Поделиться

Тема: Реализация генетической информации. Генетический код и его свойства
1. Роль нуклеиновых кислот в процессе передачи наследственной информации
Нуклеиновые кислоты выполняют важнейшие биологические функции в организме. В ДНК хранится наследственная информация о свойствах клеток и всего организма, различные виды РНК принимают участие в реализации наследственной информации через синтез белка.
Принцип реализации наследственной информации — от ДНК через РНК к белку экспрессинг (рис. 1):
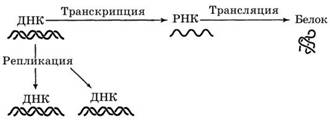 Рис. 1Основные направления внутриклеточного переноса
генетической информации (центральная догма молекулярной биологии)
Рис. 1Основные направления внутриклеточного переноса
генетической информации (центральная догма молекулярной биологии)
Расшифровка генетической информации, заключенной в молекуле ДНК, осуществляется в соответствии с центральной молекулярно-генетической догмой.
При размножении любых форм жизни происходит увеличение числа молекул ДНК. Из одной клетки, образовавшейся в результате слияния гамет, получаются тысячи, миллионы клеток тела. Каждая исходная молекула ДНК даст начало огромному числу новых молекул ДНК с сохранением в неизменном виде всех особенностей, присущих ДНК. Это происходит в процессе репликации, при которой информация, закодированная в последовательности оснований молекулы родительской ДНК, передается с максимальной точностью дочерней ДНК (рис. 2).
Репликация ДНК — единственно возможный способ увеличения числа молекул ДНК, который подсказывает сама
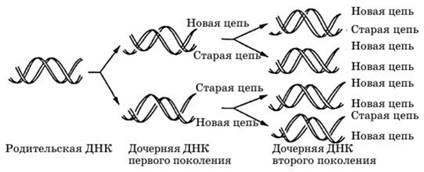
Рис.2. Репликация ДНК
структура этих молекул. С помощью фермента ДНК-полиме- разы разрываются слабые водородные связи между двумя цепями ДНК, образуются одноцепочечные цепи, которые могут раскручиваться и разделяться, после чего каждая из них служит матрицей для синтеза комплементарной (дочерней) нити — две двухцепочечные молекулы ДНК.
Таким образом, из каждой исходной молекулы ДНК получаются две копии с идентичной структурой. Уникальная структура каждой белковой молекулы, ее аминокислотный состав определяются особым процессом синтеза белка, в результате которого реализуется информация о последовательности аминокислот, записанная в ДНК.
Синтез новых нитей ДНК протекает всегда в направлении от 5' атома углерода сахара к 3' атому. Репликация имеет полу- консервативный характер — в каждой вновь образуемой молекуле ДНК одна нить происходит от родительской молекулы, а вторая синтезируется заново. Как известно, в состав ДНК входит последовательность из 4 нуклеотидов, белковые молекулы включают 20 аминокислот. Наследственная информация, закодированная с помощью нуклеотидов, переносится в первичную структуру — полипептидную цепь.
Процесс репликации нуклеиновых кислот целиком зависит от работы ряда ферментов. Установлено, что в этом процессе участвуют минимум четыре группы ферментов: ДНК-поли- меразы, РНК-полимеразы, эндонуклеазы и ДНК-лигазы:
1. Впервые ДНК-полимераза была получена в очищенном виде А. Корнбсргом из Е. coli в 1958 г. В клетках содержатся три различные формы ДНК-полимсраз, все они обладают синтезирующей активностью и способны удлинять цепи ДНК в направлении 5'—3', последовательно прибавляя по одному нуклеотиду к З'-ОН-концу цепи. Репликацию ДНК осуществляет ДНК-полимера- за, а репарацию (исправление) поврежденных участков ДНК осуществляют ДНК-полимсразы I, II.
2. РНК-полимераза — фермент, осуществляющий транскрипцию РНК.
