иерсиний имеют О - антиген (эндотоксин), схожий с О - антигенами других грамотрицательных бактерий и токсичный для человека и животных. Липополисахаридно- белковые комплексы О - антигенов иерсиний разделяют на S (гладкие) и R (шероховатые), последние - общие для Y.pestis и Y.pseudotuberculosis. Y.enterocolitica имеет поверхностный антиген, общий с другими энтеробактериями.
Возбудитель псевдотуберкулеза по О - и Н- антигенам подразделены на 13 сероваров, чаще встречаются серовар I, а также III и IV, иерсиниоза - на 34 серовара по О - антигену, чаще от человека выделяют серовары О3 и О9. При температуре от +22 до +25С Y.pseudotuberculosis и Y.enterocolitica имеют жгутиковый антиген и подвижны, при +37C теряют Н - антиген и подвижность.
Y.pestis более однородна в антигенном отношении, имея капсульный антиген (фракция I), антигены T, V - W, белки плазмокоагулазы, фибринолизина, наружной мембраны и др. Чумной микроб выделяет бактериоцины (пестицины), оказывающие бактерицидное действие на псевдотуберкулезный микроб и штаммы кишечной палочки.
Патогенные свойства.
Возбудитель чумы обладает наибольшим патогенным потенциалом среди бактерий. Он подавляет функции фагоцитарной системы, поскольку подавляет окислительный взрыв в фагоцитах и беспрепятственно в них размножается. Факторы патогенности контролируются плазмидами трех классов. В патогенезе выделяют три основных стадии - лимфогенного заноса, бактеремии, генерализованной септицемии.
Возбудители псевдотуберкулеза и иерсиниоза имеют адгезины и инвазины, низкомолекулярные протеины (ингибируют бактерицидные факторы), энтеротоксин.
Часть факторов контролируется плазмидами вирулентности.
Клинические особенности.
Чума чаще протекает в бубонной, легочной и кишечной формах. Наиболее опасны больные легочной чумой, которые выделяют с мокротой огромное количество возбудителя).
Иерсиниоз и псевдотуберкулез - кишечные инфекции. Клиника многообразна - региональная лимфоаденопатия (имитирует аппендицит), энтероколиты, реактивные артриты, анкилозирующий спондилит, скарлатиноподобная лихорадка.
Эпидемиологические особенности.
Чума - классический природноочаговый зооноз диких животных. Основные носители в природе - сурки, суслики, песчанки, пищухи, в антропургических (городских) условиях - крысы (чума портовых городов). В передаче возбудителя, особенно в очагах, где преобладают незимноспящие животные, имеют блохи животных, способные нападать и заражать человека. В песчаночных очагах могут заражаться верблюды и представлять эпидемическую опасность.
Псевдотуберкулез и кишечный иерсиниоз в природе передаются грызунами. Способны длительно сохраняться и даже накапливаться при низких температурах, например, в овощехранилищах. Способны вызывать заболевания у сельскохозяйственных животных. Человеку передаются преимущественно с пищевыми продуктами от животных, а также растительного происхождения.
Лабораторная диагностика.
Бактериологической диагностикой чумы могут заниматься только специализированные лаборатории противочумных станций и институтов (1 группа патогенности). Методами экспресс - выявления антигена являются МФА, РПГА с эритроцитарным дагностикумом, сенсибилизированным моноклональными антителами к капсульному антигену, ИФА, РНАТ. Для серологической диагностики может использоваться ИФА, РНАГ, ИФА.
При бактериологической диагностике кишечного иерсиниоза и псевдотуберкулеза в связи с накоплением возбудителя при низких температурах ( в отличии от большинства других микроорганизмов) материал предварительно забирают в забуференный физиологический раствор и сохраняют в холодильнике с периодическими высевами на среды Эндо, Плоскирева, Серова. Подозрительные колонии пересевают для получения чистых культур, изучают их по биохимическим свойствам и идентифицируют в РА с диагностическими сыворотками.
Для серологической диагностики используют РА и РНГА ( на псевдотуберкулез - с I сероваром, на иерсиниоз - с сероварами О3 и О9) с исследованием взятых в динамике инфекционного процесса парных сывороток.
Специфическая профилактика.
Применяется в очагах чумы. Используется живая ослабленная вакцина из штамма EV. Имеется сухая таблетированная вакцина для перорального применения. Для оценки иммунитета к чуме (естественного постинфекционного и вакцинального) может применяться внутрикожная аллергическая проба с пестином.
Лекция N11. Спирохеты (трепонемы, боррелии, лептоспиры).
Спирохеты - тонкие, спирально завитые бактерии длиной от нескольких до нескольких сотен микрометров, подвижные, грамотрицательные, хемоорганотрофы. Выделяют три основных типа движений - быстрое вращение вокруг продольной оси, сгибательные, штопорообразные (винтообразные). Медицинское значение имеют представители родов Treponema, Borrelia, Leptospira и Spirillum.
Род Treponema.
Род представлен туго закрученными спиралевидными бактериями длиной 5 - 20 мкм. Наиболее известный патогенный для человека вид - T. pallidum (бледная трепонема), подвид T. pallidum pallidum - возбудитель сифилиса.
Патогенные трепонемы дифференцируют по патогенности для различных видов лабораторных животных, способности ферментировать маннит, утилизовать лактат и образовывать специфические метаболиты.
“Lues Venerae” - “любовная чума” - одно из названий сифилиса (люес).
Морфология.
Возбудитель сифилиса имеет спиралевидную форму с одинаковыми по высоте завитками (более 10). Характер подвижности - плавные винтообразные и сгибательные движения (Treponema -с лат. - сгибающаяся нить). По Романовскому - Гимзе окрашиваются в бледно - розовый цвет (pallidum - с лат.- бледная). Легко выявить при помощи темнопольной микроскопии и после инпрегнации серебром.
Культуральные и биохимические свойства.
Бледная трепонема чрезвычайно прихотлива к условиям культивирования и не способна длительно расти на питательных средах. Отличается очень медленным темпом размножения. В лабораторных условиях можно культивировать на кроликах, заражая их интратестикулярно (орхит с накоплением трепонем). Биохимические свойства изучены слабо в связи с проблемами культивирования и для характеристики T.pallidum практически не используются.
Антигенные характеристики.
Трепонемы имеют перекрестно - реагирующие антигены с другими спирохетами (боррелиями, лептоспирами). В связи с наличием в составе клеточных стенок бледных трепонем фосфолипидов (кардиолипин и др.), аналогичных клеткам млекопитающих, в ответ на эти антигены в организме вырабатываются специфичные не только для возбудителя сифилиса, но и перекрестно- реагирующие на кардиолипин и другие фосфолипиды тканей человека и животных так называемые вассермановские антитела. Они выявляются в РСК с кардиолипиновым антигеном (из сердец животных). Специфические антитела к возбудителю сифилиса выявляют в РНИФ и иммуноблоте.
Антиген Вассермана - фосфолипид, входящий в состав митохондриальных мембран (кардиолипин). Его получают из бычьего миокарда - ткани, богатой митохондриями. Благодаря антигенной общности с тканевыми фосфолипидами, антитела к кардиолипину трепонем реагируют с тканевым (митохондриальным) кардиолипином. Истинные аутоантитела против тканевого кардиолипина образуются при “антимитохондриальном (антифосфолипидном) синдроме”, обусловливая положительные реакции Вассермана (ложноположительные - применительно к возбудителю сифилиса) при коллагенозах (системная красная волчанка, ревматоидный артрит), проказе и других тяжелых инфекциях, связанных с повреждением тканей, при гемолитической анемии, при наркоманиях.
Патогенетические особенности.
Развитие сифилиса определяется двумя основными механизмами - склонностью к генерализации (высокая инвазивность) и периодической активизацией длительно персистирующего в организме возбудителя (“выход из засады”).
Из-за неустойчивости возбудителя во внешней среде заражение происходит при прямых контактах ( в подавляющем большинстве случаев половым путем). Проникая через слизистые оболочки и незначительные повреждения кожи, возбудитель быстро проникает в ткани, распространяется лимфогенно и далее - гематогенно (генерализация).
Первые признаки болезни (первичный сифилис) возникает после инкубационного периода (в среднем 3 - 4 недели). В месте внедрения (первичной инвазии) происходит размножение возбудителя и возникает твердый шанкр - плотный безболезненный красноватый узелок, превращающияся в язву. Одновременно в результате проникновения трепонем в лимфатические узлы развивается регионарный лимфаденит и в дальнейшем - полиаденит. Генерализация инфекции происходит еще до появления шанкра и лимфаденита, т.е. сифилис с самого начала - системное заболевание.
После затихания клиники первичного сифилиса через некоторое время ( от нескольких недель до нескольких месяцев) происходит рецидив инфекции, связанный с полиорганной репликацией трепонем (вторичный сифилис). Наблюдаются поражения кожи и слизистых (вторичные сифилиды, в которых находится большое количество трепонем), поражения внутренних органов, лимфоузлов, костей, суставов и мн.др. С учетом многообразия поражений вторичный сифилис называют “великим мистификатором”.
После вторичного сифилиса у части больных вновь формируется латентный сифилис (возбудитель уходит в подполье на годы), который у половины носителей может переходить в третичный сифилис. Он характеризуется формированием очагов гранулематозного воспаления - гумм, содержащих мало трепонем (практически не заразных). Распад гумм приводит к разрушению костных и мягких тканей и поражению жизненно важных органов. В отличие от первичного и вторичного сифилиса, третичный сифилис - хронический процесс, который никогда не завершается самоизлечением.
Мало известно о факторах патогенности T.pallidum и механизмах, позволяющих ему длительно ускользать от иммунологического надзора. Обнадеживает,что этот возбудитель сохраняет высокую чувствительность к антибиотикам. Современный сифилис по- прежнему хорошо поддается лечению, если процесс не достигает необратимых органических поражений.
Лабораторная диагностика.
При первичном и вторичном сифилисе возбудитель можно обнаружить в очагах поражений микроскопией. Более оптимальны для выявления бледных трепонем темнопольная и иммунолюминесцентная микроскопия. При первом методе необходимо исключить возможные артефакты (нити фибрина, например), наличие сапрофитических трепонем. T.refringens колонизирует наружные половые органы, отличается высокой подвижностью и отсутствием сгибательных движений. T.denticola колонизирует ротовую полость, ее завитки короче и направлены под более острым углом. В ротовой полости человека встречается также T.vincentii, которая в ассоциации с другими микроорганизмами может вызывать язвенно - некротическую ангину.
Кроме возбудителя сифилиса имеются очень близкие в генетическом и антигенном отношении трепонемы других подвидов T.pallidum, вызывающие поражения человека в странах с жарким климатом (фрамбезию, бержель, пинту).
При окраске по Романовскому - Гимзе T.pallidum окрашивается в розовй цвет, непатогенные трепонемы - в синий или фиолетовый, T.refringens прокрашивается фуксином в красный цвет. Люминесцентная диагностика широко применяется для выявления трепонем в различных клинических материалах. Среди методов выявления возбудителя наибольшей чувствительностью и достаточной специфичностью обладает ПЦР.
Основные методы диагностики - серологические, т.е. выявление сывороточных антител. Применяют РСК с кардиолипиновым и трепонемным антигеном, флоккуляционные пробы (микроагглютинационный тест на кардиолипиновые антитела), РНГА. Более специфичными и чувствительными тестами являются тесты, основанные на непрямой иммунофлюоресценции (РИФ), реакция иммобилизации бледной трепонемы, ИФА на основе рекомбинантных белков трепонем.
В серодигностике сифилиса необходимо учитывать серонегативность первичного сифилиса в первые недели заболевания. Необходимо сочетание методов , основанных на выявлении антител как против кардиолипиновых, так и трепонемных антигенов. Кардиолипиновые тесты быстро становятся серонегативными после элиминации возбудителя (спустя несколько месяцев), антитрепонемные антитела сохраняются намного дольше. Существенное значение для определения активности процесса имеет определение антитрепонемных IgM - антител (при выявлении врожденного сифилиса -особо, поскольку IgM - антитела не переходят через плаценту от матери к плоду; при исследовании спинномозговой жидкости - для диагностики нейросифилиса и др.).
Лечение.
Чаще применяют депонированные пенициллины с пролонгированным действием, при непереносимости - макролиды, тетрациклины, цефалоспорины. Специфической профилактики не существует.
Род Borrelia
Морфология и классификация.
Спиральные, имеющие до 10 неправильной формы крупных завитков, грамотрицательные бактерии с вращательно- поступательным характером движений. Анаэробы, часто требующие сложных сред для культивирования.
Патогенные для человека боррелии являются возбудителями возвратных тифов (рекуррентных лихорадок) или боррелиозов. B.recurrens передается человеку вшами, вызывает эпидемический или вшивый возвратный тиф. Остальные боррелиозы человека делят на две самостоятельные группы - аргасовые клещевые боррелиозы (АКБ), вызываемые более 12 видами боррелий, и иксодовые клещевые боррелиозы (ИКБ), вызываемые B.burgdorferi sensu stricto, B.garinii, B.afzelii и некоторыми другими.
АКБ связаны с аргасовыми клещами рода Ornithodorus, обитающими в тропических и субтропических регионах Африки, Азии, Америки, и характеризуются повторяющимися приступами лихорадки (как при малярии). ИКБ вызывают клещи рода Ixodes (группа I.ricinus / I.persulcatus), распространены преимущественно в лесной зоне в Евразии и Америке.
Культуральные свойства.
Представители этого рода взыскательны к условиям культивирования, особенно боррелии группы ИКБ. Для них необходимы факультативно - анаэробные условия, температура плюс 33 градуса Цельсия, специальные среды (BSK-2), содержащие среду 199, глюкозу, альбумин, цистеин, кроличью сыворотку, желатин и другие компоненты.
Антигенные свойства.
Имеют перекрестно- реагирующие антигены с другими спирохетами, родо - и видоспецифические антигены. Выделяют H- (жгутиковые) флагеллиновые антигены (обладают слабой специфичностью) и поверностные белковые антигены (OspA, OspC, более специфичны, используют для межвидовой и внутривидовой идентификации).
Патогенез поражений.
После присасывания клеща боррелии со слюной попадают в макроорганизм, размножаются во входных воротах инфекции, поражая кожу (эритема) и ближайшие лимфоузлы (фаза первичной адаптации). Преодолев кожный и лимфатический барьеры, боррелии попадают в кровь, вызывают спирохетемию, проявляющуюся общетоксическим синдромом (стадия первичной диссеминации). По мере прогрессирования процесса боррелии проникают через гематотканевые барьеры (в т.ч.- через гематоэнцефалитический барьер) и вызывают поражение различных органов и систем. В ряде случаев инфекция приобретает хронический характер, вызывая поражения нервной и сердечно - сосудистой систем, опорно - двигательного аппарата, вторичные поражения кожи и др. Существует недостаточно обоснованное на современном этапе мнение о связи преимущественных клинических проявлений с геновидовой принадлежностью боррелий ( B.garinii - преимущественно неврологические проявления, например).
Эпидемиология.
ИКБ - облигатно- трансмиссивные природноочаговые инфекции, распространенные преимущественно в умеренном климатическом поясе северного полушария, лесной ландшафтной зоне и связанные с присасыванием клещей рода Ixodes. Очаги ИКБ часто сопряжены с очагами клещевого энцефалита, поскольку имеют одних и тех же переносчиков в Евразии - клещей I.reticulatus (таежный клещ) и I.ricinus (европейский лесной клещ).
Лабораторная диагностика.
Боррелии можно выделить с использованием среды BSK2 у больных из очагов кожных поражений, из крови и спинномозговой жидкости (при менингиальных формах), при исследовании переносчиков (в т.ч. снятых с людей) и теплокровных хозяев (наибольшая высеваемость - из мочевого пузыря) в природных очагах.
Боррелии можно выявить в иксодовых клещах с помощью световой микроскопии (окраска по Романовскому - Гимзе), темнопольной и люминесцентной микроскопии, ПЦР.
Основной метод серологической диагностики - реакция непрямой иммунофлюоресценции (РНИФ) с корпускулярным антигеном B.afzelii, позволяющим выявлять антитела к боррелиям группы ИКБ.
Лечение.
Применяют предупредительную терапию (при положительных результатах исследования присосавшегося клеща) и лечение больных ИКБ тетрациклинами, пенициллинами и цефалоспоринами. Мер специфической профилактики не разработано.
Род Leptospira Морфология.
Спиральные грамотрицательные бактерии с прямолинейным и винтообразным движением, часто с изогнутыми концами (в виде крючков). Род включает свободоживущие и паразитические виды. Основной патогенный для человека и животных вид - L.interrogans.
Культуральные и биохимические свойства.
Хемоорганотрофы, аэробы. Лептоспиры - типичные гидрофилы, долго сохраняются во влажных субстратах, воде, влажной почве. Оптимум температуры от плюс 28 до 30 градусов Цельсия, рН 7,2 - 7,4. Культивируют преимущественно на жидких средах с добавлением сыворотки крови кролика (среда Терских).
Антигенные свойства.
Патогенные лептоспиры на основании антигенных свойств разделяют на серогруппы и серовары.
Эколого - эпидемиологические особенности.
Основными резервуарами инфекции являются дикие животные, преимущественно грызуны и насекомоядные (природные очаги, часто связанные с околоводными стациями), а также сельскохозяйственные и домашние животные (свиньи, крупный рогатый скот, собаки). У животных лептоспиры длительно сохраняются в почках и длительно выделяются во внешнюю среду с мочой. Человек заражается в природных очагах (чаще во время сельскохозяйственных работ) и в хозяйственных очагах (чаще купальные вспышки или профессионально обусловленные случаи лептоспирозов). Существенную роль в заражении имеют серые крысы и собаки. Существуют связи лептоспир определенных сероваров с отдельными видами животных
(например, L.canicola - собаки).
Патогенез поражений.
В организм лептоспиры попадают через слизистые или повреждения (микротравмы) кожи. Патогенные лептоспиры благодаря активной подвижности преодолевают защитные барьеры, проникают в кровь (лептоспиремия) и попадают в различные органы , преимущественно в почки и печень. Особое значение имеют поражения эндотелия капилляров с различными по выраженности геморрагиями вплоть до геморрагического синдрома, поражения печени с развитием желтухи, связанные с повреждениями гепатоцитов и гемолизом эритроцитов, поражения почек, прежде всего эпителия почечных канальцев, коркового и подкоркового вещества с развитием почечной недостаточности (анурия, уремия), менингиальные проявления.
Лабораторная диагностика.
Основным методом микроскопической диагностики является темнопольная микроскопия.
Выделение возбудителя проводят посевами на среде Терских, биопробой на золотистых хомячках. Исследуют мочу, кровь, спинномозговую жидкость, корковый слой почки.
Основным методом серологической диагностики является реация агглютинации- лизиса (РАЛ) с набором культур лептоспир основных серогрупп. Метод специфический, однако трудоемкий, поскольку необходимо поддерживать диагностический набор лептоспир, агглютинировать исследуемые сыворотки с лептоспирами всех основных серогрупп с последующей темнопольной микроскопией. При положительном результате наблюдается склеивание лептоспир в виде паучков или клубков с последующим лизисом. В настоящее время для серодиагностики применяют также ИФА.
Лечение.Применяют антибиотики (пенициллины, тетрациклины).
Лекция N 12. Хламидии, уреаплазмы, микоплазмы.
Род Chlamydia и род Chlamydophila.
Представители семейства Chlamydiaceae (хламидии) являются патогенными облигатными внутриклеточными бактериями, паразитирующими в чувствительных клетках теплокровных (млекопитающих, птиц, человека и др.).
Они близки по структуре и химическому составу к классическим бактериям. Для них характерно сохранение морфологической сущности на протяжении всего жизненного цикла, деление вегетативных форм, наличие клеточной стенки, содержание ДНК и РНК, энзиматическая активность, чувствительность к антибиотикам широкого спектра, наличие общего родоспецифического антигена.
В то же время хламидии по размерам меньше классических бактерий, имеют небольшой геном, являются облигатными внутриклеточными паразитами с уникальным циклом развития. Они не способны синтезировать высокоэнергетические соединения и обеспечивать собственные энергетические потребности ( энергозависимые паразиты), что и определяет их облигатный паразитизм.
С учетом своих особенностей хламидии занимают самостоятельное (особое) положение среди других микроорганизмов - прокариот. Для человека имеют значение преимущественно представители двух родов - Chlamydia и Chlamydophila.
Наибольшее значение имеют три вида.
Chlamydia trachomatis , различные серотипы которого вызывают трахому, венерическую лимфогранулему и наиболее распространенные урогенитальные хламидиозы. Chlamydophila psittaci вызывает орнитоз и зоонозные хламидиозы.
Chlamydophila pneumoniae вызывает антропонозные пневмонии, ОРЗ, с этим возбудителем связывают развитие некоторых форм бронхиальной астмы, атеросклероза.
Морфологические особенности.
Клеточный цикл развития хламидий имеет две основных формы - элементарные тельца (ЭТ) - инфекционная форма и ретикулярные тельца (РТ) - вегетативная форма. Сферические ЭТ значительно меньше размерами (менее 300 нм в диаметре), имеют более жестную электронно- плотную структуру, метаболически мало активны, адаптированы к кратковременному внеклеточному существованию.
Цикл развития хламидий осущесвляется в цитоплазматическом включении - фагосоме (вакуоле), куда ЭТ попадают путем стимулирования эндоцитоза. В процессе адсорбции и эндоцитоза участвуют термолабильные эффекторные белковые поверхностные антигены хламидий. ЭТ подавляют фагосомо - лизосомальное слияние и преобразуются при участии главного поверхностного протеина в РТ, которые обладают активным метаболизмом, более крупными размерами и активным бинарным делением. Цикл размножения заканчивается обратным переходом РТ в ЭТ, разрывом мембран включения и ограничивающих мембран клетки хозяина, выходом ЭТ из клеток, далее ЭТ инфицируют новые клетки. Тельца включений выявляются в клетках при помощи световой и иммунолюминесцентной микроскопии.