3. Эндонуклеазы — ферменты, разрезающие двухнитсвую молекулу ДНК в местах, соответствующих последовательностям из 4-12 нуклеотидов.
4. ДНК-лигазы — ферменты, катализирующие образование фосфодиэфирной связи между 3'- и З'-концами фрагментов ДНК.
Репликация ДНК начинается с разрыва в одной из двух цепей ДНК под действием эндонуклеазы. Затем к этому месту присоединяется ДНК-полимераза и начинается непрерывный синтез нового олигонуклеотида на одной из двух родительских цепей в направлении 5'—3'. Из второй цепи родительской ДНК идет прерывистый синтез, сопровождающийся образованием фрагментов ДНК, также в направлении 5'— 3' с последующим объединением фрагментов ДНК при участии ДНК-лигаз в единую пол и нуклеотидную молекулу.
Инициация биосинтеза дочерних цепей ДНК требует предварительного синтеза на материнской ДНК затравочного олигонуклеотидного праймера со свободной гидроксильной группой. Этот олигонуклеотид, содержащий около 50 нуклеотидных остатков, синтезируется комплементарно на матрице ДНК при участии PH К-полимеразы.
Таким образом, репликация ДНК обеспечивает высочайшую точность воспроизведения генетической информации в поколениях клеток и организмов в целом, т. е. реализует одну из основных функций нуклеиновых кислот (ДНК) — сохранение генетической информации.
Бслки-фсрменты выполняют очень важную роль в обмене веществ в организме. Они являются катализаторами химических реакций, обеспечивая их в условиях гомеостаза. Ферменты — это белковые молекулы, синтезируемые живыми клетками. В каждой клетке имеются сотни различных ферментов. С их помощью осуществляются многочисленные химические реакции в организме.
Но кроме этого белковые молекулы выполняют и другие задачи. Например, они являются структурной основой как клеток, так и органов. Регуляторные белки регулируют основные процессы в клетке и в организме в целом. Сократительные белки обеспечивают движение клеток и организма. Транспортные белки переносят различные химические вещества из одного органа в другой. Защитные белки обеспечивают устойчивость организма к различным воздействиям.
2. Понятие о матричном синтезе.
Матричный синтез представляет собой образование биополимера, последовательность звеньев в котором определяется первичной структурой другой молекулы. Последняя как бы выполняет роль матрицы, "диктующей" нужный порядок сборки цепи. В живых клетках известны три биосинтетических процесса, основанных на этом механизме. Какие молекулы синтезируются на основе матрицы К реакциям матричного синтеза относят: репликацию - удвоение генетического материала; транскрипцию - синтез рибонуклеиновых кислот; трансляцию - производство белковых молекул.
3.Генетический код, его свойства
Научные исследования середины XX в. показали, что содержащаяся генетическая информация в ДНК и мРНК заключена в последовательности расположения нуклеотидов в молекулах. Установлено, что ДНК кодирует синтез белковых молекул, а последовательность оснований в нуклеотидах ДНК должна определять аминокислотную последовательность белков. Перенос информации с языка нуклеотидов на язык аминокислот осуществляется с помощью генетического кода. Генетический код — это способ записи последовательности аминокислот в белке с помощью нуклеотидов
Таблица генетического кода
|
Аминокислота |
Кодирующие триплеты — кодоны |
|
Аланин |
ГЦУ ГЦЦ ГЦА ГЦГ |
|
Аргинин |
ЦГУ ЦГЦ ЦГА ЦГГ АГА АГГ |
|
Аспарагин |
ААУ ААЦ |
|
Аспарагиновая кислота |
ГАУ ГАЦ |
|
Валин |
ГУУ ГУЦ ГУА ГУГ |
|
Г истидин |
ЦАУ ЦАЦ |
|
Глицин |
ГГУ ГГЦ ГГА ГГГ |
|
Глутамин |
ЦАА ЦАГ |
|
Глутаминовая кислота |
ГАА ГАГ |
|
Изолейцин |
АУУ АУЦ АУА |
|
Лейцин |
ЦУУ ЦУЦ ЦУА ЦУГ УУА УУГ |
|
Лизин |
ААА ААГ |
|
Метионин |
АУГ |
|
Пролин |
ЦЦУ ЦЦЦ ЦЦА ЦЦГ |
|
Серин |
УЦУ УЦЦ УЦА УЦГ АГУ АГЦ |
|
Тирозин |
УАУ УАЦ |
|
Треонин |
АЦУ АЦЦ АЦА АЦГ |
|
Триптофан |
УГГ |
|
Фенилаланин |
УУ УУЦ |
|
Цистеин |
УГУ УГЦ |
|
Знаки препинания |
УГА УАГ УАА |
В 1965 г. был расшифрован полностью весь генетический код. Из 64 кодонов 3 кодона — УАГ, УАА, УГА нс кодируют аминокислот.
Расшифровка генетического кода — одно из великих достижений науки. Первые результаты по изучению генетического кода были доложены в 1961 г. на Биохимическом конгрессе Ф. Криком и С. Бреннером, которые изучали различные формы фага Т4, получив мутации, вызванные добавлением или выпадением оснований. Это дало им возможность предположить, что одну аминокислоту может кодировать последовательность из 3 нуклеотидов (кодон, или триплет). При этом один кодон следует за другим без перерывов. Они установили также, что одну аминокислоту могут кодировать несколько триплетов. Носителем генетической информации является ДНК, но так как непосредственное участие в синтезе белка принимает мРНК — копия одной из нитей ДНК, то генетический код записан на «языке» РНК.
Расшифровка генетического кода была завершена в 1966 г., после разработки системы бесклеточного синтеза. В экспериментах М. Нирснберг и Г. Маттена в качестве мРНК в такой системе использовали нуклеиновую кислоту, состоявшую только из урациловых нуклеотидов. В результате они получили белок, содержавший только один вид аминокислоты — фенилаланин (полифенилаланин), триплет УУУ кодирует аминокислоту фенилаланин.
Генетический код обозначается четырьмя начальными буквами названий четырех типов нуклеотидов, в состав которых входят четыре разных основания: А — аденин, Т — тимин, Ц — цитозин, Г — гуанин. В белках встречается 20 различных аминокислот, длина «слова», определяющая аминокислоту, состоит из трех нуклеотидов.
Число возможных триплетов нуклеотидов — 64, так как в молекуле нуклеиновой кислоты имеются только четыре разных вида нуклеотидов, различающихся своими азотистыми основаниями, а молекула белка содержит 20 различных аминокислот. Кодирование всех аминокислот обеспечивается кодонами с тремя нуклеотидами и в этом случае возможно 43 = 64 разных трсхнуклсотидных сочетаний.
Свойства генетического кода:
1) Генетический код триплетен. Одну аминокислоту кодирует последовательность из трех нуклеотидов — триплет, или кодон.
2) Код является вырожденным — все аминокислоты, кроме метионина и триптофана, могут кодироваться нс одним, а несколькими определенными триплетами нуклеотидов. Число возможных триплетов = 64. Эти 64 триплета необходимы только для 20 аминокислот. В большинстве случаев одной аминокислоте соответствует несколько кодонов.
3) Однозначность генетического кода. Каждому кодону соответствует только одна аминокислота, т. е. триплет шифрует только одну аминокислоту'.
4) Неперекрываемость генетического кода — один нуклеотид может входить в состав только одного триплета. Ни одно из оснований, входящих в данный триплет, не является частью другого триплета. Например, последовательность мРНК, начинающаяся с нуклеотидов АУГАГЦГЦА, не считывается как АУГ/УГА/ГАГ — перекрывание по двум основаниям или АУГ/ГАГ/ГЦГ — перекрывание по одному основанию. При выпадении одного или двух нуклеотидов из цепи при считывании образуется белок, не имеющий ничего общего с тем белком, который кодировался нормальным геном.