Кроме того, хламидии способны образовывать L - формы, персистентные формы. Культуральные свойства.
Хламидии не растут на питательных средах самого сложного состава (это сближает их по свойствам с риккетсиями и особенно - с вирусами), для их культивирования могут быть использованы лабораторные животные и куриные эмбрионы (биопроба) и особенно чувствительные линии клеток животных - чаще клетки McCoy (чаще с обработкой цитостатиками для повышения чувствительности), которые считались “золотым стандартом” диагностики.
Биохимические свойства.
Способны самостоятельно синтезировать нуклеиновые кислоты, некоторые белки и липиды. Являются энергетически зависимыми от хозяина эндопаразитами. Chlamydia trachomatis обладает гликоген - синтетазной системой (гликоген содержится и выявляется при определенных методах окраски во включениях), синтезирует предшественники фолиевой кислоты и соответственно биохимической активности чувствителен к сульфаниламидам.
Антигенная структура.
1. Выделяют поверхностный родоспецифический антиген (ЛПС), локализующийся на наружной мембране клеточной стенки. ЛПС имееи две антигенные детерминанты, одна из них специфична для всего рода, другая перекрестно реагирует с некоторыми другими грамотрицательными бактериями (сальмонеллы серовара minnesota), термостабилен.
2. МОМР - главный белок наружной мембраны несет функцию структурного белка и порина. Включает термолабильные белковые детерминанты , обладающие видо - , типо - и сероварной специфичностью.
Детерминанты патогенности.
1. Эндотоксин (липополисахарид), подобный эндотоксинам грамотрицательных бактерий.
2. Экзотоксины - термолабильные субстанции.
3. Антигены клеточной поверхности, подавляющие защитные реакции.
Факторы патогенности хламидий препятствуют фагосомо - лизосомальному слиянию в фагоцитах.
Краткая характеристика распространения.
Урогенитальные хламидиозы - наиболее распространенные формы хламидиозов. В крупных странах (США, Россия) ежегодно регистрируют несколько миллионов случаев, в мире - от 50 до 90 млн. случаев. Скудность начальных проявлений и тяжелые последствия урогенитальных хламидиозов, особенно у женщих (нарушения репродуктиной сферы, инфекционные осложнения), предъявляют особые требования к своевременной лабораторной диагностике. Хламидии сероваров D - K Chlamidia trachomatis, вызывающие урогенитальные хламидиозы, передаются от человека к человеку половым путем. Бессимптомное носительство наблюдается не менее чем у 5 % мужчин и 10% женщин. Отдельные серотипы этого возбудителя вызывают такие распространенные в прошлом заболевания как трахома (сопровождается поражениями конъюнктивы и прилегающих тканей глаза, часто приводит к катарактам и слепоте) и венерическая лимфогранулема (регистрируют преимущественно в слаборазвитых странах Азии, Африки и Латинской Америки с теплым климатом).
Орнитоз - хламидийная инфекция, вызываемая C.psittaci. Человек заражается от птиц - основных хозяев этого возбудителя воздушно - пылевым и воздушно - капельным путем. В условиях города основную опасность представляют голуби (от 20% до 100% инфицировано этим возбудителем, чаще с ними контактируют дети. В домашних условиях источником могут быть канарейки и особенно попугаи (вызывают наиболее тяжелую форму - пситтакоз). Орнитоз часто протекает как тяжелая интерстициальная пневмония. Кроме этого, серотипы этого возбудителя вызывают зоонозные хламидиозы (например, так называемый вирусный аборт овец, хламидиозы крупного рогатого скота и др.), при контакте с больными животными могут развиваться различные формы хламидиозов у людей. Клинико - эпидемиологические особенности зоонозных хламидиозов у людей изучены недостаточно.
Бронхопневмонии , вызываемые C.pneumoniae. Это антропонозные инфекции, передаваемые от человека к человеку, большая часть случаев протекает субклинически. Возникают поражения верхних дыхательных путнй с последующим развитием бронхопневмонии. Это распространенные инфекции (антитела к C.pneumoniae выявляют почти у половины взрослого населения), однако диагностируются до настоящего времени плохо. С этим возбудителем связывают развитие отдельных форм бронхиальной астмы и атеросклероза.
С учетом многообразия клинических проявлений и необходимостью дифференциации различных клинических форм хламидиозов (прежде всего генитальных и экстрагенитальных) особое значение приобретает лабораторная диагностика.
Лабораторная диагностика.
Золотой стандарт - метод культивирования в культурах клеток применяется очень редко в связи с трудоемкостью и длительностью культивирования, необходимостью работы с инфекционным материалом, этот метод по чувствительности уступает ПЦР, требует быстрой доставки материала для исследования.
Применяемые лабораторные методы можно разделить на две основные группы - методы выявления антител и методы выявления антигенов.
Методы выявления антител наиболее эффективны при генерализованных формах хламидиозов, сопровождающихся выработкой антител в высоких титрах (орнитоз), и мало эффективны при локальных (особенно хронических) формах (урогенитальные хламидиозы). Большинство методов выявляет антитела к группоспецифическому липополисахариду хламидий, что не позволяет определить вид хламидий. Среди используемых методов:
- РСК - достаточно специфичный, но мало чувствительный метод;
- РНГА - более эффективный метод для диагностики текущего инфекционного заболевания;
- ИФА - наиболее чувствительный метод серологической диагностики. Некоторые варианты тест - систем ИФА позволяют дифференцировать C.pneumoniae от хламидий других видов;
- РНИФ - обладает наибольшей степенью видоспецифичности.
Методы выявления возбудителя, его ДНК и антигенов.
1. Цитологические методы с окраской мазков по Романовскому - Гимзе и другими методами мало чувствительны и специфичны, имеют преимущественно историческое значение.
2. Метод флюоресцирующих антител (МФА) с моноклональными антителами (МКА) к группоспецифическому липополисахариду хламидий - наиболее распространенный, чувствительный и специфичный метод, позволяет выявлять локализацию возбудителя (урогенитальные мазки), морфологию (характер гранул, преобладание РТ или ЭТ). Метод требует высокой квалификации микроскописта и качества мазка - отпечатка ( достаточное количество клеток с учетом внутриклеточного расположения возбудителя).
3. ИФА для выявления антигена применяется относительно реже, требует большого количества материала (соскоб), связана с получением суспензии и опасностью инфицирования персонала.
4. ПЦР для выявления ДНК хламидий - наиболее чувствительный метод. Однако и при нем возможны ложноположительные (при недостаточной чистоте работы - при контаминации) и ложноотрицательные (наличие в пробах материала ингибиторов Tag - полимеразы) результаты.
Недостатки чувствительных методов выявления антигенов возбудителя - возможность получения положительных результатов даже через 1 - 1,5 месяца после излечения. Нужна полная замена эпителия слизистой, содержащего поверхностные антигены разрушенных хламидий, для получения отрицательных результатов. Особенно это относится к ИФА, ПЦР, для МФА - в меньшей степени (этот метод позволяет оценить морфологию включений хламидий).
Лечение и профилактика.
Хламидии - внутриклеточные паразиты. Применяют антибактериальные препараты, проникающие в клетки, чаще доксициклин или сумамед (азитромицин). Эффективному лечению часто препятствует одновременное наличие у больных гонококков и трихомонад (в трихомонадах хламидии могут находиться внутриклеточно). Эффективность современных методов лечения урогенитальных хламидиозов не превышает в идеале 98 - 99%, т.е. часть пациентов эффективно освободить от хламидий не удается, даже после нескольких циклов лечения. У этих больных часто развиваются дисбактериозы, присоединяется кандидоз, снижается резистентность к различным инфекционным агентам.
Эффективных методов специфической профилактики нет.
Род Mycoplasma и род Ureaplasma
Микоплазмы - самые мелкие прокариотические микроорганизмы, не имеющие клеточной стенки (это придает им сходство с L - формами бактерий) и способные к паразитированию на мембранах эукариотических клеток. Способность персистировать на мембранах клеток связана с наличием сходства структуры и состава цитоплазматической мембраны микоплазм с мембранами клеток эукариот и использовании микоплазмами их компонентов (прежде всего холестерина и фосфолипидов) для построения собственных структур. Микоплазмы имеют трехслойную цитоплазматическую мембрану, обеспечивающую целостность микробных клеток и частично замещающую в функциональном отношении отсутствующую клеточную стенку.
Современная систематика относит представителей семейства Mycoplasmatacea к классу Mollicutes (молликут - “мягкокожих”), объединяющему микоплазмы, уреаплазмы, ахолеплазмы (первые три рода молликут встречаются у человека, среди них встречаются патогенные и сапрофитические виды), спироплазмы и анаэроплазмы.
Основные характеристики молликут - отсутствие истинной клеточной стенки, мелкая кокковидная форма, паразитизм на мембранах эукариотических клеток, рост на плотных средах в виде мелких врастающих в агар колоний с приподнятым центром (“яичница - глазунья”), абсолютная резистентность к пенициллинам и другим действующим на синтез клеточной стенки антибиотикам. Характеризуются малым размером генома, низким содержанием Г+Ц в ДНК. Имеют от 104 до 113 нуклеотидов в 5S рРНК.
Представители рода микоплазм - паразиты и возбудители заболеваний (микоплазмозов) широкого круга млекопитающих, птиц, человека. Представители некоторых видов встречаются на растениях и в насекомых. Известно около 100 видов микоплазм, число новых видов продолжает увеличиваться. 12 видов встречаются у человека, из них наибольшее значение имеют M.pneumoniae (вызывает в основном пневмонии и ОРЗ), M.hominis, M.genitalum, M.fermentans (при вызываемых ими инфекциях преобладают урогенитальные поражения). В составе рода Ureaplasma выделено пять видов. Штаммы, выделенные от людей, составляют вид U.urealyticum, от КРС - U.diversum, от птиц - U.gallorale, от кошек - U.cati и U.felinum.
Морфология
Наиболее типичны мелкие кокковидные формы, выражен полиморфизм.
Культуральные и биохимические свойства
Микоплазмы прихотливы к условиям культивирования. В составе сред необходимы холестерин и стерины, нативные сыворотки, витамины, соли. Основным источником энергии являются углеводы (особенно глюкоза) или аминокислоты (аргинин). Для усиления восстановительных свойств сред добавляют L- цистеин.
Используют различные варианты жидких, полужидких и плотных питательных сред, включающих гидролизаты мяса, казеина, ферментативный пептон, аутолизат пекарских дрожжей, сыворотку крови лошадей (больше всего холестерина), основной субстрат - глюкозу (для глюкозоферментирующих микоплазм), аргинин (для аргинин- утилизирующих микоплазм), мочевину (для уреаплазм), селективные антибиотики (пенициллин и его синтетические аналоги, амфотеррицин, нистатин или низорал, для уреаплазм - линкомицин), индикатор рН (бромтимоловый синий или феноловый красный).
Оптимум температуры для культивирования от + 35 до +37С. Оптимум рН для уреаплазм и аргинин- утилизирующих микоплазм - 6,0 - 6,5. При росте уреаплазм, обладающих ферментом уреазой, происходит расщепление мочевины до аммиака с повышением рН (сдвиг в щелочную сторону) и с изменением цвета среды с бромтимоловым синим из желто - лимонного до зеленого без образования значительного осадка и без помутнения среды. Аналогично происходит изменение цвета сред при росте аргинин- утилизующих микоплазм на средах с добавлением аргинина.
Оптимум рН для глюкозоверментирующих микоплазм (M.genitalium, M.fermentans, M.pneumoniae) - около 8,0. Эти микоплазмы ферментируют глюкозу до углекислого газа со сдвигом рН в кислую сторону и изменением цвета среды с зеленого на желто - лимонный.
Колонии уреаплазм (ранее называли Т- микоплазмами от “tiny -маленький”) значительно меньше колоний классических микоплазм, на средах с добавлением сульфата марганца их колонии окрашиваются в золотисто - коричневый цвет, в отличии от микоплазм уреаплазмы мало чувствительны к небольшим дозам линкомицина (15 мкг/ мл).
Антигенная структура
Микоплазмы характеризуются выраженной гетерогенностью и изменчивостью антигенной структуры (антигенный полиморфизм). Известно 16 серотипов U.urealyticum, разделенных на серогруппы А и В. Степень гомологии серотипов существенно отличается. Часто выделяют от больных смешанные культуры различных серотипов. Выделяют биовары уреаплазм, отличающиеся по вирулентности и строению гена основного фермента - уреазы. Аналогичная динамичность и гетерогенность антигенной структуры и наличие различных сероваров характерна и для различных видов классических микоплазм. По биохимическим и в определенной мере антигенным свойствам среди микоплазм имеются виды - двойники (например, M.pneumoniae и M.genitalium). Среди антигенов выделяют белки - адгезины, фосфо - и гликолипиды, полисахаридные компоненты.
Факторы патогенности
1. Адгезины (обеспечивают взаимодействие с клетками хозяина).
2. Эндотоксины (не тождественны ЛПС грамотрицательных бактерий).
3. Гемолизины (особенно - у M.pneumoniae).
4. Экзотоксины (плохо изучены и известны у немногих видов).
5. Ферменты - фосфолипаза А, аминопептидазы, протеазы, нейраминидаза. Патогенез.
Микоплазмы - мембранные паразиты и способны адсорбироваться на различных эукариотических клетках. Этому способствует подвижность патогенных микоплазм, связанная с хемотаксисом и наличием микроворсинок. Микоплазмы способны тесно связываться с мембранами эукариотической клетки, вступать в межмембранное взаимодействие, обмениваться мембранными компонентами (в том числе использовать холестерин и стенолы для построения собственных клеток), оказывать разнообразное влияние на клетки. Коротко остановимся на основных эффектах действия микоплазм на различные клетки.
1. Взаимодействие с фагоцитирующими клетками вызывает цитопатический или цитотоксический эффект и гибель фагоцитов или приводит к длительной персистенции микоплазм в фагосомах фагоцитов.
2. Воздействие микоплазм на макрофаги приводит к нарушению их функций.
3. Персистенция микоплазм на мембранах лимфоидных клеток оказывает существенное влияние на их функции, в том числе непосредственное деструктивное воздействие на иммунокомпетентные клетки (M.fermentans оказывает СПИДо - подобное действие на иммунную систему).
4. Микоплазмы действуют на эритропоэз, т.е. на гомеостаз организма.
5. Обмен антигенными компонентами мембран с клетками хозяина обеспечивает антигенную мимикрию и развитие аутоиммунных реакций.
6. Отсутствие клеточной стенки и частоя локализация микоплазм в инвагинатах мембран клеток объясняет их слабую иммуногенность и защиту от действия антител. Этому способствует и высокая антигенная изменчивость молликут.
7. Микоплазмы способны воздействовать на хромосомный аппарат клеток хозяина (на соматические и половые клетки), вызывая хромосомные абберации.
8. Ряд продуктов метаболизма оказывает токсическое воздействие на инфицированные клетки. Так, глюкозоферментирующие микоплазмы резко снижают рН и оказывают этим деструктивное действие на эпителиальные клетки.
Частота распространения отдельных видов микоплазм.
Среди урогенитальных молликут на первом месте уреаплазмы, на втором - M.hominis, они по частоте встречаемости уступают только хламидиям. Большинство микоплазм - возбудители оппортунистических инфекций, наиболее часто их выявляют у категорий риска (больные с воспалительными процессами гениталий, часто меняющие сексуальных партнеров и др.), у беременных. Наибольший патогенный потенциал признают за M.pneumoniae, M.fermentans, U.urealyticum, M.hominis. M.pneumoniae - один из распространенных возбудителей, вызывающих пневмонии и другие инфекционные заболевания дыхательных путей человека. Большинство выделяемых от животных микоплазм не имеет медицинского значения. Лабораторная диагностика
Наиболее оптимальна культуральная (бактериологическая) диагностика с использованием жидких и плотных питательных сред, особенно в сочетании с антибиотикограммой и определением титра микоплазм (уреаплазм) в нативных образцах. Преимущества этого подхода: быстрота (для уреаплазм 1-2 суток, для микоплазм - до 3 суток), простота (цветной пробирочный тест), достоверность (селективность сред, учет биохимизма возбудителей), возможность определения конценрации (титр 10000 цветообразующих единиц - ЦОЭ имеет диагностическое значение) и чувствительности к различным препаратам на основе антибиотикограммы (повышает эффективность лечения), возможность достоверного контроля эффективности лечения (растут только жизнеспособные микоплазмы).
Наиболее чувствительна ПЦР - диагностика. Однако: возможны ложно - положительные и ложно - отрицательные результаты, длительность выявления положительных результатов контроля излеченности, невозможность определения титра микоплазм (не разработаны количественные методики ) и чувствительности к препаратам.
Люминесцентная диагностика дает неудовлетворительные результаты чаще, чем при выявлении хламидий. Это связано с малыми линейными размерами возбудителей, серологической гетерогенностью и антигенной изменчивостью, возможностью перекрестных реакций между различными видами, наличием диагностических препаратов только для выявления уреаплазм и M.hominis.
Серологические методы многочисленны, однако коммерческие препараты отсутствуют, существуют проблемы в интерпретации результатов.
Лечение должно быть основано на подборе чувствительных антибиотиков (чаще применяют макролиды, тетрациклины - доксициклин) и бактериологическом контроле эффективности лечения. Имеются устойчивые к тетрациклинам, макролидам и другим антибиотикам штаммы (связано с наличием плазмид устойчивости). У микоплазм имеются специфические вирусы, в т.ч. бактериофаги. В лечебных и диагностических целях они до настоящего времени не используются.
Специфическая профилактика не разработана. Иммунитет нестойкий, серотипоспецифический.
Лекция № 13 Миксовирусы
Известно 13 семейств РНК- геномных вирусов, имеющих медицинское значение - пикорнавирусы, калицивирусы, реовирусы, ретровирусы, тогавирусы, флавивирусы, буньявирусы, аренавирусы, филовирусы, рабдовирусы, коронавирусы, ортомиксовирусы, парамиксовирусы.
Возбудителями острых респираторных заболеваний (ОРЗ) являются пяти семейств РНК - содержащих и одно семейство ДНК - содержащих вирусов. Особое место среди них занимают представители семейства Orthomyxoviridae (ортомиксовирусы - вирусы гриппа А, В и С), а также большая группа парамиксовирусов (семейство Paramyxoviridae), включающих род Paramyxovirus (вирусы парагриппа человека 1,2,3,4 типов, болезни Ньюкасл, парагриппа птиц, паротита), род Pneumovirus (RS или респираторно - синтициальный вирус), род Morbillivirus (вирус кори). Ортомиксовирусы - вирусы гриппа А, В, С Структурные особенности.
Ортомиксовирусы являются оболочечными (суперкапсидными, “одетыми”) вирусами, средний размер вирионов - от 80 до 120 нм. Вирионы имеют сферическую форму. Геном представлен однонитевой сегментированной (фрагментированной) негативной РНК. Вирион имеет суперкапсид, содержащий выступающие над мембраной в виде выступов (шипов) два гликопротеида - гемагглютинин (HA) и нейраминидазу (NA). Классификация вирусов гриппа основана на отличиях нуклеопротеиновых антигенов ( деление на вирусы А, В и С) и поверхностных белков HA и NA. У вирусов гриппа А выделяют 17 антигенно различных типов гемагглютинина и 10 типов нейраминидаз.
Основные функции гемагглютинина :
- распознает клеточный рецептор - мукопептид;
- отвечает за проникновение вириона в клетку, обеспечивая слияние мембран вириона и клетки;
- его антигены обладают наибольшими протективными свойствами. Изменения антигенных свойств (антигенные дрейф и шифт) способствуют развитию эпидемий, вызванных новыми Аг вариантами вируса (против которых не сформировался в достаточной мере коллективный иммунитет).
Нейраминидаза отвечает за диссеминацию вирионов, совместно с гемагглютинином определяет эпидемические свойства вируса.
Нуклеокапсид состоит из 8 сегментов вРНК и капсидных белков, образующих спиралевидный тяж.
Жизненный цикл вируса.
Репликация ортомиксовирусов первично реализуется в цитоплазме инфицированной клетки, синтез вирусной РНК осуществляется в ядре. В ядре на вРНК синтезируется три типа вирусспецифической РНК : позитивные матричные мРНК (матрица для синтеза вирусных белков), полноразмерная комплементарная кРНК (матрица для синтеза новых негативных вирионных РНК) и негативные вирионные вРНК (геном для вновь синтезируемых вирионов).
Вирусные белки синтезируются на полирибосомах. Далее вирусные белки в ядре связываются с вРНК , образуя нуклеокапсид. Заключительный этап морфогенеза контролируется М - белком. Нуклеокапсид, проходя через мембрану клетки, покрывается вначале М - белком, затем клеточным липидным слоем и суперкапсидными гликопротеинами HA и NA. Цикл репродукции составляет 6-8 часов и завершается отпочковыванием вновь синтезированных вирионов.
Антигенная изменчивость.
Современное разделение ортомиксовирусов на рода (или типы А,В и С) связано с антигенными свойствами главных белков нуклеокапсида (нуклеокапсидный белок - фосфопротеин NP) и матрикса вирусной оболочки (белок М). Кроме отличий по NP и M белкам, ортомиксовирусы отличаются высочайшей антигенной изменчивостью , обусловленной вариабельностью поверхностных белков HA и NA. Выделяют два основных типа изменений - антигенный дрейф и антигенный шифт.
Антигенный дрейф обусловлен точечными мутациями, изменяющими структуру этих белков. Основным регулятором эпидемического процесса при гриппе является популяционный (коллективный) иммунитет. В результате его формирования происходит отбор штаммов с измененной антигенной структурой (прежде всего гемагглютинина), против которых антитела менее эффективны. Антигенный дрейф поддерживает непрерывность эпидемического процесса.