5) Универсальность кода. Одни и те же триплеты кодируют одни и те же аминокислоты у всех организмов. Генетическая информация для всех организмов, обладающих разным уровнем организации (от ромашки до человека), кодируется одинаково.
6) Линейность кода — кодоны прочитываются последовательно в направлении закодированной записи от 5'-кон- ца к З'-концу.
7) Непрерывность кода — триплеты в ДНК следуют один за другим без перерывов.
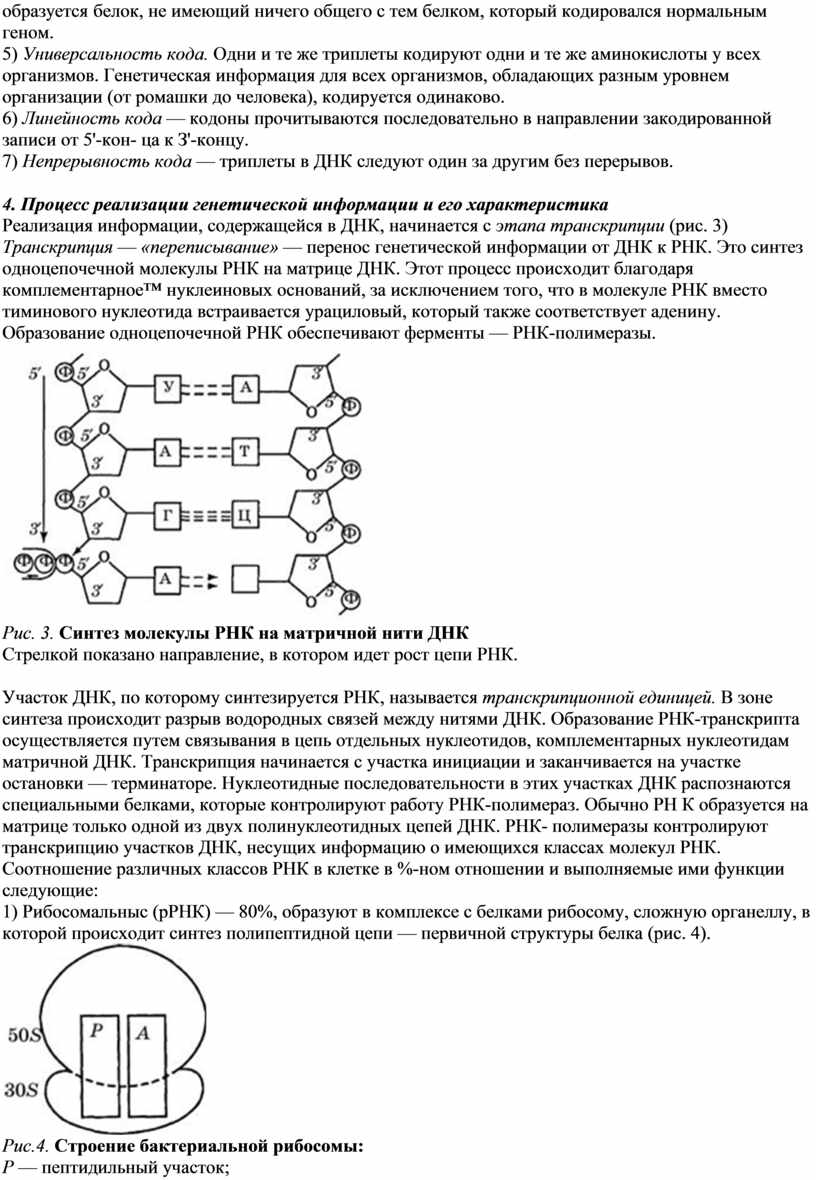
4. Процесс реализации генетической информации и его характеристика
Реализация информации, содержащейся в ДНК, начинается с этапа транскрипции (рис. 3)
Транскрипция — «переписывание» — перенос генетической информации от ДНК к РНК. Это синтез одноцепочечной молекулы РНК на матрице ДНК. Этот процесс происходит благодаря комплементарное™ нуклеиновых оснований, за исключением того, что в молекуле РНК вместо тиминового нуклеотида встраивается урациловый, который также соответствует аденину.