Однако у вирусов гриппа А обнаружена и другая форма антигенной изменчивости - антигенный шифт (сдвиг), связанный со сменой одного типа гемагглютинина (или нейраминидазы) на другой, т.е. на появлении нового антигенного варианта вируса. Это наблюдается редко и связано с развитием пандемий. За всю известную историю гриппа выделено только несколько антигенных фенотипов, вызывающих эпидемии гриппа у людей : HoN1, H1N1, H2N2, H3N2, т.е. только три типа гемагглютинина (HA1-3) и два - нейраминидазы (NA 1 и 2). Вирусы гриппа типа В и С вызывают заболевания только у человека, вирусы гриппа А - у человека, млекопитающих и птиц. Наибольшую эпидемическую роль имеют наиболее изменчивые вирусы гриппа А. У вирусов гриппа С отсутствует нейраминидаза, эти вирусы обычно вызывают более легкую клиническую картину.
Существует мнение, что антигенный шифт - результат генетического обмена (рекомбинации) между вирусами гриппа человека и животных. До сих пор окончательно не установлено, где в межэпидемический период - вне человеческой популяции (у птиц или млекопитающих) или в человеческой популяции (благодаря длительной персистенции, локальной циркуляции) сохраняются вирусы, на время исчерпавшие свои эпидемические возможности.
Птиц считают первичными и основными хозяевами вирусов гриппа А, у которых в отличии от человека распространены вирусы со всеми 17 типами HA и 10 типами NA. Дикие утки - естественные хозяева вирусов гриппа А, у которых возбудитель находится в желудочно - кишечном тракте и не приносит хозяевам заметного ущерба. Вирусы проявляют свои патогенные свойства при переходе на других птиц и на млекопитающих. Среди млекопитающих наибольшее значение придают свиньям, которых считают промежуточным хозяином и сравнивают со “смешивающим сосудом”.
Современные вирусы гриппа человека слабо переходят на животных. Все пандемии гриппа А с 1930г. начинались в Китае, основными воротами распространения является Сибирь (массовые миграции птиц).
Н1N1- 1930г. Выявлен у человека, свиньи, китов (1972г.), домашних и диких птиц.
С ним связана знаменитая пандемия “испанки” (испанского гриппа). Этот тип вновь получил распространение с 1977г.
H2N2 выявляется с 1957г. у человека и птиц. Эпидемии, связанные с этими вирусами, приходили периодически. Сейчас оба типа выявляют параллельно.
H3N2 выявлен в 1963г. (Гонконг).
Вирус А/ Сингапур/1 /57 (H2N2) имеет три гена от вирусов гриппа птиц Евразии, вирус А/ Гонконг /1 /68 (H3N2) содержит 6 генов от вируса “Сингапур” и два - от птиц. Эти данные подтверждают, что новые эпидемические типы вирусов гриппа А человечество получает от птиц - первичного хозяина. Ближайший прогноз - возможность появления новых эпидемических вариантов вируса гриппа А, имеющих гемагглютинин HA5 или 7 (достаточно замены одной - двух аминокислот в их структуре).
Особенности патогенеза.
Возбудитель реплицируется в эпителии верхних дыхательных путей, вызывает гибель клеток. Вирусы и продукты распада клеток попадают в кровь, вызывают интоксикацию и повышение температуры тела. Вирусемия сопровождается множественными поражениями эндотелия капилляров и кровоизлияниями (от точечных до обширных геморрагий) в бронхах, трахее, легких, миокарде, различных паренхиматозных органах. Вирус обладает выраженным действием на иммунную систему - вызывает транзиторный иммунодефицит с элементами аутоиммунопатологии. Частое следствие этого - присоединение вторичных вирусных и бактериальных осложнений.
Постинфекционный иммунитет носит типоспецифический характер. Главную роль имеют вируснейтрализующие (против гемагглютинина и нейраминидазы) антитела - сывороточные IgG и секреторные IgAs, клеточные иммунные реакции.
Лабораторная диагностика.
Материал для выявления возбудителя - отделяемое носоглотки.
Вирусологические методы - заражение культур клеток или куриных эмбрионов. Типовую принадлежность определяют с помощью РСК, тип гемагглютинина - в РПГА, тип нейраминидазы - в реакции ингибирования активности этого фермента.
Экспресс - индикация - выявление антигена вируса в цитоплазме эпителиальных клеток носа и носоглотки при исследовании мазков - отпечатков методом флюоресцирующих антител (МФА), а также в ИФА и РНК - зондами.
Серологическая диагностика - выявление антител и нарастания титров антител в динамике (исследование парных сывороток) - в РТГА, РСК, ИФА, в т.ч. ИФА - выявление специфических IgAs- антител в слюне.
Лечение и специфическая профилактика.
В лечебных целях рекомендуют производные амантадина (рементадин), дибазол и различные индукторы интерферона, интерферон, противогриппозный иммуноглобулин.
Существует целый ряд вакцин против гриппа. Их основные типы :
- живая ослабленная (аттенуированная) вакцина, более иммуногенная и реактогенная;
- убитая (инактивированная) цельновирионная;
- субъединичные вакцины (содержат HA и NA);
- расщепленные и дезинтегрированные (различными детергентами, например).
Основная проблема всех вакцин - подобрать оптимальный набор антигенов с учетом конкретного эпидемического штамма.
Парамиксовирусы.
В это семейство также включены суперкапсидные “одетые” вирусы. Форма вириона сферическая, геном представлен однонитевой нефрагментированной негативной РНК, связанной с главным нуклеокапсидным белком NP. Оболочка содержит два гликопротеида - HN (обладает гемагглютинирующей и нейраминидазной активностью) и F (ответственен за слияние клеток - образование синцития и слияние мембран вируса и клеток, обладает гемолитической и цитотоксической активностью), а такде М- белок, формирующий внутренний слой вирусной оболочки.
Особенностью размножения парамиксовирусов в отличие от вирусов гриппа является отсутствие необходимости в “затравочной” мРНК и соответственно - в проникновении в ярдо клеток. Репликация полностью реализуется в цитоплазме клеток хозяина. Парамиксовирусы чаще вызывают инфекции у детей.
Вирусы парагриппа человека.
Вирусы парагриппа человека - одни из наиболее распространенных возбудителей ОРЗ. У взрослых парагрипп протекает клинически более легко, чем грипп. У детей отмечается более тяжелое течение заболевания. Часто вызывает ларингиты, ложный круп.
Антигенная структура.
На основе различий антигенной структуры HN, F и NP- белков вирусов парагриппа человека выделяют четыре основных серотипа. Типы 1, 2 и 3 антигенно родственны и перекрестно реагируют с антителами к вирусу паротита. Серотип 4 не имеет выраженного антигенного родства с остальными и имеет два подтипа. Серотип 3 чаще вызывает заболевания у детей до 1,5 лет (бронхиолиты, пневмонии), серотипы 1 и 2 часто вызывают ложный круп.
Патогенез - аналогичный большинству ОРВИ и гриппу.
Лабораторная диагностика в целом аналогична гриппу. Специфическая профилактика не разработана.
Вирус эпидемического паротита (свинки).
Представитель рода парамиксовирусов. Вызывает вирусную инфекцию, характеризующуюся преимущественным поражением околоушных слюнных желез.
Антигенная структура.
Известен один серовар. Антигенная структура аналогична другим парамиксовирусам.
Патогенез.
Первоначально вирус попадает в эпителий носоглотки, далее в кровоток. В период вирусемии он проникает в околоушные железы, яички, яичники, поджелудочную и щитовидную железы, головной мозг. Наиболее специфическое поражение - эпидидимит и орхит у мальчиков, возможно развитие серозных менингитов, панкреатитов и других осложнений.
Лабораторная диагностика.
В типичных случаях лабораторная диагностика может не проводиться. В случае необходимости используют вирусологические методы (выделение вируса на культурах фибробластов или куриных эмбрионах) - из спинномозговой жидкости, слюны, пунктатов желез), серологические методы (чаще РТГА или РСК, ИФА).
Специфическая профилактика.
Могут применяться живые аттенуированные вакцины, специфический донорский иммуноглобулин, проводится серопрофилактика.
Респираторно - синцитиальный вирус.
Возбудитель относится к роду Pneumovirus семейства парамиксовирусов, является одним из наиболее частых возбудителей острых респираторных заболеваний у детей первых лет жизни.
Вирион сферической формы. В отличии от других парамиксовирусов RS - вирус не обладает гемагглютинином, нейраминидазой, гемолитической активностью. В состав суперкапсида входят гликопротеиды - белки G и F, образующие поверхностные шипы. Белок G обеспечивает фиксацию вируса на чувствительных клетках, белок F - слияние двух типов : а) мембран вируса с мембранами клеток и их лизосом,
б) слияние инфицированных и неинфицированных клеток с образованием синцития - симпласта клеток, связанных цитоплазматическими отростками (“сетчатая ткань”). Вирус вызывает характерный цитопатический эффект и образование бляшек в культуре клеток.
Основные методы лабораторной диагностики.
1. Вирусологический - на культуре клеток определяют цитопатический эффект, идентификацию вируса проводят в МФА, РСК реакции нейтрализации.
2. Экспресс - диагностика в МФА, чаще исследуют клеточный материал из носоглотки.
3. Серологические методы - РСК, РН, ИФА.
Специфическая профилактика не разработана.
Вирус кори.
Вирус кори - представитель рода Morbillivirus семейства парамиксовирусов. По морфологии существенно не отличается от других представителей семейства. У него отсутствует нейраминидаза. Обладает гемагглютинирующей, гемолитической и симпластической активностью. Вирус имеет гемагглютинин, гемолизин (F), нуклеопротеид (NP) и матричный белок, отличающиеся антигенной специфичностью и иммуногенностью. Вирус кори имеет сероварианты, имеет общие антигенные детерминанты с другими морбилливирусами (вирусом чумы собак и вирусом чумы крупного рогатого скота).
Патогенез поражений.
Вирус первоначально размножается в эпителии верхних отделов дыхательных путей и регионарных лимфоузлах, затем проникает в кровь, гематогенно разносится по организму, фиксируется в ретикуло - эндотелиальной системе. Он вызывает поражения клеток эндотелия сосудов, сыпь, отек и некротические изменения тканей. Частые осложнения - пневмония, возможен отек гортани, круп, редко - энцефалит.
Лабораторная диагностика.
1.Метод экспресс - диагностики - обнаружение вирусных антигенов методом флюоресцирующих антител в пораженных клетках.
2. Вирусологическая диагностика - исследуют кровь до появления сыпи, слизь из носоглотки заражением культур клеток. Определяют цитопатический эффект, идентифицируют вирус в РТГА, РН и МФА.
3. Серологические методы - РСК, РТГА, ИФА.
Специфическая профилактика.
Применяют живые аттенуированные вакцины. У контактных можно проводить серопрофилактику противокоревым иммуноглобулином или иммуноглобулином донорским нормальным.
С проблемой морбилливирусов связан подострый склерозирующий панэнцефалит - очень тяжелое прогрессирующее заболевание нервной системы, протекающее по типу медленной инфекции. В основе поражений - персистенция вируса (близкого, но не идентичного вирусу кори) в клетках нейроглии с накоплением дефектных вирионов (без суперкапсида и белка М), с нарушением иммунного реагирования (высокие титры антител к вирусу кори и снижение клеточного иммунитета).
Лекция № 14. Энтеровирусы, вирусы гепатитов А и Е. Ротавирусы.
Большая часть острых кишечных инфекций (ОКИ) вызывается вирусами. Вирусов - возбудителей ОКИ известно более 120, среди них - энтеровирусы, ротавирусы, коронавирусы, калицивирусы, астровирусы и другие.
Семейство Picornaviridae объединяет четыре рода - Enterovirus, Cardiovirus, Rhinovirus, Aphtovirus. Это мелкие “голые” вирусы с икосаэдральной (кубической) симметрией. Геном образован несегментированной позитивной РНК. Репликация РНК и сборка вируса осуществляется в цитоплазме, выход вируса сопровождается лизисом клетки. Репликация вРНК осуществляется по схеме: вРНК-> кРНК-> вРНК. Название семейства происходит от pico (лат. - маленький) и RNA (РНК), т.е. маленькие РНК- вирусы. Из рода афтовирусов наибольшее значение имеет вирус ящура. Представители рода риновирусов (более 100 серотипов) - возбудители острых респираторных инфекций.
Род энтеровирусов.
Род объединяет несколько групп вирусов: полиовирусы (1 - 3 типов), вирусы Коксаки А (24 серовара), вирусы Коксаки В (6 сероваров) и ECHO (34 серовара), а также неклассифицированные вирусы (вирусы 68 - 72). Энтеровирус 72 - возбудитель гепатита А.
Все энтеровирусы кислотоустойчивы (могут выживать в кислой среде желудка), отсутствие оболочки обусловливает их устойчивость к действию жировых растворителей и желчных кислот (особенно устойчивы полиовирусы). Патогенные виды поражают желудочно - кишечный тракт. Для них характерны : фекально - оральный механизм заражения, летне - осенняя сезонность, выделение вирусов из кишечника, носоглотки, ликвора и крови, обнаружение в сточных водах, широкое носительство, преимущественное поражение детского населения.
Полиовирусы.
Полиовирусы вызывают полиомиелит - острую инфекцию с поражением нейронов продолговатого мозга и передних рогов спинного мозга. Важнейшее биологическое свойство полиовирусов - тропизм к двигательным клеткам серого вещества спинного мозга (polios - серый, myelitis - воспаление спинного мозга).
Капсид вириона образован четырьмя белками, образующими внешнюю (VP1, VP2, VP3) и внутреннюю (VP4) поверхности капсида. Белки оболочки имеют значение в распознавании и прикреплению к клеточным рецепторам, высвобождении вирионной РНК внутри клетки, развитии параличей.
По антигенным свойствам полиовирусы подразделяют на три типа, наибольшей вирулентностью и эпидемической активностью обладают полиовирусы 1 типа.
Патогенез поражений. Входными воротами для полиовирусов являются слизистые глотки, желудка, тонкого кишечника. После размножения в эпителиальных клетках вирус проникает в регионарные лимфатические узлы, затем - в кровь (первичная вирусемия). Эти первые стадии характеризуют “малую болезнь”, которая может протекать практически бессимптомно (легкое недомогание, кратковременное повышение температуры) и заканчиваться формированием постинфекционного иммунитета и выздоровлением, что и происходит в большинстве случаев.
Если полиовирус преодолевает гематоэнцефалитический барьер и поражает нейроны передних рогов спинного мозга, продолговатого мозга и варолиевого моста, несущие рецепторы к полиовирусам, развивается “большая болезнь” - паралитические формы (спинальный полиомиелит обычно с ассиметричными поражениями нижних конечностей, бульбарный полиомиелит в ряде случаев с поражениями центров, контролирующих дыхательные мышцы, а также сочетанные спинально - бульбарные поражения.
Лабораторная диагностика имеет особое значение, особенно при стертых формах, поскольку многие энтеро - и герпетовирусы способны вызывать схожие поражения. Поэтому исследования необходимо проводить одновременно на все эти группы вирусов.
1.Вирусологическая диагностика включает выделение вируса на различных культурах клеток или (в некоторых случаях - Коксаки А) на новорожденных белых мышах, с последующей идентификацией по цитопатическому эффекту, в РН, РТГА, РСК с эталонными сыворотками.
2. Серологическая диагностика осуществляется в различных реакциях (в настоящее время - ИФА), необходимо исследование в парных сыворотках, выявление специфических IgM - антител.
Иммунитет и специфическая профилактика.
Иммунитет к полиовирусам прочный, обусловленный вируснейтрализующими антителами и клетками иммунной памяти. Для специфической профилактики используют убитые (вакцина Солка) и живые (вакцина Сэбина) ослабленные вакцины (содержат аттенуированные штаммы полиовирусов 1, 2 и 3 типов). Существуют программы массовой иммунизации против полиомиелита и программы полной ликвидации этой инфекции.
На фоне резкого снижения и ликвидации полиомиелита наблюдаются полиомиелитоподобные заболевания, вызываемые преимущественно вирусами Коксаки и ECHO.
Вирусы Коксаки.
Они образуют группу, близкую к полиовирусам. Вирусы Коксаки групп А и В в отличии от полиовирусов патогенны для новорожденных мышей, отличаясь друг от друга характером вызываемых у них поражений (группа А - преимущественные поражения скелетной мускулатуры, группы В - преимущественные поражения центральной нервной системы).
Антигенная структура. Вирусы Коксаки не дают перекрестных серологических реакций с полиовирусами, группа А имеет 24 серовара, группа В - 6 сероваров. Серовары не содержат группоспецифического антигена, однако обладают некоторой перекрестной реактивностью. Наличие у возбудителей типоспецифических антигенов обусловливает синтез типоспецифических антител. Серовары Коксаки В и некоторые серовары Коксаки А обладают в отличии от полиовирусов гемагглютинирующими свойствами.
Клинические проявления.
Среди всех энтеровирусов вирусы Коксаки (особенно группы В) обладают наибольшей кардиотропностью, вызывая миокардиты. Наиболее часто они поражают детей, вызывают в большинстве случаев легкие формы преимущественно с “простудной” симптоматикой. Наряду с полиомиелитоподобными заболеваниями (вялыми параличами) и миокардитами, вирусы этой группы способны вызывать ОРЗ, гастроэнтериты, герпангины и пузырчатку полости рта и конечностей.
Лабораторная диагностика - как у всех энтеровирусов. Принадлежность к сероварам определяют в РСК или РН с типоспецифическими сыворотками.
Эффективных методов специфической профилактики и противовирусной терапии не разработано.
ECHO - вирусы получили свое название от слов Enteric (кишечные) Cytopathogenic (цитопатогенные) Human (человеческие) Orphan (сиротские) вирусы. Они были выделены из кишечника человека, по ряду признаков оказались схожими с полиовирусами и вирусами Коксаки, однако первоначально не были связаны с какими - либо заболеваниями (т.е. оказались “сиротскими”).
Классификация и антигенная структура.
В настоящее время кишечная группа ECHO- вирусов насчитывает 34 серовара. Разделение основано на типоспецифичности антигенов вирусного капсида. Некоторые антигены обладают перекрестной реактивностью, 12 серотипов способно к гемагглютинации.
Патогенез заболеваний, вызываемых ECHO- вирусами сходет с патогенезом полиомиелита. Заражение происходит преимущественно фекально - оральным путем. Размножение вирусов происходит в эпителиальных клетках слизистых, а также в лимфоидной ткани. Способны вызывать “простудные заболевания” (по типу ОРВИ), некоторые серотипы (11, 18 и особенно 19) - кишечные диспепсии, более редко - менингиты, восходящие параличи и энцефалиты, отдельные серотипы - гепатиты, конъюнктивиты, увеиты (серотипы 11 и 19).
Группа неклассифицированных энтеровирусов. Из этой группы наибольшее значение имеет энтеровирус 72 - вирус гепатита А (возбудитель болезни Боткина) - HAV (hepatitis A virus).
Вирусные гепатиты представляют большую разнородную по этиологии, но схожую по клиническим проявлениям группу тяжелых (по последствиям) заболеваний, широко распространенных в мире. Вирус гепатита А - энтеровирус 72, В - гепадновирус, С и G - тогавирусы рода Flavivirus, D - неклассифицированный вирус, Е - калицивирус. Из них вирусы гепатитов А и Е характеризуются преимущественно фекально - оральным механизмом передачи, В, С и G - парентеральным (гемоконтактным), D (дельта)- является дефектным вирусом - сателлитом вируса гепатита В, передаваемым парентерально и вертикально (от матери плоду).
Вирус гепатита А имеет “голый” капсид с кубическим типом симметрии - икосаэдр. Геном образует однонитевая молекула позитивной РНК. Белковая оболочка (капсид) содержит 4 структурных белка - VP1, VP2, VP3, VP4. HAV является одним из наиболее устойчивых во внешней среде вирусов.
Антигенная структура.
Вирус имеет один антигенный тип и содержит главный антиген (НА Аг), развитие иммунного ответа к которому обеспечивает прочный пожизненный иммунитет.
Патогенез поражений.
Вирус проникает в организм в результате реализации фекально - орального механизма заражения, реплицируется в эпителии слизистой тонкой кишки и регионарных лимфатических узлах, затем проникает в кровь (наибольшие титры вируса в крови - в конце инкубационного и в преджелтушный период), выделяется с фекалиями. Затем возбудитель проникает в печень и вызывает острый диффузный гепатит, связанный с поражением гепатоцитов (основной мишени для размножения и цитопатогенного действия вируса) и ретикуло - эндотелиальных элементов печени. Это сопровождается снижением барьерной и дезинтоксикационной функций печени, нарушениями белкового, углеводного и пигментного обмена, возрастанием уровня в сыворотке крови альдолазы и печеночных (разрушение гепатоцитов) аминотрансфераз (аланин - и аспартат - аминотрансфераз), билирубина.
Клинические особенности.
Наиболее типична острая желтушная циклическая форма, однако преобладают легкие безжелтушные и бессимптомные формы. Для этой инфекции характерно относительно легкое течение, практическое отсутствие вирусоносительства и хронических форм болезни.
Лабораторная диагностика
1. Определение желчных пигментов и аминотрансфераз в сыворотке крови.
2. ИФА для выявления антигенов вируса и IgM- антител к нему. Антигены HAV в фекалиях можно выявить только в конце инкубации до появления клинических проявлений. Наиболее надежный метод диагностики - обнаружение ранних антиHAV - IgM антител. Они выявляются практически у всех больных независимо от формы заболевания и свидетельствуют о наличии текущей или недавней инфекции.