Образование одноцепочечной РНК обеспечивают ферменты — РНК-полимеразы.
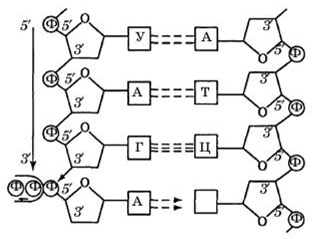
Рис. 3. Синтез молекулы РНК на матричной нити ДНК
Стрелкой показано направление, в котором идет рост цепи РНК.
Участок ДНК, по которому синтезируется РНК, называется транскрипционной единицей. В зоне синтеза происходит разрыв водородных связей между нитями ДНК. Образование РНК-транскрипта осуществляется путем связывания в цепь отдельных нуклеотидов, комплементарных нуклеотидам матричной ДНК. Транскрипция начинается с участка инициации и заканчивается на участке остановки — терминаторе. Нуклеотидные последовательности в этих участках ДНК распознаются специальными белками, которые контролируют работу РНК-полимераз. Обычно PH К образуется на матрице только одной из двух полинуклеотидных цепей ДНК. РНК- полимеразы контролируют транскрипцию участков ДНК, несущих информацию о имеющихся классах молекул РНК.
Соотношение различных классов РНК в клетке в %-ном отношении и выполняемые ими функции следующие:
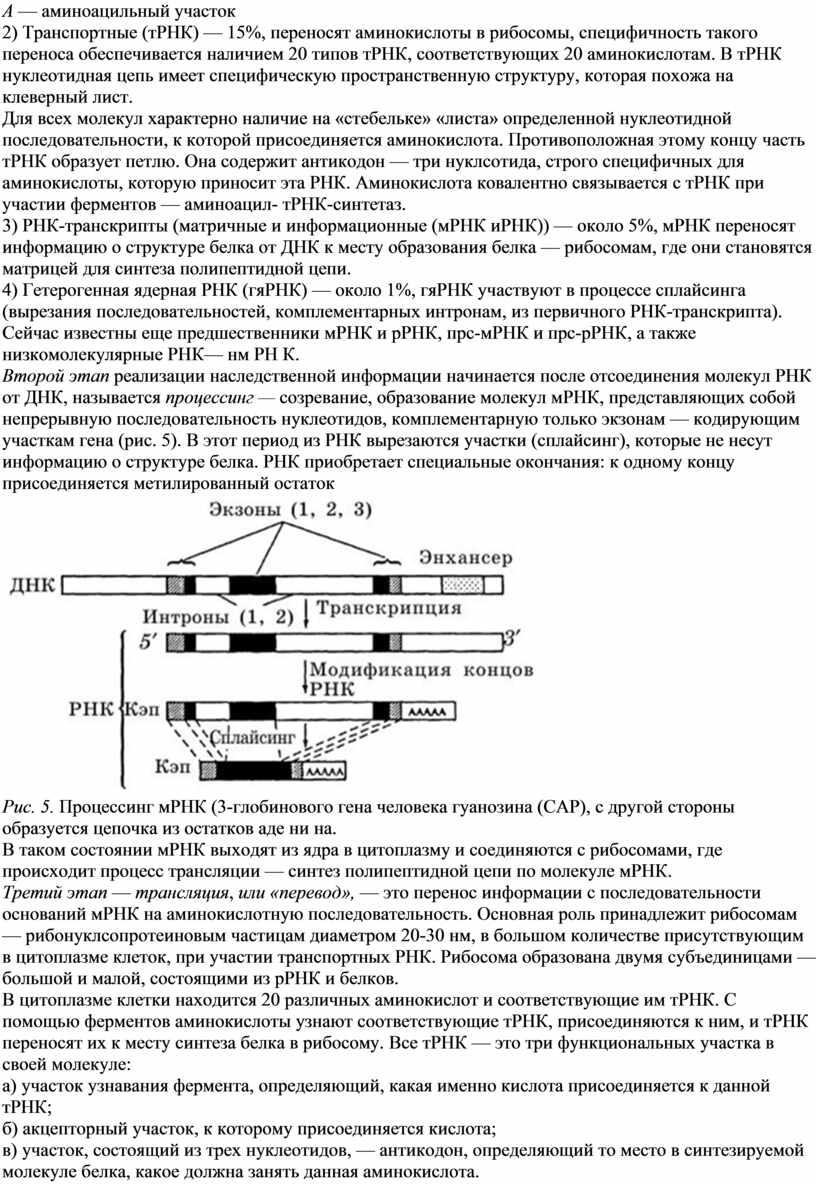
1) Рибосомальныс (рРНК) — 80%, образуют в комплексе с белками рибосому, сложную органеллу, в которой происходит синтез полипептидной цепи — первичной структуры белка (рис. 4).