Специфическая профилактика.
Используют инактивированные вакцины против вируса гепатита А отечественного “Геп - А инвак”) и зарубежного (“Хаврикс 1400” фирмы “Смит Кляйн Бичем”) производства. Трехкратная (при рождении , в 1 и 6 месяцев) вакцинация формирует защитный иммунитет у 99% детей. Вирус гепатита Е
Общая характеристика.
Гепатит Е - инфекция, имеющая эпидемиологическое сходство с гепатитом А, известная ранее как “гепатит ни -А, ни -В с фекально - оральным механизмом передачи”. Возбудитель - вирус гепатита Е (HEV) относится к семейству калицивирусов, однако в последние годы это ставится под сомнение. Он имеет сферическую форму, диаметр вириона около 30 нм, суперкапсида не имеет. Геном представлен однонитевой нефрагменторованной позитивной РНК. Антигенного родства с HAV не имеет, отличается меньшей вирулентностью для человека. Представители семейства калицивирусов по ряду признаков близки пикорнавирусам. Свое название они получили от лат. calix - чаша за счет наличия чашеобразных углублений на поверхности капсида. Среди представителей семейства - возбудители гастроэнтеритов животных (например, вирус Норфолк), вирус гепатита Е.
Эпидемиология.
Вирус имеет фекально - оральный механизм распространения, реализуемый преимущественно при употреблении инфицированной воды. Широко распространен в странах с жарким климатом и плохим водоснабжением, при употреблении некачественной воды (из арыков, оросительных каналов, загрязнении грунтовыми водами и т.д.) происходит заражение. Вспышки наблюдают в Юго - Восточной Азии, Африке, Южной Америке. Инфекция распространена в Среднеазиатском регионе бывшего СССР. Особенность - высокая летальность HEV- инфекции у беременных во второй половине беременности.
Лабораторная диагностика.
Разработаны тест - системы ИФА и иммуноблота для выявления антител к HEV классов IgM и IgG, пригодные для диагностики в острой стадии болезни и в период реконвалесценции.
Профилактика ВГЕ - неспецифическая. Иммунитет - прочный, обусловлен вируснейтрализующими антителами и клетками памяти.
Ротавирусы.
В семейство реовирусов входят рода реовирусов, ротавирусов и орбивирусов. Название реовирусов происходит от respiratory enteric orphan - “reo”, т.е. респираторные кишечные вирусы. Эти вирусы имеют двунитевую фрагментированную РНК, окруженную капсидом с двуслойной оболочкой (наружный и внутренний капсид). Реовирусная инфекция характеризуется поражением респираторного и кишечного трактов. Основной путь передачи - воздушно - капельный, однако важное значение имеет и алиментарный, поскольку эти вирусы широко распространены не только у человека, но и у многих видов животных, люди могут получать эти вирусы алиментарным путем - с продуктами питания.
Ротавирусы свое название получили в связи со своеобразием морфологии вируса (лат. rota - колесо). Впервые ротавирусы выявлены у детей с острым гастроэнтеритом их эпителиальных клеток слизистой оболочки двенадцатиперсной кишки. Вирионы при электронно - микроскопическом исследовании имеют вид колес диаметром 70 нм с круговым ободком по периферии и отходящими внутрь “спицами”. Имеют наружный и внутренний капсид, внутри которого содержится двунитевая фрагментированная РНК. К ротавирусам человека серологически близки ротавирус телят (Небраски) и обезьян, которые легче культивировать в клеточных культурах и которые используют в качестве антигенных препаратов для серодиагностики ротавирусной инфекции у людей. Ротавирус человека - фекально - оральный вирус. Он вызывает гастроэнтериты у новорожденных (внутрибольничные вспышки), дошкольников и младших школьников. В мире ежегодно регистрируют до нескольких миллионов летальных исходов ротавирусного гастроэнтерита у детей. Наиболее опасны больные в первые 3-5 дней заболевания в связи с интенсивным выделением вируса с фекалиями. Лабораторная диагностика основана на выявлении антигена вируса в фекалиях (ИФА, реакция коагглютинации, иммунофлюоресцентный анализ, РНК- зонды, иммунная электронная микроскопия), серологических методах (РСК, РТГА, ИФА), изоляции на культурах клеток.
Орбивирусы - вирусы клещей и насекомых (например, вирус Кемерово).
Лекция №15. Вирусы гепатитов В, С, D и G.
Вирусные гепатиты представляют большую разнородную по этиологии, но схожую по клиническим проявлениям группу тяжелых по последствиям заболеваний, широко распространенных в мире. Многие вирусы могут вызывать среди прочих действий и поражения печени (например, некоторые серотипы ECHO- вирусов), однако имеется большая группа обладающих преимущественно гепатотропным действием вирусов. Они по эколого - эпидемиологическим особенностям могут быть разделены на две группы - с преимущественно фекально - оральным механизмом передачи (вирусы гепатитов А и Е) и с парентеральным (гемоконтактным) путем передачи (В, С, G, D). Вирус гепатита D (дельта) является дефектным вирусом - сателлитом вируса гепатита В, передаваемым парентерально и вертикально (от матери плоду). Вирус гепатита А - энтеровирус 72, В- гепадновирус, С и G- флавивирусы, D- неклассифицированный вирус, Е- калицивирус.
Вирус гепатита В.
Вирус гепатита В (HBV) вызывает сывороточный гепатит, относится к семейству гепадновирусов - оболочечных ДНК - вирусов, вызывающих гепатиты у различных видов животных (сурков, уток и др.).
Гепатит В - серьезная проблема здравоохранения во всем мире. Этому способствует рост заболеваемости, частое формирование неблагоприятных исходов (хронический гепатит, цирроз печени, гепатокарцинома, довольно высокая летальность).
Гепадновирусы поражают преимущественно клетки печени. Геном HBV представлен двуцепочечной кольцевой молекулой ДНК, наружная цепь длиннее внутренней.
Цикл репродукции HBV очень сложен и проходит через промежуточное звено - РНК ( ДНК→ РНК→ ДНК), т.е. с механизмом обратной транскрипции. При транскрипции вирусного генома в ядре гепатоцита клеточная ДНК - зависимая РНК - полимераза синтезирует два типа мРНК - большего размера (прегеном) и меньшего размера (для синтеза вирусных белков). Прегеном и вирусная ДНК - полимераза упаковываются в капсид и переносятся в цитоплазму. Под действием вирус - индуцированной обратной транскриптазы на матрице прегенома (РНК) синтезируется новая минус - нить ДНК. Вирионная ДНК - полимераза на минус - цепи синтезирует плюс -цепь. Если вирусная двухцепочечная ДНК не вступает в дальнейшую репликацию, сформировавшийся нуклеокапсид, проходя через мембрану клетки, покрывается суперкапсидом и отпочковывается от клетки.
Строение и антигенная структура.
Вирусные частицы размером 42 - 45 нм (частицы Дейна) имеют достаточно сложное строение и включают ДНК, ассоциированную с ней ДНК - полимеразу и четыре антигена - поверхностный (HBs Ag - “австралийский”), сердцевинный или коровский (HBc Ag или cor Ag), антиген инфекционности (HBe Ag, выявляемый в крови при активной репликации HBV) и наименее изученный HBx Ag.
Циркулирующие штаммы HBV отличаются по антигенной структуре HBs антигена. В его составе имеется общий антиген , обусловливающий перекрестный (между субтипами) иммунитет и четыре типоспецифических антигенных детерминанты и соответственно четыре субтипа HBs Ag ( и HBV).
С учетом сложной антигенной структуры вируса гепатита В, в диагностике данной инфекции используют целый ряд маркеров инфицирования, в т.ч. антигены (HBs Ag, HBc Ag, HВe Ag) и соответствующие им антитела (анти - HBs, анти - HBc и анти - HВe).
Важное значение для определения прогноза и тактики лечения больных гепатитом В имеет выделение двух качественно различных биологических фаз развития HBV - репликативной и интегративной. При репликативной фазе (т.е. массовом размножении вируса) вирусная ДНК - полимераза реплицирует ДНК HBV и все вирусные субкомпоненты и белки копируются в большом количестве. При интегративной фазе развития (т.е. когда вирусные частицы не подвергаются дальнейшей репликации) геном HBV встраивается в геном гепатоцита. В процессе интеграции основную роль играет фрагмент, несущий ген, кодирующий HBs антиген, поэтому при этой фазе идет преимущественное образование HBs Ag. Следовательно, биологические стадии HBV отличаются спектром выявления маркеров инфицирования. Для стадии репликации характерно выявление ДНК HBV, HBe Ag и (или) анти - HBc - IgM, возможно - HBs Ag. В стадии интеграции преобладает HBs Ag, анти - HBe, анти - HBc - IgG.
Эпидемиологические особенности.
Вирус гепатита В распространяется эволюционно сложившимися естественными и искусственными путями распространения. Для эффективного заражения оказывается достаточным введение 0,0000007 мл инфицированной крови (искусственные парентеральные пути заражения - через медицинские манипуляции) . Среди естественных путей - вертикальный ( от матери - потомству), половой и контактный (семейный) - “гемоконтактный” (Л.М. Шляхтенко и др., 1990,1998). Передача возбудителя обеспечивается устойчивостью во внешней среде, вирус передается при контакте с кровью и другими биологическими жидкостями организма (слюна, сперма, содержимое носоглотки, вагины и др.). Факторами передачи могут быть различные предметы личной гигиены (зубные щетки, бритвенные и маникюрные приборы, мочалки, расчески и др.). В последние годы увеличилось значение наркозависимого и полового пути передачи.
Клинико - патогенетические особенности.
Орган - мишень для вируса гепатита В - печень. Поражение гепатоцитов напрямую не связано с непосредственным действием вируса (цитопатического эффекта нет), а с иммунными (аутоагрессивными) реакциями хозяина, связанными с модификацией клеточных мембран вирусными белками. Аутоагрессия реализуется Т - цитотоксическими лимфоцитами и другими киллерными клетками, выработкой аутоантител против тканей печени. Поражения печени могут быть в виде острой и хронической форм различной тяжести.
Постинфекционный иммунитет длительный, направлен против основного протективного HBs антигена, обусловлен вируснейтрализующими анти - HBs антителами. Методы диагностики.
В основе лабораторной диагностики - ИФА и ПЦР.
HBs антиген - основной и первый маркер инфицирования HBV. Его элиминация и появление анти - HBs - антител является непременным условием выздоровления. Анти HBs - антитела - показатель перенесенной инфекции.
HBc антиген - сердцевинный антиген, белок нуклеокапсида, выявляемый только в ядрах гепатоцитов, но отсутствующий в чистом виде в крови. Большое диагностическое значение имеет определение в крови анти- HBc - IgM. Эти антитела при остром гепатите выявляются раньше, чем антитела к другим вирусным антигенам. АнтиHBc -IgM выявляют у 100% больных острым гепатитом В, как HBs - позитивным, так и HBs - негативным. Анти HBc - антитела могут быть единственным маркером вируса гепатита В в фазу “окна”, когда в крови не удается выявить ни HBs антиген, ни антител к нему.
Выявление анти- HBc -IgM сочетается с выявлением ДНК вируса и ДНК - полимеразной активности (т.е. показателями продолжающейся репликации вируса) и активностью патологического процесса в печени. Анти HBc - IgG - маркер перенесенного инфицирования HBV.
HBe Ag - антиген инфекционности, циркулирует только при наличии HBs антигена. Его наличие в сыворотке крови коррелирует с выявлением ДНК вируса, полимеразной активностью и с продукцией полных вирусных частиц, т.е. с активной репликацией вируса. Длительность циркуляции HBe антигена - важный прогностический признак. Его выявление через два месяца с начала заболевания - признак вероятного развития хронического гепатита. В большинстве случаев происходит смена HBe Ag на антиHBe - антитела, что является маркером завершенной репликации вируса гепатита В.
Важную диагностическую информацию представляют методы выявления ДНК HBV. В ряде случаев при отсутствии в крови HBs антигена, а также серологических маркеров вирусной репликации (HBe Ag, антиHBcor - IgM), о продолжающейся репродукции вируса в печени можно судить по результатам молекулярной гибридизации нуклеиновых кислот (МГНК) и ПЦР. Используя технику ПЦР, можно определить и подтип HBs антигена.
Специфическая профилактика в настоящее время осуществляется с использованием рекомбинантных вакцин (“Энжерикс В”, “Рекомбивакс В” и др.), полученных методами генной инженерии на культурах дрожжей Saccharomyces cerevisae. Рекомбининтный клон дрожжей вырабатывает поверхностный антиген HBV. Эффективность - 95% , продолжительность - не менее 5 - 6 лет. Предусмотрена трехкратная иммунизация - сразу после рождения, через 1 - 2 месяца, до конца первого года жизни ребенка. Для экстренной профилактики контактным может применяться донорский иммуноглобулин, содержащий антитела к HBV.
Вирус гепатита С.
Гепатит С - инфекция, вызываемая вирусом гепатита С (HCV). Раньше эту инфекцию обозначали как “гепатит ни - А, ни - В, передающийся парентерально”. Вирус HCV является РНК - содержащим вирусом семейства флавивирусов.
Морфология.
HCV - оболочечный вирус со средним размером вириона 35 - 50 нм. Геном образует однонитевая позитивная РНК. Различные гены кодируют структурные (капсидный, мембранный и оболочечный) и неструктурные белки.
К настоящему времени идентифицировано от 6 до 11 генотипов и более 80 подтипов HCV (Львов Д.К., Дерябин П.Г.,1997). Степень гомологии генотипов - порядка 65%, подтипов - 77 - 79%. Различия в генотипе определяют тяжесть заболевания, ответную реакцию на лечение, взаимодействие вируса с хозяином.
Клинико - эпидемиологические особенности.
Эпидемиология вирусного гепатита С напоминает эпидемиологию гепатита В. Несмотря на относительно легкое течение, около половины случаев приводит к хроническим гепатитам, в ряде случаев заканчивающихся циррозом и карциномой печени. Вирус более редко (по сравнению с HBV) передается вертикально и половым путем, преобладает парантеральное заражение (особо - наркоманы). Показана общность групп риска инфицирования HBV и HCV. Выявлены географически преобладающие варианты (генотипы) HCV. В Европейской части России преобладает “западный” генотип, в Азиатской части - “восточный” (близкий к китайскому и японскому).
Лабораторная диагностика.
В настоящее время лабораторная диагностика вирусного гепатита С основана на определении сывороточных маркеров инфицирования - антител суммарных (анти - HCV) и анти - HCV IgM, а также детекции РНК HCV.
Во все тест - системы, начиная с тест - систем “второго поколения” включены аминокислотные последовательности структурного сердцевинного нуклеокапсидного белка (HCc Ag). Антитела к HCc Ag являются самими ранними маркерами инфицирования HCV. С целью повышения эффективности и специфичности ИФА применяют подтверждающие тесты - рекомбинантный иммуноблоттинг или вестерн - блоттинг, позволяющие определить антитела к различным структурным и неструктурным белкам HCV. Анти-HCV - IgM антитела свидетельствуют о репликативной активности вируса гепатита С. Снижение титров IgM - антител свидетельствует о благоприятном течении заболевания.
Определение РНК HCV методом ПЦР свидетельствует об виремии, которая может быть транзиторной (острый гепатит с последующим выздоровлением), персистирующей (хронические гепатиты) и прерывающейся (повторное выявление).
Специфическая профилактика. В настоящее время проводится работа по созданию вакцин.
Вирус гепатита D (HDV).
Гепатит Д - инфекция, вызываемая HDV, характеризующаяся тяжелым поражением печени. Эпидемиологически связана с гепатитом В. Возбудитель - дефектный РНК - содержащий вирус, выделяемый только от пациентов, инфицированных HBV. Дефектность вируса заключается в его полной зависимости от HBV, который является для него вирусом - помощником. Поэтому гепатит D протекает либо как коинфекция (одновременное заражение HBV и HDV), либо как суперинфекция (наслоение HDV на текущую HBV- инфекцию, чаще хроническую).
Вирус гепатита Д представляет собой сферическую частицу средним диаметром 36 нм. Суперкапсид состоит из HBs Ag и кодируется HBV. Внутренний HD Ag кодируется HDV. По структуре генома, представленной кольцевидной РНК, HDV напоминает вироиды, является неклассифицированным к настоящему времени вирусом. Вирус гепатита Д является вирусом - спутником (сателлитом) HBV, его распространение связано с распространением HBV.
Клинико - эпидемиологические особенности.
Наибольшее значение в распространении имеют искусственные пути распространения (медицинские манипуляции). Естественные пути - аналогичны HBV. Вирус гепатита Д не может вызывать развитие гепатитов без одновременной репликации HBV. Наиболее опасна суперинфекция, вызванная HBV и HDV, у 60 - 70% пациентов с хроническими гепатитами такого типа наблюдается цирроз печени. При гепатите Д возможно острое и хроническое течение инфекции.
Лабораторная диагностика.
Для постановки диагноза имеют значение следующие маркеры инфицирования : дельта - антиген, антитела к нему класса IgG (анти - HDV - IgG) и IgM (анти - HDV -IgM), РНК HDV. Дельта - антиген для тест - систем ИФА и РИА получают из печени инфицированных шимпанзе или генно - инженерными методами (экспрессия антигена рекомбинантными штаммами E.coli).
При коинфекции в желтушный период на протяжении двух недель выявляют HDV Ag, РНК вируса, антиHDV- IgM. Несколько позднее (через 1 - 2 месяца с момента появления желтухи) начинают выявлять антиHDV IgG. Одновременно выявляются маркеры активной репродукции HBV (HBs Ag, HBe Ag, антиHBc - IgM, ДНК HBV).
При суперинфекции в острый период HDV Ag может не выявляться или выявляться кратковременно. Наряду с HDV- IgM выявляются HDV- IgG, причем в более высоких титрах, чем при коинфекции. При суперинфекции HDV происходит снижение уровня HBs Ag, падают титры или полностью исчезают HBcor- IgM. Стойкое увеличение HDV антител является маркером хронического HDV гепатита.
С учетом зависимости вируса гепатита Д от вируса гепатита В в основе профилактики гепатита дельта - профилактика гепатита В. Специфическая профилактика - вакцинация против гепатита В - основное мероприятие профилактики гепатита Д.
Вирус гепатита G.
Гепатит G вызывается РНК - содержащим вирусом из группы флавивирусов. Вирус передается парентерально, способен вызывать острые и хронические гепатиты. Описано несколько генотипов HGV. У этого вируса обнаружено два структурных оболочечных белка - Е1 и Е2 и пять неструктурных белков. Маркеры гепатита G распространены среди населения даже чаще, чем - вируса гепатита С, особенно при трансплантации почки, у наркоманов, больных гемофилией, при гемодиализе и наиболее часто - у носителей вируса гепатита С.
Как метод лабораторной диагностики чаще используют ПЦР - определение РНК вируса (при остром гепатите G в первые шесть месяцев можно определить только вРНК), антитела к оболочечному белку Е2 выявляют не ранее 5 - 6 месяцев с момента инфицирования.
Острый гепатит G переходит в хронический реже, чем гепатит С. С хроническими гепатитами G связана персистенция вируса, возникновение циррозов печени. Гепатит G к настоящему времени изучен еще недостаточно.
Современные представления о вирусных гепатитах изменяются быстро, список вирусов расширяется. К новым гепатотропным вирусам, действие которых уточняется, следует отнести TTV (ДНК- вирусы нового семейства Circinoviridae) и SEN- вирусы.
TTV- вирус- спутник, сопутствующий известным вирусным парентеральным гепатитам. Это- вирус - оппортунист, активирующийся при иммунодефицитах (в т.ч. ВИЧ- инфекции). SEN- вирусы- это ДНК -вирусы, близкие к семейству TTV, связаны с переливаниями крови, реплицируются в гепатоцитах, чаще встречаются у больных с парентеральными вирусными гепатитами В и С, генетически высоко изменчивы.
Лекция №16. Ретровирусы. Вирус иммунодефицита человека.
Вирус иммунодефицита человека (ВИЧ или HIV) относится к семейству ретровирусов, подсемейству лентивирусов (медленных вирусов). Геном ретровирусов уникален - он представлен двумя идентичными молекулами позитивной РНК, т.е. это РНК - вирусы с диплоидным геномом. Свое название ретровирусы получили за отличительные особенности репродукции (РНК → ДНК → иРНК → геномная РНК). Особенности репродукции связаны с функциями фермента обратной транскриптазы (ревертазы или РНК - зависимой ДНК - полимеразы), обладающей тремя видами активности - обратной транскриптазы, РНК - азы и ДНК - полимеразы.
Семейство Retroviridae включает три подсемейства.
1. Lentivirinae - возбудители медленных вирусных инфекций, в т.ч. ВИЧ.
2. Oncovirinae - онкогенные вирусы, с которыми связано превращение клеток в опухолевые. Раньше не знали, как РНК - вирусы могут встраиваться в геном клетки и способствовать опухолевому росту (не были известна возможность обратной транскрипции у вирусов), что тормозило научную разработку вирусологии опухолевого роста.
3. Spumavirinae - “пенящие” вирусы, название которых связано с характерным
“вспененным” видом инфицированных ими клеточных культур как результатом интенсивного симпластообразования.
Первыми открытыми в конце 70- х годов ретровирусами были HTLV-1 и HTLV-2 (от “human T- lymphotropic virus) - возбудители Т - клеточных лейкозов и лимфом. Наиболее известны обладающие лимфотропным и цитопатическим действием вирусы ВИЧ-1 (HIV-1 в английском варианте) и ВИЧ-2, вирус иммунодефицита обезьян (ВИО или SIV), к которому по ряду свойств ближе (чем к ВИЧ-1) ВИЧ-2. К настоящему времени имеется информация уже о нескольких сотнях ретровирусов, интегрированных с геномом человека и практически очень мало об их роли в патологии. История изучения и происхождения ВИЧ.
Синдром приобретенного иммунодефицита (СПИД) впервые выделен как самостоятельное заболевание в США в 1981г. Возбудитель (ВИЧ-1) был описан почти одновременно в 1983г. французом Л.Монтанье и американцем Р.Галло. Эпидемиология нового вида патологии поражала своей необычностью. Почти 100% больных были мужчинами в возрасте 25-49 лет, 94% - гомо- или бисексуалами, отмечалась высокая летальность. У больных были выявлены дефекты клеточного иммунитета, развитие пневмоцистозной пневмонии, кандидоза и саркомы Капоши у них было расценено как оппортунистические заболевания. СПИД окрестили болезнью четырех “H”- по первым буквам английских вариантов слов гомосексуалы, гемофилия, гаитяне и героин. В 1986г. был идентифицирован еще один вирус - ВИЧ-2.
Окончательного ответа на вопрос о месте, времени и условиях возникновения ВИЧ нет. Ретроспективные исследования показали циркуляцию этого вируса по крайней мере с конца 50-х - начала 60-х годов. Прародительницей ВИЧ считают тропическую Африку, где широко распространен ВИО (близкородственный ВИЧ-2) среди обезьян. Первые известные серологические находки ВИЧ отмечены в Африке, здесь же - наибольшая интенсивность передачи ВИЧ наиболее естественным гетеросексуальным путем. Однако эпидемическое распространение ВИЧ - инфекция получила с конца 70-х - начала 80-х годов. В 1987г. выявлен первый случай в России, в настоящее время счет идет на десятки тысяч инфицированных в год. В мире регистрируют десятки миллионов инфицированных ВИЧ в год, ежегодно число вновь инфицированных увеличивается, т.е. налицо пандемия ВИЧ - инфекции.
Структура вириона ВИЧ.
ВИЧ имеет сферическую форму и размеры 100-120 нм в диаметре. Наружная оболочка образована двойным липидным слоем с гликопротеиновыми “шипами”, состоящими из трансмембранного белка gp41 (пронизывает липидный слой) и наружного белка gp120. Эти оболочечные белки кодируются геном env и участвуют в прикреплении вириона к мембранам клеток хозяина. С внутренней стороны липидной оболочки находится матричный каркас, образованный белком р17. Он окружает внутреннюю структуру вириона - нуклеокапсид или сердцевину (англ. - core). Собственная оболочка сердцевины образована “коровским” белком р24. Внутри нуклеокапсида находится геном вируса в виде двух цепочек, связанных белками р7 и р9, полимеразный комплекс ревертазы, протеаза, интеграза (эндонуклеаза), затравочная т - РНК. Наиболее распространен ВИЧ-1, который в зависимости от строения гена env имеет субтипы. Субтипы A-H составляют доминирующую группу M (major), наиболее распространены субтипы С и Е.
Жизненный цикл ВИЧ.
Инфекционный процесс при заражении ВИЧ носит последовательный фазовый характер и начинается с проникновения вируса через слизистую оболочку половых путей или с непосредственного поступления в кровоток. Проникнув в организм, вирус в первую очередь атакует клетки, имеющие специфичный для него рецептор CD4. Этот рецептор имеют в большом количестве Т - хелперы, в меньшем - макрофаги и моноциты, нейроны, клетки нейроглии и некоторые другие клетки (см. лекции по общей иммунологии). Вирус распознает CD4 - рецепторы с помощью своего белка gp120. Процесс инфицирования клетки вирусом осуществляется в два этапа : прикрепления и слияния. Прикрепленный через белок gp120 к рецептору CD4 клетки - мишени вирус белком оболочки gp41 сливается с мембраной клетки. Белок gp41 обеспечивает не только слияние вирусной и клеточной мембран, но и слияние мембран клеток с образованием синцития (многоядерных клеток), обреченного на гибель. Нуклеокапсид, освобожденный от суперкапсида при слиянии мембран, попадает в цитоплазму. На пути к ядру освобождается геномная РНК и ассоциированные с ней компоненты сердцевины. Обратная транскриптаза синтезирует на вирионной РНК минус - цепь ДНК, РНК-аза разрушает вирионную РНК, а вирусная ДНК - полимераза синтезирует на минус - цепи плюс - цепь ДНК.
Двунитевая ДНК транспортируется в ядро клетки, где приобретает кольцевую форму и интегрируется под действием эндонуклеазы (интегразы) с ядром клетки, превращаясь в ДНК - провирус.Последующие этапы жизненного цикла ВИЧ - латентная фаза, фаза активации транскрипции с ДНК- провируса и последующая трансляция белков вируса, наработка компонентов вируса и формирование новых вирионов, их выход из клетки, сопровождающийся цитопатическим эффектом для клетки - мишени.
ДНК - провирус может длительно находиться в неактивном состоянии (персистентное инфицирование). В этот период вирус можно выявить только с помощью ПЦР. Активация транскрипции особым ядерным фактором в результате действия иммунокомпетентных клеток или микробных антигенов приводит к продуктивной фазе - активному размножению ВИЧ. Факторы экспрессии генов ВИЧ - специфические антигены (прежде всего - герпес - вирусы), неспецифические митогены (фитогемагглютинин), цитокины (фактор некроза опухолей, интерфероны, гамма - интерферон), бактериальные иммуномодуляторы (фосфолипиды сальмонелл), глюкокортикостероиды.
Антигенные свойства ВИЧ.
Функция обратной транскриптазы не подвергается контролю, что обусловливает высокую частоту генетических ошибок при репликации и мутации структурных белков вируса. С учетом частоты изменчивости ни один ВИЧ не производит при репликации вирион, в точности соответствующий родительскому. Высокая генетическая изменчивость реализуется в вариабельности антигенных и биологических свойств ВИЧ. Высокой изменчивостью характеризуется продукт гена env - оболочечный белок gp120, особенно петлеобразный V3 - домен (из 35 аминокислот), к которому образуется до 90-95% всех вируснейтрализующих антител.
ВИЧ-1 и ВИЧ-2 имеют существенные отличия в строении, гомология первичной структуры геномов составляет только около 42%, перекрестного иммунитета между этими вирусами нет. Антигенными свойствами обладают все основные структурные элементы вириона, прежде всего - белки. Исключительная генетическая и антигенная изменчивость позволяет вирусу выживать в инфицированном организме.
Наряду с генетическими особенностями, ВИЧ-1 имеет фенотипические различия по ряду свойств - эффективности репликации, характеру цитопатического действия и способности образовывать синцитий (этот признак связан с вирулентностью), преимущественному тропизму к клеткам - моноцитотропные изоляты (начальные этапы болезни) и лимфотропные изоляты (разгар болезни).
Патогенез СПИДа.
Рецептором для ВИЧ является дифференцировочный антиген CD4, имеющий гомологичные участки с иммуноглобулинами и белком gp120 ВИЧ. Расположенный на мембранах Т- хелперов и Т- индукторов, рецептор CD4 в комплексе с белками HLA II класса выполняет функцию распознавания антигенов. Фиксация вируса через gp120 ВИЧ-1 (или gp105 ВИЧ-2) с мембранным рецептором CD4 блокирует основную функцию этих иммунокомпетентных клеток - восприятие сигналов от антиген- представляющих клеток. Последующая репликация вируса ведет к гибели этих клеток и выпадению их функций, т.е. к развитию иммунодефицита. Чем активнее CD4+ клетки, тем активнее процесс репродукции вируса. ВИЧ угнетает преимущественно Т- хелперы -1 (связанные с многими цитокинами клеточного иммунитета), что способствует развитию вирусных инфекций и опухолей.
Сродство вирусного gp120 (gp105 в случае ВИЧ-2) к этому рецептору определяет высокую избирательность поражения клеток. В патологический процесс вовлекаются в первую очередь и в наибольшей степени CD4+ лимфоциты, моноциты крови и макрофаги тканей, дендритные клетки крови, лимфоузлов, селезенки, кожи, альвеолярные и интерстициальные макрофаги легких, микроглия и другие клетки нервной системы, имеющие CD4+ рецепторы. Поражаются также В- и 0- лимфоциты, ретикулярные клетки, эпителиальные клетки кишечника. Большое значение в распространении ВИЧ и длительному сохранению в организме придают клеткам Лангерганса.
В патогенезе ВИЧ - инфекции большое значение имеют механизмы иммунного повреждения. Наличие сходных участков в структуре белков gp120, HLA класса II и CD4 - рецепторов определяет перекрестное реагирование антител к ВИЧ с этими структурами с рядом патологических эффектов (блокада кооперации CD4+ лимфоцитов и HLA класса II, неадекватная стимуляция CD4+ клеток и др.).
Поражение иммунной системы при ВИЧ - инфекции носит системный характер, проявляясь глубокой супрессией Т- и В- звеньев иммунитета. Происходят изменения гиперчувствительности немедленного и замедленного типа, гуморального иммунитета и факторов неспецифической защиты. Наряду с дефицитом CD4+ лимфоцитов в динамике болезни нарастает функциональная недостаточность CD8+ лимфоцитов, NK клеток, нейтрофилов. Нарушение иммунного статуса проявляется рядом синдромов - инфекционным, аллергическим, аутоиммунным, лимфопролиферативным.
Манифестный синдром приобретенного иммунодефицита (СПИД) проявляется в трех основных клинических формах: нейроСПИД, онкоСПИД, инфектоСПИД (оппортунистические инфекции). Это зависит от путей внедрения ВИЧ, его преимущественного тропизма к CD4 Т- лимфоцитам или макрофагам, наличия кофакторов (цитомегаловирус, вирус Эпштейна- Барр), дозы инфекта, иммунного статуса организма и др.
В динамике ВИЧ- инфекции можно выделить следующие основные стадии: заражение, латентный период, появление лабораторных признаков инфекции, первичная клиника острой вирусной (ретровирусной) инфекции (эта стадия может отсутствовать), клинический СПИД (иммунодефицит плюс индикаторные болезни). Особое значение имеет выявление лабораторных признаков ВИЧ- инфекции.
Можно выделить три типа инкубации:
- вирусологическую ( от инфицирования до определения в крови вируса или его антигенов) - в среднем 2 - 4 недели;
- серологическую (от заражения до сероконверсии - появления положительных серологических результатов) - в среднем 8 - 12 недель;
- СПИД - инкубацию (равно или более 10 лет). Безусловный иммунологический критерий СПИДа - снижение CD4+ лимфоцитов до 200 клеток в микролитре.
Лабораторная диагностика.
Лабораторная диагностика ВИЧ - инфекции методически базируется на ИФА, иммуноблоте и ПЦР. Основными ее направлениями являются:
- выявление антител к ВИЧ;
- выявление ВИЧ или его антигенов;
- определение изменений в иммунном статусе.
Для выявления антител применяют ИФА с различными тест - системами (лизатными, рекомбинантными, пептидными антигенами ВИЧ-1 и ВИЧ-2). Основная проблема - ложноположительные результаты (перекрестная реактивность gp120, CD4+ рецепторов, белков HLA II класса и др.). Поэтому исследования в ИФА проводят как правило с использованием параллельно нескольких различных тест - систем.
Иммуноблот чаще применяют как подтверждающий тест для выявления антител к отдельным белкам ВИЧ. Антитела к основным внутренним белкам (р17, р24) обнаруживают у 70% инфицированных и примерно у половины больных СПИДом. В иммуноблоте чаще всего выявляются антитела к gp41 (до 85%) и gp160 (до 100%).
В ранние сроки используют выявление в ИФА антигена р24. Наиболее чувствительным методом выявления ВИЧ является ПЦР - диагностика.
Основным клинико - лабораторным показателем диагностики СПИДа у ВИЧ - инфицированных является определение количества CD4+ лимфоцитов. Уровень ниже 200 клеток/мкл является основным критерием СПИДа.
Лечение является одним из наиболее актуальных и до настоящего времени не решенных проблем ВИЧ - инфекции. Теоретически наиболее оправдано применение препаратов, ингибирующих обратную транскрипцию - зидовудин, азидотимидин, диданозин, ставудин и др. Вакцины против ВИЧ находятся в стадии разработки. С учетом высокой изменчивости вируса это очень сложная задача.
Лекция №17. Сепсис. Клебсиеллы, протей, синегнойная палочка.
Сепсис относится к проблемам клинической микробиологии. Эта патология связана связана с такими понятиями как “внутрибольничные инфекции”, “госпитальные штаммы микроорганизмов”, “оппортунистические инфекции” и др. В лекции о ВИЧ - инфекции шла речь о возбудителях - оппортунистах, размножающихся и вызывающих патологию в условиях иммунокомпроментированного организма (иммунодефицита).
Госпитальные (внутрибольничные) или нозокомиальные (возникающие при уходе за больным) инфекции возникают в условиях лечебных учреждений. Возбудителями этих инфекций могут быть патогенные или условно - патогенные (оппортунисты) микроорганизмы.
В настоящее время оппортунистические инфекции приобрели большое медицинское и социальное значение (проблема ВИЧ - инфекции и снижения популяционного иммунитета).
Особенности оппортунистических инфекций - полиэтиологичность, полиорганность, склонность к генерализации процесса, малая специфичность клинических проявлений. В их развитии играют роль как экзогенные возбудители (легионеллы, токсоплазмы, листерии, микоплазмы и др.), так и представители резидентной микрофлоры (эндогенные микроорганизмы), а также состояние иммунной системы организма.
Этиология внутрибольничных инфекций многообразна - бактерии, вирусы, грибы, простейшие. Большую роль имеют грамположительные кокки - стафилококки (особенно S.aureus и S.epidermidis), стрептококки, пептококки и пептострептококки. Все большую роль играют грамотрицательные бактерии, в том числе кишечная палочка, протей, синегнойная палочка, клебсиеллы и др.
Причины возникновения госпитальных инфекций и пути передачи их возбудителей разнообразны. Это и медицинские манипуляции (инвазивные методы исследования и лечения), микробная контаминация готовых лекарственных форм, дезинфектантов и антисептиков, нарушения санитарно - эпидемиологического режима медицинских учреждений, занос различными путями патогенных возбудителей (гепатиты, сальмонеллезы, аденовирусные и энтеровирусные инфекции и мн.др.), формирование среди медицинского персонала носителей патогенных стафилококков и других микроорганизмов, отсутствие достаточного надзора за циркуляцией различных возбудителей в условиях медицинских учреждений. Однако важнейшими причинами является формирование в результате селекции госпитальных штаммов микроорганизмов, чему способствует пребывание ослабленных больных с тяжелыми формами гнойно - септических и других инфекционных заболеваний, формирование и распространение лекарственно - устойчивых штаммов патогенных и условно - патогенных микроорганизмов в результате нерациональной антибиотико- и химиотерапии, широкого распространения R- плазмид.
Основными формами госпитальных (и оппортунистических) инфекций являются: гнойно - воспалительные процессы, бактеремия, сепсис, бактериальный эндотоксический шок, дисбактериозы (дисмикробиоценозы - дисбиозы). Большингству из них дано определение в предыдущих лекциях.
Отдельно следует остановиться на сепсисе (греч. sepsis - гниение) - тяжелом генерализованном лихорадочном заболевании, обусловленном наличием очагов гнойного воспаления с непрерывным или периодическим поступлением возбудителя в кровь. Для сепсиса характерны тяжелые общие расстройства и последовательное образование новых очагов в органах и тканях. Возбудители сепсиса обычно отличаются полиорганотропностью и получают возможность в условиях иммунодефицита и снижения бактерицидных свойств крови размножаться в кровеносной и лимфатической системах.
Формами сепсиса являются септицемия (размножение возбудителя в кровеносной и лимфатической системах без образования гнойных очагов в органах и тканях) и септикопиемия (размножение возбудителя как в кровеносной и лимфатической системах, так и метастатическое размножение в различных органах и тканях).
Чаще всего сепсис является следствием генерализации локальных гнойных очагов. Наиболее частые возбудители сепсиса - стафилококки, стрептококки, грамотрицательные бактерии из семейств Enterobacteriaceae и Pseudomonadaceae, многие возбудители - оппортунисты. В зависимости от входных ворот, локализации первичного очага и других пречин выделяют сепсис послеродовый, отогенный, одонтогенный, послеабортный, перитонеальный, раневой, ожоговый, уросепсис, сепсис новорожденных. Выделяют также криптогенный сепсис - когда первичный очаг обнаружить не удается.
Из возбудителей, приобретающих все большее значение как причины госпитальных инфекций, остановимся на синегнойной палочке, клебсиеллах и представителей рода Proteus. Род Pseudomonas
Представители рода - прямые или изогнутые палочки, аэробы, хемоорганотрофы, подвижны (имеют полярно расположенные жгутики), кроме P.mallei, спор не образуют, оксидаза - положительны, характеризуются низкой сахаролитической активностью. Среди представителей рода - свободноживущие обитатели почвы и воды, а также патогены растений и животных. Наибольшее медицинское значение имеют следующие виды.
1.P.aeruginosa (синегнойная палочка) - один из основных возбудителей локальных и системных гнойно - воспалительных процессов в условиях медицинских стационаров.
2.Pseudomonas (Burkholderia) cepacia - широко распространенный, но менее вирулентный, чем синегнойная палочка, микроорганизм.
3. Pseudomonas (Burkholderia) mallei - возбудитель сапа - тяжелого (особо опасного) зоонозного заболевания человека и животных.
4. Pseudomonas (Burkholderia) pseudomallei - возбудитель мелиоидоза - тяжелого зооноза.
5. Определенным патогенным потенциалом в госпитальных условиях, наряду с первыми двумя видами, особенно у ослабленных больных, обладают и другие виды - P.putida, P.fluorescens, P.(Xanthomonas) maltophilia и др.
Синегнойная палочка - основной возбудитель инфекционных поражений человека, вызванных псевдомонадами. Возбудитель распространен повсеместно (вода, почва, растения, животные), встречается в норме у человека (наиболее часто - в кишечнике, на коже и слизистых). Возбудитель способен активно контаминировать медицинское оборудование, циркулировать среди персонала и пациентов. Риск развития вызываемых синегнойной палочкой инфекций возрастает у больных с нарушениями барьерных функций и факторов резистентности (опухоли, лейкозы, ожоги, операции, сахарный диабет и др.). Инфицирование синегнойной палочкой - причина 1520% внутрибольничных инфекций, 20% нозокомиальных пневмоний, трети всех поражений у урологических больных, 20-25% гнойных хирургических осложнений и бактеремий, вызываемых грамотрицательными бактериями.
Морфология - грамотрицательная прямая или слегка изогнутая палочка, подвижна, в мазках располагается одиночно, парами или короткими цепочками. Синтезирует слизь (капсульное вещество), особенно более вирулентные мукоидные штаммы.
Культуральные свойства. Является аэробом и обладает соответствующим типу дыхания набором ферментов (цитохромы, цитохромоксидаза, дегидразы). Растет с широком диапозоне рН и температур. Ограниченная потребность в питательных веществах способствует росту этого возбудителя на простых (универсальных) питательных средах - МПБ, МПА, Эндо, Левина и др. На жидких средах образует серовато - серебристую пленку. На плотных средах часто наблюдается феномен радужного лизиса. Уже к концу суток вследствие синтеза пигмента пиоцианина появляется сине - зеленое окрашивание культуры и в ряде случаев - самих сред (Мюллера - Хинтона, Мак Конки). Другие виды псевдомонад могут продуцировать пигменты желтого цвета с зеленой флюоресценцией (P.fluorescens, P.putida), красный (пиорубин), коричнево - черный (пиомеланин).
Биохимические свойства. Для синегнойной палочки характерна низкая сахаролитическая активность (окисляет только глюкозу), высокая протеолитическая активность и образование на кровяном агаре зоны бета- гемолиза. Синтезирует триметиламин, придающий культурам приятный запах жасмина. Продуцирует выработку бактериоцинов - пиоцинов. Пигмент пиоцианин обладает свойствами пиоцина, действующего на грамположительные и грамотрицательные бактерии.
Антигенные и патогенные свойства. Основные антигены синегнойной палочки - группоспецифический соматический О- антиген и типоспецифический жгутиковый Н- антиген. О- антигенный комплекс - агрегат ЛПС с белками и липидами клеточной стенки, обладает свойствами эндотоксина, является одним из главных факторов патогенности. В его состав входит белковый родовой О- антиген. ЛПС эндотоксина образует типо- или группоспецифический термостабильный О- антиген. Часть штаммов (мукоидные), имеющих капсулоподобную оболочку, обладают капсульным К- антигеном.
Синегнойная палочка обладает большим набором факторов патогенности - эндотоксином (ЛПС, аналогичный другим грамотрицательным бактериям), рядом экзотоксинов - цитотоксином, экзоэнзимом S, гемолизинами, экзотоксином А (наиболее важный, напоминает дифтерийный экзотоксин), ферментами (коллагеназой, нейраминидазой, протеазами).
Патогенез поражений. Синегнойная палочка - оппортунистический патоген, внеклеточный паразит, размножение которого связано со способностью противостоять действию защитных факторов организма. Возбудитель вызывает тяжелые гнойно - септические процессы (септицемия вызывает гибель 30-70% пациентов). Это связано с наличием разнообразных факторов патогенности и R- плазмид, обусловливающих множественную устойчивость к антибиотикам, к действию антисептиков и дезинфектантов.
Лабораторная диагностика. Свое название P.aeruginisa получила за голубовато - зеленое окрашивание отделяемого ран и перевязочного материала. Основным методом диагностики является бактериологический. Важным является обнружение пигмента пиоцианина. В жидкой среде его выявляют добавлением небольшого объема хлороформа - наблюдается сине - зеленое окрашивание. При микроскопии обнаруживают грамотрицательные палочки, пробы на цитохромоксидазу и каталазу положительны, культуры типируют в РА с О- и Н- антисыворотками. Видовая идентификация в пределах рода ведется с учетом биохимических, культуральных и антигенных свойств.