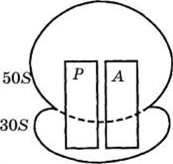
Рис.4. Строение бактериальной рибосомы:
Р — пептидильный участок;
А — аминоацильный участок
2) Транспортные (тРНК) — 15%, переносят аминокислоты в рибосомы, специфичность такого переноса обеспечивается наличием 20 типов тРНК, соответствующих 20 аминокислотам. В тРНК нуклеотидная цепь имеет специфическую пространственную структуру, которая похожа на клеверный лист.
Для всех молекул характерно наличие на «стебельке» «листа» определенной нуклеотидной последовательности, к которой присоединяется аминокислота. Противоположная этому концу часть тРНК образует петлю. Она содержит антикодон — три нуклсотида, строго специфичных для аминокислоты, которую приносит эта РНК. Аминокислота ковалентно связывается с тРНК при участии ферментов — аминоацил- тРНК-синтетаз.
3) РНК-транскрипты (матричные и информационные (мРНК иРНК)) — около 5%, мРНК переносят информацию о структуре белка от ДНК к месту образования белка — рибосомам, где они становятся матрицей для синтеза полипептидной цепи.
4) Гетерогенная ядерная РНК (гяРНК) — около 1%, гяРНК участвуют в процессе сплайсинга (вырезания последовательностей, комплементарных интронам, из первичного РНК-транскрипта).
Сейчас известны еще предшественники мРНК и рРНК, прс-мРНК и прс-рРНК, а также низкомолекулярные РНК— нм PH К.
Второй этап реализации наследственной информации начинается после отсоединения молекул РНК от ДНК, называется процессинг — созревание, образование молекул мРНК, представляющих собой непрерывную последовательность нуклеотидов, комплементарную только экзонам — кодирующим участкам гена (рис. 5). В этот период из РНК вырезаются участки (сплайсинг), которые не несут информацию о структуре белка. РНК приобретает специальные окончания: к одному концу присоединяется метилированный остаток
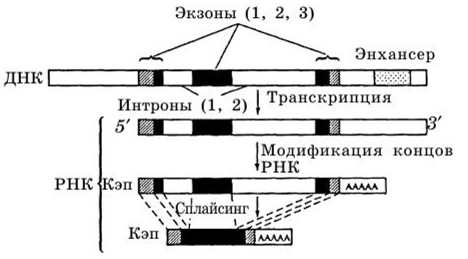
Рис. 5. Процессинг мРНК (3-глобинового гена человека гуанозина (САР), с другой стороны образуется цепочка из остатков аде ни на.
В таком состоянии мРНК выходят из ядра в цитоплазму и соединяются с рибосомами, где происходит процесс трансляции — синтез полипептидной цепи по молекуле мРНК.