Лечение и специфическая профилактика. Специфической профилактики нет. При пищевых токсикоинфекциях и дисбактериозах кишечника, вызванных синегнойной палочкой, эффективен комплексный интести - бактериофаг, в состав которого входит псевдомонадный фаг. Из антибактериальных препаратов чаще применяют аминогликозиды, цефалоспорины и хинолоны.
Род Klebsiella.
Род Klebsiella относится к семейству энтеробактерий. Особенность представителей рода - способность образовывать капсулу. Основной вид - K.pneumoniae (с подвидами pneumoniae, ozaenae, rhinoscleromatis). Определенную роль в патологии имеют также K.oxytoca, K.planticola, K.terrigena. Вызывают оппортунистические поражения - госпитальные пневмонии, инфекции мочевыводящих путей, диареи у новорожденных. Клебсиеллы вызывают маститы, септицемии и пневмонии у животных, постоянно обнаруживаются на коже и слизистых оболочках человека и животных.
Клебсиеллы - прямые неподвижные палочки различных размеров. Факультативные анаэробы. Оксидаза - отрицательны, каталаза - положительны.
Культуральные свойства. Хорошо растут на простых питательных средах. Оптимум рН - 7,2-7,4, температуры - от +35 до 37 градусов Цельсия, хотя клебсиеллы могут расти в широком диапозоне температур. На плотных средах чаще образуют мутные слизистые колонии, на жидких средах - равномерное помутнение, иногда со слизистой пленкой на поверхности.
Антигенная структура. Выделяют капсульные (К) и соматические (О) антигены. Для серотипирования в РА используют К- антигены (безкапсульные варианты в РА не выявляют) с антиК- сыворотками. Некоторые К- антигены родственны К- антигенам стрептококков, эшерихий и сальмонелл.
Факторы патогенности. К ним относят полисахаридную капсулу (К- антиген), эндотоксин, фимбрии, сидерофорную систему (связывает ионы двухвалентного железа и снижает их содержание в тканях), термолабильные и термостабильные экзотоксины.
Клинические проявления. Для K.pneumoniae (subsp. pneumoniae) характерны госпитальные бронхиты и бронхопневмонии, долевые пневмонии, инфекции мочевыводящих путей, поражения мозговых оболочек, суставов, позвоночника, глаз, а также бактеремии и септикопиемии. Подвид ozaenae вызывает особую форму хронического атрофического ринита - озену, подвид rhinoscleromatis - хроническое гранулематозное поражение дыхательных путей.
Лабораторная диагностика. Основной метод - бактериологический. Чаще используют дифференциально - диагностическую среду К-2 (с мочевиной, рафинозой, бромтимоловым синим), на которой через сутки вырастают крупные блестящие слизистые колонии. Колонии окрашенные (желтые или зеленые). Идентификация культур - по биохимическим свойствам и в РА с К- сыворотками (определение серогруппы).
Лечение. Одна из особенностей клебсиелл - их множественная лекарственная устойчивость и развитие поражений на фоне снижения резистентности организма. Антибиотики применяют при генерализованных и вялотекущих хронических формах клебсиеллезов как правило в сочетании с препаратами, стимулирующими иммунитет. Препараты выбора - аминогликозиды и бета- лактамовые антибиотики широкого спектра действия.
Род Proteus.
Род Proteus относится к семейству энтеробактерий. Род получил название в честь сына Посейдона Протея, способного менять свой облик. Представители рода способны менять внешние проявления роста на плотных питательных средах, а также отличаются наибольшим плеоморфизмом (изменчивостью морфологии) по сравнению с другими энтеробактериями.
Протеи расщепляют тирозин, восстанавливают нитраты, оксидаза - отрицательны, каталаза - положительны. Они обитают в кишечниках многих видов позвоночных и безпозвоночных животных, почве, сточных водах, разлагающихся органических остатках. Могут вызывать инфекции мочевыводящих путей у человека, а также септические поражения у пациентов с ожогами и после хирургических вмешательств. Достаточно часто вызывают также пищевые токсикоинфекции. Наиболее часто роль в патологии имеют P.vulgaris и P.mirabilis.
Культуральные свойства. Протеи растут на простых средах в широком диапозоне температур. Оптимальная рН - 7,2-7,4, температура - от +35 до 37 градусов Цельсия. Колонии протеев в О- форме округлые, полупрозначные и выпуклые, Н- формы дают сплошной рост. Рост протеев сопровождается гнилостным запахом. Характерен феномер роения, Н- формы дают на МПА характерный ползучий рост в виде голубовато - дымчатой нежной вуали. При посеве по методу Шушкевича в конденсационную влагу свежескошенного МПА культура постепенно поднимается в виде вуали вверх по поверхности агара. На МПБ отмечают диффузное помутнение среды с густым белым осадком на дне. На среде Плоскирева формируют блестящие прозрачные желтовато- розовые колонии (с подщелачиванием и пожелтением среды вокруг колоний). В качестве сред обогащения используют среды Мюллера, Кауфмана, 5% желчный бульон.
Антигенные свойства. Как и у других энтеробактерий, у протеев имеются О- , Н- и К- антигены. Соматические О-антигены термостабильны, жгутиковые Н- антигены термолабильны. Для серологической идентификации определяют структуру О- и Н- антигенов. У протеев ОХ имеется антигенное родство с риккетсиями. Это их свойство ранее применяли для серологической диагностики риккетсиозов.
Факторы патогенности. К ним относят ЛПС клеточной стенки, способность к “роению”, фимбрии, протеазы и уреазу, гемолизины и гемагглютинины.
Лабораторная диагностика. Основной метод - бактериологический. Используют дифференциально - диагностические среды (Плоскирева), среды обогащения и МПА по методу Шушкевича. Чистую культуру идентифицируют по биохимическим свойствам, серологическая дифференциация проводится в РА живой и гретой культуры с поливалентными и моновалентными О- и Н- сыворотками. Самый простой метод определения - по феномену роения. Среди биохимических методов - просветление и побурение сред с тирозином и триптофаном характерно для бактерий группы Proteus- Providencia- Morganella. Важный дифференциальный признак протея - способность дезаминировать фенилаланин до фенилпировиноградной кислоты, в присутствии FeCl2 это приводит к окрашиванию среды в зеленый цвет. Дифференциальный признак, отличающий протеи от Providencia и Morganella - покраснение лизино - железного агара, обусловленное дезаминированием лизина и образованием кетокислот.
Лечение. При дисбактериозах кишечника, связанных с протеями (колиты), можно использовать протейный фаг и препараты, в состав которых он входит (интести- фаг, коли- протейный бактериофаг). Протеи обладают природной устойчивостью ко многим антибиотикам. При гнойно - септических заболеваниях, связанных с Proteus, применяют антибиотики выбора - ампициллин, цефалоспорины третьего поколения, фторхинолоны.
Лекция №18.Возбудители СПИД- ассоциированных заболеваний.
Классические инфекционные болезни вызывают патогенные микроорганизмы. Существует группа условно - патогенных возбудителей, способных вызывать патологический процесс у лиц с нарушениями иммунного статуса (оппортунистические возбудители). Следовательно, оппортунистические инфекции являются иммунодефицит - ассоциированными. Определенная группа возбудителей оппортунистических инфекций ассоциирует с развитой стадией ВИЧ- инфекции (СПИДом). Эти инфекции называют СПИД- ассоциированными, их можно рассматривать как суперинфекцию. Для каждого региона мира характерна своя структура СПИД - ассоциированных инфекций в зависимости от региональной структуры инфекционной патологии.
В зависимости от этиологии и патогенеза иммунодефицита манифестируют (проявляются) различные инфекции и инвазии. При дефиците гуморального иммунитета преобладают бактериальные инфекции, при преимущественном клеточном иммунодефиците - вирусные, протозойные и грибковые. Из большого числа известных оппортунистических инфекций в стандартный перечень СПИД - индикаторных включено 13 нозологических форм, относящихся к вызываемым простейшими
(Toxoplasma gondii, Isospora belli, Cryptosporidium spp., Pneumocystis carinii- пневмоцисты отнесены к бластомицетам - почкующимся дрожжеподобным грибам), грибам (Candida albicans, Cryptococcus neoformans, Histoplasma capsulatum, Coccidioides immitis), бактериями (Salmonella spp., Mycobacterium tuberculosis, Mycobacterium avium complex), вирусами (Herpes simplex, Cytomegalovirus hominis). По данным многих авторов примерно у половины больных СПИДом в клинике доминирует пневмоцистная пневмония, у четверти - саркома Капоши (предположительно вызывается вирусом герпеса человека 8 типа - ВГЧ8). Часто встречаются инфекции, вызываемые различными герпес- вирусами, кандидоз.
Обращают на себя внимание два обстоятельства:
- крайняя таксономическая разнородность СПИД- ассоциированных инфекций и инвазий ( вирусы, бактерии, простейшие, грибы);
- двуликость форм инфекционного процесса - латентное бессимптомное течение (чаще при нормальном иммунном статусе) или склонность к генерализации (при иммунодефиците).
Остановимся на характеристике наиболее важных групп возбудителей. Среди них одни из наиболее распространенных - герпес - вирусы и вызываемые ими инфекции. Вирусы семейства Herpesviridae.
Название эти вирусы получили от греч. herpes - ползучий. Относятся к ДНК - содержащим вирусам. Выделяют три подсемейства.
1.Alphaherpesvirinae характеризуются высокой цитопатической активностью, широким кругом поражаемых хозяев, часто вызывают латентное инфицирование нервных ганглиев. К ним относятся Herpes simplex virus (вирус простого герпеса - ВПГ типа1 и типа2), virus varicella- zoster- вирус ветряной оспы - опоясывающего лишая, многочисленные вирусы животных.
2.Bethaherpesvirinae характеризуют более длительным репродуктивным циклом, медленным развитием цитопатического эффекта, узким кругом хозяев. К этой группе относится цитомегаловирус человека - возбудитель цитомегаловирусной инфекции (цитомегалии).
3.Gammaherpesvirinae специфичны для Т- и В- лимфоцитов, часто обусловливая их латентную инфекцию, размножаются преимущественно в лимфобластоидных клетках, имеют узкий круг хозяев. К ним относится вирус Эпштейн- Барр (возбудитель инфекционного мононуклеоза), вирус болезни Марека (у птиц), некоторые герпес- вирусы обязьян.
Значительная часть герпесвирусов вызывает острые и латентные инфекции, а также обладает онкогенным потенциалом (ВПГ2 может вызывать аденокарциному шейки матки, вирус ЭБ- саркому Беркитта, ВПГ8- саркому Капоши и др.). Вирусы этого семейства относятся к числу наиболее распространенных патогенов человека. Антитела к ВПГ выявлены у 70-100% взрослого населения, к вирусу Эпштейн- Барр - у 95%, ЦМВ - у 60%.
Вирусы семейства герпесвирусов - ДНК - вирусы довольно крупных размеров (150-200 нм в диаметре). Группоспецифические антигены - белки нуклеокапсида, типоспецифические антигены - гликопротеины внешней оболочки (шипы). Имеют типичное строение оболочечных ДНК- вирусов - нуклеоид, капсид, оболочку. Геном представлен двунитевой молекулой ДНК. Вирусы обладают цитолитическим действием - они разрушают клетки. Как и другие оболочечные вирусы, они также наделяют зараженные клетки новыми антигенными свойствами, благодаря чему клетки становятся объектом атаки Т- киллеров.
Основные герпесвирусы.
1.ВПГ типа 1 присутствует в организме человека чаще, чем любой другой вирус. Инфицирование происходит в раннем детском возрасте (чаще через слюну), латенция вируса чаще наблюдается в тройничном нервном узле, инфекция протекает по типу рецидивов, вирус обладает дермонейротропным действием. Наиболее частая форма - herpes labialis.
2.ВПГ типа 2 передается половым путем или новорожденному при родах от больной матери, чаще вызывает генитальные поражения и герпес новорожденных, не исключается его передача со спермой. Мигрируя из первичного очага размножения в эпителии слизистых (чаще генитально - ректальный герпес), ВПГ проникает в поясничные ганглиозные узлы, где может сохраняться в латентном состоянии, иногда обусловливает кресцовый радикулит.
3.Вирус ветряной оспы (V-Z ) может вызывать высококонтагиозное инфекционное заболевание - ветряную оспу (при первичном контакте вируса с организмом), а также опоясывающий лишай (зостер), сопровождающийся воспалительной реакцией в задних корешках спинного мозга и в ганглиях и высыпанием везикул на коже по ходу зоны иннервации (ответ иммунного хозяина на реактивацию вируса в ганглиях).
4.Цитомегаловирус (ЦМВ) обладает способностью образовывать гигантские (цитомегалия) клетки, содержащие тельца включений. ЦМВ способен вызывать врожденную (внутриутробное заражение) и приобретенную цитомегалию, способен поражать различные органы и ткани, вызывать бессимптомное носительство и клинически выраженные формы инфекции. Вирус обладает выраженным иммунодепрессивным действием, способен инфицировать иммунокомпетентные клетки и латентно персистировать в них. Вирус может поражать печень, легкие, головной мозг, сердце, мочеполовые органы, слизистые оболочки желудочно - кишечного тракта. Вирус обнаружен в слюнных железах (и в слюне), почках (соответственно - в моче), материнском молоке, сперме, вагинальном содержимом, может передаваться половым путем. Длительное сохранение ЦМВ в клетках слюнных желез и выделение со слюной позволило назвать эту инфекцию “болезнью поцелуев”. Наиболее тяжелые последствия имеет внутриутробная ЦМВ инфекция (врожденная цитомегалия).
5.Вирус Эпштейн- Барр (ЭБ) вызывает в подростковом возрасте инфекционный мононуклеоз, способен вызывать хроническую ВЭБ инфекцию, лимфому Беркитта, карциному носоглотки. Вирус передается воздушно - капельным путем, реже - половым. Вирус имеет мембранный антиген (МА), ядерный антиген (EBNA), поздний антиген вирусного капсида (VCA). Вирус индуцирует появление популяции реактивных Т- лимфоцитов и атипичных мононуклеаров. Он специфически связывается с В- лимфоцитами, размножается в них, вызывая их поликлональную активацию и дифференцировку в плазматические клетки, секретирующие гетерофильные антитела с низкой активностью к ВЭБ, но реагирующие с различными антигенами, в т.ч. антигенами эритроцитов различных животных. Геном ВЭБ может сохраняться в В- лимфоцитах в латентном состоянии.
6-7. Вирусы герпеса человека типа 6 и 7 (ВГЧ6 и ВГЧ7). Оба вируса поражают лимфоциты и могут вызывать лимфопролиферативные заболевания. ВГЧ6 может вызывать В- клеточные лимфомы, ложную краснуху (острую вирусную инфекцию детей младшего возраста с розеолезной сыпью), возможно - синдром хронической усталости. ВГЧ7 может вызывать экзантемы у детей.
8.ВГЧ8 признают возможным фактором развития саркомы Капоши.
Лабораторная диагностика. Используют три основных подхода - ПЦР- диагностику, ИФА (прежде всего выявление антител класса IgM), МФА с моноклональными антителами.
Вирус краснухи.
Этот распространенный вирус относится к семейству Togaviridae, монотипному роду Rubivirus. Представляет собой небольшой сферический оболочечный РНКвирус с икосаэдральным нуклеокапсидом, заключенным в липидную оболочку. Вирус передается воздушно- капельным и вертикальным (трансплацентарная передача с развитием внутриутробной инфекции и тератогенным действием) путями. Отличается от других тогавирусов наличием нейраминидазы.
У человека вызывает острую вирусную инфекцию, чаще наблюдаемую у детей - краснуху. Наиболее опасна эта инфекция для беременных, особенно в первом триместре беременности. Чаще наблюдают пороки сердца, микроцефалию, множественные пороки развития.
Краснуха - высококонтагиозная инфекция, возможны крупные эпидемические вспышки, при выздоровлении формируется пожизненная невосприимчивость к инфекции. При иммунопрофилактике можно использовать живую аттенуированную вакцину.
Протозойные инвазии.
Простейшие - наиболее просто устроенные одноклеточные эукариотические микроорганизмы. Наиболее подробно их изучают в курсе медицинской биологии. Передвигаются простейшие либо с помощью псевдоподий, либо с помощью жгутиков или ресничек, при неблагоприятных условиях образуют цисты. Часто имеют сложный морфогенез. Являются одноклеточными представителями животного мира. Основными их типами являются споровики, жгутиковые, саркодовые и инфузории.
К СПИД- ассоциированным относят токсоплазмы (Toxoplasma gondii) и ряд простейших, вызывающих поражение желудочно - кишечного тракта, преимущественно в виде диарейного синдрома (криптоспоридии, микроспоридии и изоспоры).
Toxoplasma gondii вызывает токсоплазмоз - инвазию приобретенного или врожденного генеза, являющуюся причиной разнообразной патологии. Относится к споровикам (Sporozoa). Токсоплазма - внутриклеточный паразит. Заражение происходит алиментарным путем ооцитами или тканевыми цистами, чаще через мясные продукты, овощи. Ооциты выделяются с испражнениями основного хозяина - кошек и диких кошачьих, хорошо сохраняются в почве.
Человек - промежуточный хозяин, в котором развитие токсоплазм доходит до тканевых цист, в которых осуществляется бесполое размножение. Для острого токсоплазмоза характерно образование псевдоцист - скоплений токсоплазм в пораженных клетках и токсоплазмемия. При хроническом токсоплазмозе формируются истинные цисты.
Токсоплазмы могут поражать различные органы и ткани, чаще выделяют следующие формы: лимфаденопатическую, экзантемную, поражение центральной нервной системы, преимущественное поражение глаз (хориоретинит, увеит, иридоциклит), поражение сердечно - сосудистой системы.
Наиболее тяжелая инвазия наблюдается при заражении матери и трансплацентарно - плода в первом триместре беременности - врожденный токсоплазмоз, проявляющийся гидро- и микроцефалией, хориоретинитом, образованием кальцинатов в мозге. У больных СПИДом токсоплазмоз развивается преимущественно в виде патологии центральной нервной системы (токсоплазменный энцефалит).
Лабораторная диагностика.
1.МФА- диагностика.
2.Серологическая диагностика - РСК, РНИФ, ИФА.
Микозы.
Грибы - эукариотические одно- и многоклеточные организмы, в большинстве случаев - сапрофиты. Из многих десятков тысяч известных видов грибов патогенны для человека не более нескольких сотен видов. Они вызывают заболевания - микозы. Выделяют два типа роста грибов - дрожжевой и гифальный (плесневой). Дрожжи идентифицируют по биохимическим свойствам - утилизации углеводов и другим метаболическим особенностям, а также по данным микроскопии, плесневые грибы - по характеру колоний, типу роста, морфологии структур, обусловливающих вегетативное размножение.
Большую часть возбудителей микозов человека относится к несовершенным грибам (дейтеромицетам), половая (совершенная) стадия размножения у которых отсутствует. В зависимости от условий существования некоторые грибы обладают диморфизмом - могут быть мицелиальными (гифальными) или дрожжеподобными.
Выделяют следующие основные группы микозов.
1.Поверхностные микозы (сапрофитии).
2.Микозы кожи или ее придатков - дерматомикозы.
3.Подкожные микозы (болезни имплантации, при травме).
4.Системные микозы - респираторные, при поражении внутренних органов - висцеральные микозы.
5.Оппортунистические микозы.
Оппортунистические микозы.
Их возбудители - преимущественно сапрофиты из внешней среды (Aspergillus, Mucor) или эндоинфекция (Candida). Оппортунистические микозы чаще развиваются при иммунодефицитах (при СПИДе), при нарушениях местной резистентности (ожоги), сахарном диабете. Среди СПИД - ассоциированных микозов чаще отмечают пневмоцистоз, криптококкоз, гистоплазмоз, кандидоз, мукоромикоз.
Пневмоцистоз.
Pneumocystis carinii имеет тонкую капсулу, круглой или серповидной формы. Инфекционная форма - спорозоит, который в ткани легкого превращается в трофозоит, который размножается делением. Пневмоцистоз чаще развивается у детей в возрасте до одного года с врожденными дефектами иммунитета, у взрослых наиболее часто связан с ВИЧ- инфекцией. Пневмоциста - внеклеточный паразит со строгим тропизмом к легочной ткани, в просвете альвеол она осуществляет жизненный цикл. Основной путь передачи - воздушно - капельный (со слюной или мокротой), иногда - трансплацентарный. Основные источники (кроме человека) - овцы, собаки, грызуны.
Клинически характерна интерстициальная пневмония с поражением межальвеолярных перегородок, нарушениями дыхания и газообмена, развитием гипоксии. У лиц с тяжелыми иммунодефицитами пневмоцистоз носит фатальный характер.
Лабораторная диагностика. На питательных средах пневмоциста не растет. Используют :
- выявление возбудителя в окрашенных мазках мокроты и слизи (бронхоальвеолярный лаваж, ларингоскопия) - наиболее типичны восьмиядерные цисты (розетки), вегетативные формы выявить трудно;
- МФА - микроскопию;
- ИФА, однако необходимо учитывать частое выявление антител у здоровых лиц.
Кандидоз.
Род Candida относят к группе условно - патогенных дрожжевых грибов. Увеличение кандид связано с применением антибиотиков и развитием дисбактериозов, поскольку некоторые виды (Candida albicans, например) входят в состав нормальной микробной флоры организма человека. Обычно кандидозы возникают эндогенно как следствие дисфункций иммунной системы и метаболических нарушений (сахарный диабет, применение глюкокортикоидов и др.). Урогенитальный кандидоз передается половым путем. В настоящее время грибы рода Candida (чаще C.albicans) - одни из наиболее распространенных возбудителей оппортунистических микозов.
Candida - диморфные грибы, представленные дрожжевой фазой овальной формы и мицелиальной фазой (псевдо- и истинные гифы). Псевдомицелий (цепочки из удлиненных клеток) встречается у большинства видов, некоторые виды (C.albicans) образуют терминальные хламидоспоры.
Candida выделяют на среде Сабуро, кровяном и глюкозном агаре, пивном сусле, растительных отварах. Колонии мягкие, кремоватые, напоминают “капли майонеза”.
Диагностика - микроскопическая (в т.ч. МФА), бактериологическая, серологическая (РСК, ИФА).
Наиболее типичные клинические проявления - молочница (чаще ротовой полости), вульвовагинит, диссеминированный кандидоз.
Криптококкоз.
Из представителей рода Cryptococcus основное значение имеет C.neoformans, чаще выделяемый из птичьего помета и преимущественно попадающий в организм воздушно - пылевым путем. Возбудитель - оппортунист, развивается в организме на фоне Т- клеточного иммунодефицита. C.neoformans существует в несовершенной (дрожжевой) и совершенной (половой) фазах. В первом случае выявляют сферические клетки, окруженные слизистой капсулой, в совершенной фазе криптококк имеет гифы с большим количеством концевых базидий.
На микологических средах образует блестящие сочные колонии, на среде Сабуро - кремово- коричневые.
Капсула - основной фактор патогенности. Первичный очаг чаще образуется в легких, возможно вовлечение в процесс регионарных лимфатических узлов. Основная форма поражений при ВИЧ- инфекции - менингит. Нейротропизм криптококка объясняется отсутствием в спинномозговой жидкости антикриптококкового и фунгиостатических факторов, имеющихся в крови, наличием в спинномозговой жидкости ряда питательных веществ для размножения этого гриба. При ВИЧ- инфекции криптококк может поражать легкие, криптококкоз может также быть в глазной, кожной и диссеминированной формах.
Диагностика криптококкоза включает микологическое исследование патологического материала, цитологическое (выявляют капсулированные дрожжевые клетки) и биохимическое исследование спинномозговой жидкости, выявление антигена и антител в ликворе и сыворотке крови (ИФА).
Аспергиллез.
Плесневые грибы рода Aspergillus относятся к группе головчатых септических плесеней с преимущественно бесполым размножением посредством конидий. Для вегетативных гиф характерно наличие перегородок (септ) и ветвление. Конидии могут быть окрашены в различные цвета. Грибы этого рода широко распространены в почве, на растениях (зерновые культуры), в воде. Споры часто попадают в воздух. Основные возбудители аспергиллезов человека - A.fumigatis, A.flavus, A.niger, A.terreus. Повсеместная распространенность аспергилл делает их наиболее опасными агентами для пациентов с дефектами иммунной системы.
Клинически аспергиллезы протекают с преимущественным поражением легких, часто сопровождаются выраженными аллергическими реакциями, а у лиц с иммунодефицитом может генерализованно возникать диссеминированный аспергиллез, часто заканчивающийся летально.
Лабораторная диагностика.
1. Обнаружение мицелия и характерных органов спороношения (конидиеносцев) в биоптатах (микроскопия, в т.ч. - МФА).
2. Выделение культуры гриба. На среде Сабуро быстро образуют плоские колонии, сначала белые, слегка пушистые или бархатистые, затем в зависимости от вида принимают синеватую, коричневую, желтоватую окраску. 3. Иммуноферментный анализ.
Лекция №19. Арбовирусы. Вирусы клещевого энцефалита, ОГЛ, бешенства, хантавирусы.
Арбовирусы (от лат. arthropoda - членистоногие и англ. borne - передающийся, происходящий ) - вирусы различных таксономических групп, передающиеся позвоночным (в т.ч. человеку) через кровососущих членистоногих (комаров, клещей, блох, вшей, клопов и др.). Арбовирусы - нетаксономическое понятие. В настоящее время известно более 400 арбовирусов, относящихся преимущественно к семействам тогавирусов, флавивирусов, буньявирусов, рабдовирусов, аренавирусов, реовирусов.
Большинство арбовирусов - возбудители природноочаговых зоонозов, распространенных в определенных (эндемичных) регионах и ландшафтных зонах. Они имеют характерных (специфических) переносчиков, определяющих эколого - эпидемиологические особенности арбовирусных инфекций. Передача возбудителей у членистоногих осуществляется трансовариально и трансфазово (по этапам метафорфоза) из поколения в поколение. Арбовирусы обладают способностью размножаться при температуре тела теплокровных животных, а также в организме членистоногих, температура тела которых зависит от природных условий окружающей среды.
Клинически арбовирусные инфекции проявляются преимущественно в трех вариантах - лихорадки неясного генеза (“денге - подобной”), поражения центральной нервной системы (энцефалита), геморрагической лихорадки.
Семейство Togaviridae (тогавирусы).
Свое название тогавирусы получили от лат. toga - плащ. Это сферические оболочечные вирусы с икосаэдральным нуклеокапсидом, заключенным в липидную оболочку. Геном представлен позитивной линейной РНК.
Собственно к арбовирусам относят род Alphavirus (альфа - вирусы или арбовирусы антигенной группы А) с типовым представителем - вирусом Синдбис. Альфа - вирусы имеют родоспецифический нуклеокапсидный белок и два гликопротеида суперкапсидной оболочки - Е1 - группоспецифический гемагглютинин и Е2 - видоспецифический белок, антитела к которому обладают вируснейтрализующими свойствами.
В состав рода входят возбудители ряда экзотических для России инфекций - восточного и западного американского, венесуэльского энцефаломиелитов лошадей, лихорадок Чикунгунья, Росс- Ривер и др. В России из вирусов этой группы встречается вирус карельской лихорадки. Многие альфавирусы экологически связаны с птицами.
Семейство Flaviviridae (флавивирусы).
По некоторым классификациям относят в качестве рода Flavivirus к семейству тогавирусов, имея ряд близких к другим тогавирусам характеристик. Строение вирионов в целом аналогично альфавирусам. Имеют капсидный белок, шипы суперкапсида состоят из двух белков, один из которых является гликопротеидом ( Е1) и обладает гемагглютинирующей активностью. Гемагглютинирующие свойства альфа - и флавивирусов лучше выявлять в отношении птичьих (особенно гусиных) эритроцитов.
По данным РТГА альфавирусы разделяют на три, а флавивирусы - на четыре группы, представители которых (внутри групп) обладают перекрестной реактивностью по гемагглютинину. Отличительная особенность флавивирусов - наличие неструктурного растворимого антигена, выявляемого в РСК.
Основные группы флавивирусов (антигенный комплекс по РТГА).
1. Группа клещевого энцефалита (КЭ, ОГЛ, Лангат, Повассан и др.).
2. Группа японского энцефалита (ЯЭ, Западного Нила, Сент- Луис и др.).
3. Группа желтой лихорадки.
4. Группа лихорадки Денге.
Флавивирусы по экологическим предпочтениям условно делят на преимущественно комариные (вирусы желтой лихорадки, энцефалита Сент- Луис, японского энцефалита, лихорадки Денге) и преимущественно клещевые (вирусы КЭ, ОГЛ, Повассан, Кьясанурской лесной болезни).
Клещевой энцефалит.
Клещевой энцефалит (КЭ) - широко распространенная в лесной зоне умеренного климатического пояса Евразии природноочаговая облигатно - трансмиссивная инфекция, переносчиками которой являются преимущественно иксодовые клещи рода Ixodes. В соответствии с основным видом переносчика выделяют два основных типа вируса КЭ - восточный (переносчик таежный клещ Ixodes persulcatus) и западный (переносчик - европейский лесной клещ I.ricinus). Наиболее тяжело протекает КЭ, вызываемый восточным вариантом, наиболее тяжелое течение с высокой частотой очаговых поражение головного мозга и наибольшая летальность - на Дальнем Востоке России). Наиболее типичные формы клинического проявления - лихорадочная, менингиальная и очаговая. Очаговая, наиболее тяжелая и прогностически неблагоприятная форма может сопровождаться параличами, поражением стволовых отделов мозга с нарушениями дыхательной и сердечной деятельности.
Нозоареал КЭ связан с ареалом основных переносчиков, распространенных преимущественно в лесной зоне. Сезонность заболеваний связана с периодом активности переносчиков (максимальная - в мае - июне).
Лабораторная диагностика. Основные методы - вирусологические и серологические. Вирус КЭ можно выделить из крови и спинномозговой жидкости больных, снятых с человека переносчиков. Чаще производят внутримозговое заражение сосунков белых мышей, у которых вирус вызывает развитие параличей и парезов. Идентификация вируса осуществляется в РТГА, РСК и различных вариантах реакции нейтрализации вируса, в последние годы - в ИФА и с помощью генетических методов.
При серологической диагностике используют РТГА (группоспецифическая диагностика), РСК (более специфический тест, позволяет дифференцировать КЭ и ОГЛ), в последние годы - ИФА для выявления IgM- и IgG- антител.
Профилактика. Выделяют неспецифическую профилактику (борьба с переносчиками, использование средств индивидуальной защиты), экстренную серопрофилактику (введение в случае присасывания клещей в очагах противоклещевого иммуноглобулина), специфическую вакцинопрофилактику (использование инактивированной вакцины). Ни один из этих методов не может полностью обезопасить человека на неблагополучной (эндемичной) территории от заболевания КЭ, однако применение вакцины, а также иммуноглобулина в ранние сроки (первые сутки с момента присасывания клеща) существенно улучшает клинический прогноз.
Омская геморрагическая лихорадка (ОГЛ).
ОГЛ - природноочаговая инфекция, эндемичная для лесостепных районов Западной Сибири (преимущественно Омская и Новосибирская области). Заражение людей может происходить через присасывание иксодовых клещей Dermacentor reticulatus (клещевой тип заражения) или нетрансмиссивным путем - при прямом или опосредованном контакте с ондатрами (ондатровый тип). В последние десятилетия абсолютно преобладает ондатровый тип заражения. У больных отмечается лихорадка и геморрагический синдром (мелкоточечная сыпь, носовые, желудочно - кишечные и маточные кровотечения).
Вирус ОГЛ близок по свойствам и происхождению к вирусу КЭ. Имеются гипотезы происхождения вируса ОГЛ от вируса КЭ. Вирус ОГЛ антигенно близок вирусу КЭ, их серологическая дифференциация осуществляется в РСК с растворимыми антигенами. Вакцинация против КЭ обеспечивает перекрестный протективный иммунитет против ОГЛ.
Семейство Bunyaviridae (буньявирусы).
Семейство буньявирусов считают крупнейшим по числу входящих в него вирусов (более 250). Оно включает пять основных родов и ряд не отнесенных к основным родам вирусов. Большинство членов семейства - типичные арбовирусы (рода Bunyavirus, Phlebovirus, Nairovirus и Uukuvirus), недавно к буньявирусам отнесены также природноочаговые вирусы рода Hantavirus.
Буньявирусы - сферические оболочечные вирусы с тремя нуклеокапсидами, содержащими по три линейных фрагмента негативной РНК. Оболочка липопротеидная, имеет гликопротеидные шипы, проявляющие гемагглютинирующие свойства.
Наибольшее распространение на территории России имеют вирусы рода Hantavirus. Хантавирусы Хантаан, Добрава, Пуумала, Сеул и другие вызывают геморрагическую лихорадку с почечным синдромом (ГЛПС).
ГЛПС - тяжелое природноочаговое зоонозное нетрансмиссивное заболевание, характеризующееся лихорадкой, геморрагическим синдромом, почечной недостаточностью, возможен легочной синдром. Наиболее эпидемически активные очаги выявлены на Дальнем Востоке (восточная часть нозоареала), в Европейском части России и на Урале (западная часть нозоареала). ГЛПС - одна из наиболее распространенных и тяжелых природноочаговых инфекций.
Основной резервуар и источник заражения хантавирусами - мелкие млекопитающие (особенно - рыжие полевки, наибольшая численность которых отмечена в липовых лесах). Заражение человека может происходить при прямом или опосредованном (загрязненная экскрементами грызунов почва, пищевые продукты, вода) контакте, в том числе аэрогенным путем (вдыхание инфицированной пыли).
Лабораторная диагностика. Вирусологическая диагностика не применяется, вирус можно выявить в паренхиматозных органах с помощью МФА с моноклональными антителами. Преимущественно используют серологическую диагностику - реакцию непрямой иммунофлюоресценции (РНИФ) с инфицированными вирусом клеточными культурами, исследуют парные сыворотки в динамике инфекционного заболевания.
Лечение - преимущественно патогенетическое, направленное на основные клинические синдромы. При острой почечной недостаточности, уремии и геморрагическом нефрозонефрите необходим гемодиализ.
Семейство Rhabdoviridae (рабдовирусы).
Выделяют три рода - Vesiculovirus, Sigmavirus и Lyssavirus. Наибольшее значение имеет род лиссавирусов, к которому относится вирус бешенства.
Бешенство - летальная инфекция центральной нервной системы, связанная с попаданием рабдовируса со слюной (чаще в результате укусов) больных бешенством животных. Рабдовирусы - оболочечные РНК - вирусы пулевидной формы, геном образован негативной РНК. Вирус бешенства имеет широкий круг хозяев. Основной источник и резервуар этого лиссавируса - различные дикие плотоядные (волки, лисицы, шакалы и др.), от них происходит заражение домашних плотоядных - собак и кошек. Заболевание происходит через укус или ослюнение.
Вирус из входных ворот мигрирует по аксонам периферических нервов в базальные ганглии и центральную нервную систему, где размножается в сером веществе, поражая нейроны гиппокампа, продолговатого мозга, черепных нервов, симпатических ганглиев, центробежно - в клетках слюнных желез. Вирус вызывает дегенеративные поражения нейронов с образованием в клетках телец включений, известных как тельца Бабеша - Негри (выявляют гистологически - включения проявляют эозинофилию и при окраске методом флюоресцирующих антител).
Клиника бешенства у человека достаточно характерна. Второе название болезни - гидрофобия (водобоязнь), у человека наблюдается возбуждение, нарушения тонуса мышц ( в т.ч. затруднения глотания), галлюцинации, затем развиваются судороги и параличи. Смерть наступает от паралича сердечного или дыхательного центров.
Лабораторная диагностика. Применяют вирусологические методы - внутримозговое заражение белых мышей и других животных мозговой тканью или материалом подчелюстных слюнных желез. Выявляют тельца Бабеша - Негри (скопления вирусных нуклеокапсидов вблизи ядер клеток) окраской по Романовскому - Гимза или МФА.
Специфическая профилактика. Используется антирабическая инактивированная культуральная вакцина. Экстренная лечебно - профилактическая вакцинация проводится вакциной или вакциной в сочетании с антирабическим иммуноглобулином. Чем короче инкубационный период, тем хуже эффект специфической профилактики (наименьший инкубационный период - при укусах в голову, кисти рук). Обязательно проводят промывание и обработку ран или места попадания слюны с прижиганием спиртовым раствором йода.
Основа профилактики - борьба с бездомными собаками и кошками, вакцинация домашних животных. Проводятся попытки пероральной вакцинации (вскармливание разбросанных приманок с вакциной) диких плотоядных - основного резервуара вируса в природе.
Лекция №20.Риккетсии и риккетсиоподобные микроорганизмы.
Термин "риккетсии", введенный H. da Rocha-Lima (1916), объединяет обширную группу грамотрицательных микроорганизмов, как правило тесно связанных в своей жизнедеятельности с членистоногими. Систематическое положение риккетсий до настоящего времени нельзя считать окончательно определенным. Появляющиеся квалификационные схемы отражают процесс накопления знаний по биологии, генетике и экологии этих своеобразных микроорганизмов, имеющих ряд общих свойств:
а) риккетсии являются облигатными внутриклеточными паразитами;
б) в отличии от подавляющего большинства бактерий не способны к росту на бес-
клеточных питательных средах;
в) их биология связана с паразитизмом у членистоногих (клещи, вши, блохи);
г) они имеют ряд особенностей в строении, размножении, биохимических, генети-
ческих и иммунобиологических характеристиках;
д) вызываемые риккетсиями заболевания характеризуются своеобразием клиники и
эпидемиологии;
е) имеются специализированные методы изучения (риккетсиологические).
Учение о риккетсиях и риккетсиозах, благодаря работам многих отечественных и зарубежных исследователей, приведено в стройную систему. Важную роль в этом отношении имеют труды академика П.Ф.Здродовского и его школы.
Среди многообразных представителей микроорганизмов этой группы значительное место занимают риккетсии, апатогенные для человека и теплокровных животных, однако высокоадаптированные к членистоногим. Риккетсии и риккетсиоподобные микроорганизмы широко распространены среди различных представителей членистоногих, в том числе у вшей, блох, комаров, клопов, а также в большинстве видов иксодовых клещей. Принято считать эту группу микроорганизмов эндосимбионтами, находящимися в мутуалистических отношениях с хозяевами-членистоногими. Высокая адаптация к организму членистоногих большинства видов риккетсий, в том числе патогенных для позвоночных животных, позволяет многим исследователям рассматривать их в качестве первичных хозяев риккетсий. Вместе с тем, многие виды риккетсий патогенны для человека и животных, что определяет их медицинское и ветеринарное значение.
Таксономия. Патогенные риккетсии и близкие к ним возбудители - эндоцитобионты эукариотических клеток по существующей таксономии отнесены в царстве прокариотов к отделу Gracilicutes, классу Proteobacteria, порядку Rickettsiales, семействам Rickettsiaceae, включающей роды Rickettsia, Orientia, Ehrlichia и Bartonellaceae (род Bartonella и, вероятно, род Brucella), а род Coxiella (лихорадка Ку) относят к группе гамма- протеобактерий.
Краткая характеристика рода Rickettsia (da Rocha-Lima, 1916)
Морфология. Риккетсии- мелкие плеоморфные микроорганизмы от кокковидных до палочковидных, иногда нитевидные, однако чаще короткие палочки 0,3-0,6 х 0,82,0 мкм, у некоторых видов длиной до 4 мкм перед делением клеток. Жгутиков и капсул нет, но на электронных микрофотографиях клеток, подвергнутых минимальным лабораторным манипуляциям, обычно виден внешний слой аморфного материала. Грамотрицательные микроорганизмы, плохо окрашиваются обычными анилиновыми красителями. Удерживают основной фуксин. Применяется также модификация окраски по П.Ф.Здродовскому, который в связи с наличием большого количества липоидов рекомендовал использование карболового фуксина. При этом риккетсии окрашиваются в ярко-розовый или рубиново-красный цвет, цитоплазма клеток - в голубой, ядра - в синий.
Риккетсии имеют сходное с классическими грамотрицательными бактериями строение клетки: снаружи расположен микрокапсулярный слой толщиной 10-15 нм, обладающий антигенными свойствами, далее выявляется трехслойная мембрана клеточной стенки шириной 8-12 нм; цитоплазма образуется рибосомоподобными гранулами, между которыми обнаруживаются нити ДНК. При электронной микроскопии различают двухслойную клеточную оболочку, напоминающую по химическому составу клеточную оболочку бактерий, а также диффузно расположенный в цитоплазме ядерный материал - нуклеоид, рибосомы, РНК, ДНК. При изучении поверхностных структур риккетсий с помощью электронной микроскопии выявлены, как у многих бактерий, особого рода образования - волосовидные придатки или фимбрии. С наличием жгутикоподобных образований у R.sibirica, возможно, связана подвижность. У ряда видов риккетсий отмечают наличие вегетативных и покоящихся форм.
Химический состав. У риккетсий отмечено высокое содержание липидов (до 50%) и низкое - углеводов. По высокому содержанию нуклеиновых кислот (до12%) и наличию в составе как ДНК, так и РНК, риккетсии представляют бактериальные организмы. Сходны по химическому составу и клеточные стенки риккетсий и классических бактерий. В них выявлены диаминопимелиновая и мурамовая кислоты, белки, липиды, полисахариды. Однако у риккетсий содержится и глюкуроновая кислота, которая в оболочках бактерий обычно отсутствует. Соотношение ДНК и РНК составляет 1:6-1:8. РНК содержится в риккетсиях преимущественно в цитоплазме, а ДНК образует скопления, которые по существу и представляют нуклеоид.
О сложности химического состава риккетсий свидетельствует наличие в них витаминов (никотинамид, фолиевая кислота, биотин, рибофлавин, тиамин, пантотеновая кислота, витамины В6, В12). У риккетсий обнаружены энзимные системы, в частности трансаминазы, глютамат-оксидазная система, с помощью которых осуществляется в живой клетке хозяина автономный метаболизм этих микроорганизмов.
При изучении суммарных белков риккетсий методом электрофореза в полиакриламидном геле, в зависимости от разрешающей способности метода выявляется от 19 до 175 полипептидов, в том числе 6 из них являются основными с молекулярными массами в пределах 11х10 - 155х10 . Содержание Г + Ц в ДНК исследованных видов - 30-32,5 мол.%.
Физиология. Окисление осуществляется по циклу Кребса с образованием цитрата, CO2 и переаминированием глютаминовой кислоты в аспарагиновую, что свидетельствует об их энергетической активности. Вместе с тем, облигатный характер внутриклеточного паразитизма риккетсий требует для их развития веществ и энзимов, содержащихся в клетках хозяина. С учетом особенностей метаболизма риккетсии представляют собой бактерии, которые, приспособившись к внутриклеточному существованию, утратили в значительной степени способность к внеклеточному существованию.
Микроэкология и особенности культивирования. Экологической микронишей представителей рода Rickettsia служит цитоплазма, для ряда из них (риккетсии группы КПЛ) - ядро эукариотической клетки, где они размножаются свободно, без окружения паразитофорной вакуолью. Не культивируются в отсутствии клеток хозяина. Размножаются в клетках позвоночных и членистоногих, в эпидермальных клетках, выстилающих желточный мешок развивающегося куриного эмбриона. Хороший рост получен in vitro в клетках куриного эмбриона и в некоторых стационарных линиях клеток млекопитающих. Образуют бляшки на фибробластах куриного эмбриона. Температурный оптимум роста от 32 до 35 С. Риккетсии являются медленно растущими микроорганизмами, размножаются поперечным бинарным делением, время их генерации составляет не менее 8-9 часов.
Типовой вид - Rickettsia prowazekii da Rocha-Lima, 1916.
Схема изучения. При изучении штаммов риккетсий придерживаются общей схемы, предложенной П.Ф.Здродовским и Е.М.Голиневич (1972), которая включает:
а) изучение морфологии;
б) характеристику размножения в желточных мешках куриных эмбрионов;
в) воспроизведение экспериментальной инфекции на лабораторных животных;
г) иммунологическую характеристику в опытах перекрестного иммунитета;
д) серологический анализ антигенной структуры.
Основные группы и виды микроорганизмов рода Rickettsia
|
|
|
|
|
|
A. R.rickettsii группа
1. R.conorii
2. R.rickettsii
3. R.sibirica
B. R.massiliae группа
4. R.massiliae
5. R.rhipicephali
6. R.montanensis
C. R.akari группа
7. R.akari
8. R.australis
D. R.prowazekii группа
9. R.prowazekii
10. R.typhi
E. R.canadensis группа
11.R.canadensis
В инфекционной патологии основное значение имеют риккетсии группы сыпного тифа (R.prowazekii - возбудитель сыпного тифа и R.typhi - возбудитель крысиного сыпного тифа) и группы клещевых пятнистых лихорадок (КПЛ) -
R.rickettsii - возбудитель пятнистой лихорадки Скалистых гор (в Америке), R.conorii - возбудитель марсельской лихорадки (преимущественно в Средиземноморском регионе, а также в бассейнах Черного и Каспийского морей),
R.sibirica - возбудитель клещевого риккетсиоза или клещевого сыпного тифа (Северная и Центральная Азия, включая регионы юга Сибири и Дальнего Востока), R.akari - возбудитель осповидного (везикулезного) риккетсиоза, R.australis - возбудитель австралийского риккетсиоза, R.japonica - возбудитель японской клещевой пятнистой лихорадки. Из представителей других родов в патологии человека имеют значение Coxiella burnetii - возбудитель лихорадки Ку, Orientia tsutsugamushi - возбудитель лихорадки цуцугамуши, недавно открытые возбудители гранулоцитарного эрлихиоза (агент HGE из рода Ehrlichia) и моноцитарного эрлихиоза (Anaplasma chaffeensis), различные виды бартонелл, вызывающих широкий круг клинической патологии (от окопной лихорадки до болезни кошачьих царапин). Остановимся на некоторых группах патологии, вызываемых риккетсиями и коксиеллами, подробнее.
Сыпной тиф.
Сыпной тиф - антропоноз, при котором циркуляция возбудителя -Rickettsia prowazekii происходит в паразитарной системе, включающей человека (резервуар) и платяную вошь - переносчика. В организме вши риккетсии размножаются в эпителии кишечника, вызывая его разрушение (несовершенная адаптация). Риккетсии в высоких концентрациях содержатся в фекалиях переносчиков. Платяная вошь покидает хозяина при сыпнотифозной лихорадке, что и определяет ее роль как переносчика. Путь передачи - трансмиссивный (контаминация инфицированных фекалий вшей при расчесах).
Исторически эпидемический сыпной тиф - одна из наиболее важных эпидемических инфекций, получавшая наибольшее распространение в период войн, других социальных и природных потрясений (т.е. увеличения вшивости населения). Кроме этого с R.prowazekii связана болезнь Брилля - Цинссера - рецидив эпидемического сыпного тифа, возникающий у переболевших через десятки лет (эндогенная реактивация возбудителя).
Патологический процесс при риккетсиозах обусловлен тем, что риккетсии размножаются, главным образом, в эндотелиальных клетках сосудов, особенно мелких, и сосудорасширяющим действием риккетсиального токсина, что вызывает значительные изменения центральной нервной системы и расстройства кровообращения. Высказывается мнение о возможности не только длительной персистенции риккетсий в организме переболевшего но и, с учетом ангиотропизма риккетсий, развития различной сердечно - сосудистой патологии через годы после перенесенного риккетсиоза.
Лабораторная диагностика сыпного тифа и других риккетсиозов осуществляется преимущественно с использованием серологических методов - РСК, РНГА, РНИФ, ИФА.
Разработана и применяется живая сыпнотифозная вакцина. Однако наибольшее значение имеет борьба с педикулезом, своевременное лабораторное обследование на сыпной тиф длительно лихорадящих больных, особенно из категорий риска (завшивленные, бездомные, беженцы и др.).
Клещевой риккетсиоз.
Возбудитель - Rickettsia sibirica из группы клещевых пятнистых лихорадок. Облигатно- трансмиссивная природноочаговая инфекция, передаваемая человеку клещами преимущественно из родов Dermacentor и Haemophysalis. Природные очаги распространены в Сибири и на Дальнем Востоке России, в Казахстане, Монголии, Китае. Путь передачи - трансмиссивный (инокуляция при присасывании переносчика с инфицированной слюной). Клинически заболевание проявляется лихорадкой, первичным аффектом на месте присасывания клеща, регионарным лимфаденитом, розеолезно - папулезной полиморфной сыпью, относительной доброкачественностью течения. В отличии от сыпного тифа поражаются преимущественно сосуды кожи, а не головного мозга, деструкция эндотелиальных клеток сосудов менее выражена. В типичных случаях диагноз можно ставить клинически, из лабораторных методов чаще применяют РСК со специфическим антигеном.
Лихорадка Ку.
Свое название инфекция получила от первых букв (Qu) английского слова querry (неясный, неопределенный), т.е. - лихорадка неяного генеза. Возбудитель - Coxiella burnetii- облигатный фаголизосомальный паразит эукариотических клеток, не размножающийся на питательных средах. C.burnetii культивируют в куриных эмбрионах, культурах клеток, в биопробах на различных лабораторных животных.
Выделяют фазовые вариации коксиелл, аналогичные R- и S- формам бактерий. Возбудитель в фазе1 и фазе2 отличается по вирулентности, строению, иммуногенности и другим свойствам. Коксиеллы мельче риккетсий, способны образовывать инфраформы (менее 40 нм), легко проходящие через бактериальные фильтры, проявляют значительную устойчивость во внешней среде.
Лихорадка Ку - зооноз преимущественно сельскохозяйственных животных. Характеризуется множественностью источников (прежде всего пуховые козы, овцы, крупный рогатый скот, меньше - птицы) и факторов передачи инфекции (молоко, мясо, шкуры, вода, солома, пыль и др.). С наибольшей частотой заражение людей происходит прямо или опосредовано от сельскохозяйственных животных. Ведущее значение имеют аспирационный и контактный пути передачи, меньшее - алиментарный. С учетом высокой устойчивости возбудителя особое значение при лихорадке Ку имеет пылевая инфекция.
Клинически лихорадка Ку многолика, часто проходит под масками других инфекций (лихорадка неясного генеза, грипп, ОРЗ, бруцеллез и мн.др.). Наиболее характерны острое начало, высокая температура, лихорадка длительностью около недели или больше, гипергидроз, головные, суставные и мышечные боли, гепато- лиенальный синдром, возможно развитие интерстициальной пневмонии, осложнения в виде тромбофлебита, панкреатита. Наряду с острой инфекцией возможны рецидивы, хроническое течение с развитием эндокардита. Диагноз лихорадки Ку вследствие полиморфизма клинического течения невозможен без лабораторного подтверждения. Основной метод - РСК. У больных преобладают антитела к антигену C.burnetii фазы 2; антитела к антигену фазы 1 преобладают при формировании хронического течения. В лечении лихорадки Ку, равно как и риккетсиозов групп сыпного тифа и клещевых пятнистых лихорадок назначают преимущественно препараты тетрациклинового ряда. Разработана живая вакцина против лихорадки Ку, основные мероприятия проводятся по санации очагов лихорадки Ку среди сельскохозяйственных животных.
Лекция N 21. Возбудители новых инфекций Легионеллы.
Имеется большая группа облигатных внутриклеточных паразитов (коксиеллы, хламидии, легионеллы) и паразитов мембран эукариотических клеток (микоплазмы), которые вызывают так называемые “атипичные пневмонии”, этиологически редко расшифровываемые в настоящее время. Легионеллы занимают самостоятельную ветвь на филогенетическом древе прокариотов, относятся к семейству Legionellaceae, роду Legionella, включающему более 30 видов, многие из которых вызывает атипичные пневмонии. Наиболее частым возбудителем является L.pneumophila. Морфология. Легионеллы - грамотрицательные палочки средней величины с заостренными концами. Спор и капсул не образуют, подвижны, обитают в водной среде.
Культуральные свойства. Легионеллы - факультативные внутриклеточные паразиты. Размножаются в развивающихся куриных эмбрионах, в организме экспериментальных животных, в клеточных культурах, на специальных средах сложного состава. Рост медленный - крупные, выпуклые, блестящие колонии, часто с коричневой пигментацией появляются на 3-5-е сутки.
Антигенная структура. Легионеллы имеют соматический и жгутиковый антигены, обладающие групповой и типовой специфичностью. По антигенным свойствам выделяют 14 сероваров.
К важнейшим факторам патогенности относят эндотоксин и внутрифагоцитарный паразитизм.
Эколого - эпидемиологические особенности. Легинеллез (болезнь легионеров) - сапронозная инфекция. Экологически легионеллы относят к водным микроорганизмам - сапрофитам, для человека являющихся случайными паразитами, реализующими свою патогенность. Современные особенности бытового использования воды обусловили появление этой “новой” инфекции.Основной путь заражения - воздушно - капельный. Для распространения легионелл необходимо наличие мелкодисперсного источника распыления (кондиционеры - “болезнь кондиционеров”, душевые установки и др.). Вспышки болезни легионеров связаны преимущественно с гостиницами и больницами, различными типами их водных систем. Наибольшее значение имеют системы горячего водоснабжения (температурный диапозон от 25 до 46˚С оптимален для размножения легионелл и возникновения вспышек), системы охлаждения, водного массажа, инголяционное оборудование, природные источники теплой минеральной воды, фонтаны, мойки, зубные дрели. Основное значение имеет колонизация легионеллами соединительных узлов, резиновых прокладок. С целью снижения концентрации легионелл необходима чистка внутренних поверхностей (особенно резиновых) от биопленок, содержащих большие концентрации возбудителя. В природных водоемах для сохранения и размножения легионелл существенное значение имеет их ассоциация с сине - зелеными водорослями, почвенными и водными амебами. Легионеллы длительно сохраняются в цистах амеб, проникновение и размножение легионелл в амебах сходно с их поведением в макрофагах. В организме человека легионеллы - внутрифагоцитарные паразиты размножаются в альвеолярных и других макрофагах, нейтрофилах, моноцитах. Болезнь легионеров - острая, тяжело протекающая инфекция, поражающая легкие и другие системы.
Лабораторная диагностика. Материалом для микробиологического исследования чаще служит мокрота. Обычно используют угольно - дрожжевой агар с добавлением L - цистеина, пирофосфата железа, полимиксина и ванкомицина. Возбудитель можно выявить также с помощью иммунолюминесцентного метода, ИФА, ПЦР. Серологическая диагностика осуществляется в РНИФ и ИФА с исследованием парных сывороток в динамике инфекционного процесса, максимальный уровень антител наблюдается в период реконвалесценции.
Лечение и профилактика. Этиотропная терапия проводится антибиотиками (чаще эритромицином и рифампицином). Специфическая профилактика не разработана.
Бартонеллы.
Бартонеллы – требовательные к питательным средам грамотрицательные бактерии, которые были реклассифицированы из родов Rochalimaea (Brenner D.J. et al.,1993) и Grahamella (Birtles R.J. et al.,1995) в род Bartonella после сравнения результатов секвенирования 16S рРНК гена у входящих в него видов. Представителей семейства Bartonellaceae описывают как аэробные грамотрицательные палочковидные бактерии, являющиеся паразитами эритроцитов человека и других позвоночных, в отдельных случаях кожных и костных тканей, которых культивируют на бактериальных питательных средах (типа кровяного агара), многие из них имеют членистоногих переносчиков. Род к настоящему времени включает 14 видов, семь из них (Bartonella bacilliformis, B.quintana, B.henselae, B.elizabethae, B.clarridgeiae,
B.grahamii и B.vinsonii) являются патогенами человека, ассоциируя с возрастающим разнообразием вызываемой патологии. Основными, известными к настоящему времени клиническими формами бартонеллезов являются траншейная лихорадка, болезнь Карриона (лихорадка Оройя и перуанская бородавка), болезнь кошачьих царапин (cat scratch disease – CSD), бациллярный ангиоматоз кожи, рецидивирующая лихорадка с бактеремией, эндокардит, гепатит, хроническая лимфоаденопатия. Общими проявлениями бартонеллезных инфекций являются лихорадка, кожные поражения, лимфаденит, спленит. Первая из них (чаще называемая траншейной или окопной лихорадкой) была известна еще в средние века, ее выявляли в войсках под разными названиями. Эпидемиологически (основной переносчик – платяная вошь) эта инфекция весьма напоминала сыпной тиф и часто ему сопутствовала. Болезнь Карриона является эндемичной для северо – западных райнов Южной Америки, ее разновидность – перуанская бородавка была известна индейцам еще до открытия Америки европейцами, характеризуется намного более тяжелым течением, передается москитами. Бартонеллы генетически близки с представителями Brucella и Agrobacterium (входят вместе с ними в группу α2 Proteobacteria).
Исторически наши представления о рохалимеях (так назывались первые известные бартонеллы) были длительно связаны с возбудителем траншейной или волынской лихорадки – Rochalimaea (Bartonella) quintana, обширные эпидемии которой прошли в Европе в периоды 1 и 2 мировой войн. Заболевание распространялось через платяных вшей, при наличии педикулеза и людской скученности принимала широкое эпидемическое распространение, особенно в войсках во время войн. Ее возбудителем считается Bartonella quintana, хотя в современный период дополнительных микробиологических и генетических обследований больных траншейной лихорадкой не проводили. Во время первой мировой войны этой инфекцией преимущественно в Европе переболело около одного миллиона человек, наблюдалась заболеваемость и в период второй мировой войны. В послевоенный период заболеваемость траншейной лихорадкой значительно снизилась, одновременно интерес к ее изучению был потерян. Однако и в 90-е годы при обследовании ранее неблагополучных территорий (Западная Украина) антитела к этому возбудителю выявляли у различных возрастных групп сельского населения (Климчук, Курганова и др.). Наиболее типичными признаками траншейной лихорадки считают приступы лихорадки, у части больных отмечают сыпь, неврологическую симптоматику, головные, суставные и мышечные боли, увеличение селезенки, довольно часто отмечается анемизация. Одной из особенностей этой кровяной инфекции является возможность длительного носительства возбудителя в крови (месяцы – годы). Заражение возбудителем может передаваться через укусы инфицированных платяных вшей, при втирании экскрементов вшей в кожу при почесывании. Необходимо признать недостаточность современных представлений о распространении, эколого – эпидемиологических особенностях траншейной лихорадки, генетических характеристиках его возбудителя.
Болезнь кошачьих царапин (CSD) описана в 1950г. Debre с соавторами. Заболевание обычно связано с повреждениями кожных покровов, причиненными кошками, или бытовыми контактами с этими животными (особенно котятами). При легкой форме заболевания в патологический процесс вовлекаются кожные покровы и периферические лимфатические узлы, что манифестируется лимфаденопатией, эритематозными папулами или пустулами. Кожные поражения обычно развиваются на 310 дни после ранения и предшествуют лимфоаденопатии, возможна невысокая лихорадка. Однако возможна генерализация процесса, особенно у больных с иммунодефицитами, с осложнениями в виде нарушений центральной нервной системы, печени, селезенки, легких с формированием гранулем, микроабсцессов. В 1983г. Wear с соавторами выявили мелкие полиморфные грамотрицательные микроорганизмы в лимфатических узлах больных с диагнозом CSD. Однако этиология этого заболевания была установлена значительно позже, а возбудителем считали различные возбудители - хламидии, Afipia felis, Bartonella henselae. К примеру, в “Руководстве по зоонозам,1983“ заболевание описывают под названием доброкачественный лимфоретикулез (болезнь кошачьих царапин, небактериальный регионарный лимфаденит) и указывают, что его возбудитель впервые выделен В.И.Червонским и др.(1963) и отнесен к хламидиям (гальпровиям). В 1988г. от больных болезнью кошачьих царапин был выделен возбудитель, в дальнейшем идентифицированный как Afipia felis (Brenner D.J. et al.,1991). Однако оказалось, что хотя этот возбудитель достаточно часто выявляют у больных CSD, другой патоген является возбудителем болезни кошачьих царапин - Bartonella henselae, что было убедительно доказано с помощью серологических и генетических методов (Regnery R. et al.,1992; Anderson B. et al.,1993,1994). Этот вид бартонелл был выделен от больных CSD людей, домашних кошек и их блох.
Персистентная лихорадка, сопровождающаяся бактеремией бартонеллезной этиологии, чаще вызывается двумя видами – B.henselae и B.quintana. Заболевание часто развивается у иммунодефицитных больных, сопровождается высокой температурой, длительным течением с рецидивами, истощением, слабостью, болями различной локализации. Эпидемиология и особенности заражения больных изучены недостаточно, считается, что заболевания не связаны с платяными вшами.
Бациллярный ангиоматоз клинически выражается в формировании кожных и подкожных образований – пиогенных гранулем, подкожных узелков, гиперпигментированных уплотнений. Морфологически выявляется эпителиоидный ангиоматоз с выраженной дольчатой пролиферацией капиллярных сосудов с скоплением эндотелиальных клеток. Патологический процесс может затронуть глубокие мягкие ткани, кости, нередко – лимфатические узлы, селезенка, печень. Основным этиологическим агентом считают B.henselae, хотя у части больных выявляют B.quintana.
Наиболее драмотическим проявлением бартонеллезной инфекции является эндокардит, при котором часто необходимо хирургическое вмешательство на клапанном аппарате сердца. Эта патология чаще вызывается B.henselae и B.quintana.
Эрлихии
Род Ehrlichia относится к трибе Ehrlichiae семейства Rickettsiaceae порядка Rickettsiales и объединяет более 20 видов, подразделяющихся на 3 геногруппы: 1- Ehrlichia canis , 2- E. phagocytophila и 3- E. risticii . Экология эрлихий из первой и второй геногрупп связана с иксодовыми клещами, экология E. sennetsu - одного из представителей третьей геногруппы связана с рыбами. В последнее время благодаря широкому внедрению методов генетического анализа пересмотрена филогенетическая позиция представителей рода Ehrlichia. Вследствие этого E. phagocytophila - этологический агент (ГЭЧ) была реклассифицирована и помещена в соседний род Anaplasma трибы Ehrlichiae [7].
Эрлихии являются облигатными внутриклеточными паразитами, поражающими клетки крови и эндотелия сосудов теплокровных. Их резервуаром являются преимущественно клещи рода Ixodes, которые при кровопитании передают эрлихии своим хозяевам. Возбудители локализуются в цитоплазматических вакуолях (эндосомах) лейкоцитов и вызывают у человека острые гриппоподобные лихорадочные заболевания. По спектру поражаемых клеток человека различают возбудителей МЭЧ (поражают преимущественно моноциты периферической крови) и ГЭЧ (поражают преимущественно гранулоциты, в основном нейтрофилы).
Эрлихии – грамотрицательные кокко – бациллярные бактерии небольшого размера (в длину от 0,5 до 1,5 миллимикрон). Их лабораторное поддержание связано с культивированием на специальных линиях клеток – гистиоцитомы собак (DH82) и лейкемии человека (линия HL60).
До недавнего времени эрлихии были известны как возбудители заболеваний животных на ряде континентов, проблема эрлихиозов интересовала только ветеринарных работников. Ситуация стала меняться, когда в США был описан первый случай моноцитарного эрлихиоза человека –МЭЧ (Maeda K. et al.,1987). Гранулоцитарный эрлихиоз человека (ГЭЧ) впервые выявлен в 1991г., его этиология уточнена в 1994г.(Rikihisa Y.,1991; Bakken J.et al.,1994; Chen S.-M. et al.,1994). Проведенные в последующие годы молекулярно-генетические и клинико-эпидемиологические исследования позволили установить относительно высокую эпидемическую значимость этой группы инфекций в Америке и Европе. К настоящему времени в Европе заболеваемость МЭЧ зарегистрирована в Италии, Португалии, Швеции, Бельгии, а ГЭЧ в Испании, Италии, Франции, Словении, Швейцарии, Израиле, Дании, Болгарии, Польше, Германии, Швеции, Англии и Норвегии.
Исследования, выполненные в России в последние годы, позволили выявить Ehrlichia muris в клещах Ixodes persulcatus на территории Пермской области, где были описаны случаи заболевания людей моноцитарным эрлихиозом [1]. На территории Алтайского края на фоне роста заболеваемости клещевым риккетсиозом (КР) часть случаев серонегативного КР была верифицирована с использованием реакции непрямой иммунофлюоресценции (РНИФ) как гранулоцитарный эрлихиоз [3]. В Дальневосточном регионе серологически установлена заболеваемость людей гранулоцитарным эрлихиозом с некоторыми клиническими отличиями от описанного в Америке ГЭЧ [2].

















































Материалы на данной страницы взяты из открытых источников либо размещены пользователем в соответствии с договором-офертой сайта. Вы можете сообщить о нарушении.