Третий этап — трансляция, или «перевод», — это перенос информации с последовательности оснований мРНК на аминокислотную последовательность. Основная роль принадлежит рибосомам — рибонуклсопротеиновым частицам диаметром 20-30 нм, в большом количестве присутствующим в цитоплазме клеток, при участии транспортных РНК. Рибосома образована двумя субъединицами — большой и малой, состоящими из рРНК и белков.
В цитоплазме клетки находится 20 различных аминокислот и соответствующие им тРНК. С помощью ферментов аминокислоты узнают соответствующие тРНК, присоединяются к ним, и тРНК переносят их к месту синтеза белка в рибосому. Все тРНК — это три функциональных участка в своей молекуле:
а) участок узнавания фермента, определяющий, какая именно кислота присоединяется к данной тРНК;
б) акцепторный участок, к которому присоединяется кислота;
в) участок, состоящий из трех нуклеотидов, — антикодон, определяющий то место в синтезируемой молекуле белка, какое должна занять данная аминокислота.
Акцепторный участок одинаков у всех тРНК, он имеет последовательность Ц-Ц-А. Участки узнавания и антикодоны разные у различных тРНК.
Трансляция состоит из трех стадий: инициации, элонгации и терминации.
Инициация начинается с прикрепления малой субъединицы рибосомы к мРНК на участке связывания, который содержит обычно 3 нуклеотида: АУГ. Каждой аминокислоте в мРНК соответствует определенная тройка — триплет-нуклеотидов, называемая кодоном этой аминокислоты. Кодон комплементарен триплету, образующему антикодон соответствующей тРНК. Если в рибосоме на мРНК будет кодон АУГ, то к нему подойдет тРНК с комплементарным антикодоном УАЦ. Затем туда же присоединяется тРНК, иссушая метионин, поскольку ее антикодон комплементарен последовательности АУГ на мРНК. После этого большая и малая субъединицы объединяются и образуется активная рибосома. В рибосоме, в большой ее субъединице, между аминокислотами образуются пептидные связи. В субъединице имеются 2 области: Р-участок и А-участок. При образовании рибосомы тРНК с метионином оказывается в Р-участке, а в А-участкс присоединяется следующая тРНК, антикодон которой комплементарен нуклеотидам, расположенным на мРНК следом за последовательностью АУГ. В большой субъединице рибосомы происходит соединение двух аминокислот с образованием пептидной связи с участием специального фермента — пептидил-трансфс- разы.
Затем рибосома смешается вдоль мРНК на 3 нуклеотида, и тРНК из А-участка оказывается в Р-участке. Таким образом освобождается А-участок для другой тРНК, антикодон которой будет комплементарен следующим нуклеотидам на мРНК. Так происходит считывание информации. Многократное повторение подобного процесса, включающего образование пептидной связи между аминокислотами и продвижение рибосомы вдоль мРНК на 1 участок, соответствующий трем нуклеотидам, приводит к постоянному удлинению полипептидной цепи. При этом последовательность аминокислот в этом белке четко обусловлена составом нуклеотидов в мРНК.
Стадия удлинения полипептидной цепи на этапе трансляции получила название элонгации.
При освобождении начального участка мРНК по мере продвижения рибосомы к нему опять присоединяются другие большая и малая субъединицы и образуется новая система для синтеза белка. Обычно на одной мРНК может находиться до 100 рибосом. Такая структура называется полирибосомой, или полисомой.
Образование полипсптидной цепи продолжается, пока рибосома не достигнет нуклеотидной последовательности на мРНК, которая не кодирует аминокислот, — «стоп-кодона» (УАГ, УАА, УГА), заканчивающего синтез белка. Когда в А-участке рибосомы оказывается «стоп-кодон», туда входит специальный терминирующий белок, который освобождает полипептид от соединения с рибосомой. Затем тРНК вытесняется в цитоплазму, а рибосома передвигается на следующий кодон, происходит считывание информации. Таким образом, появление стоп-кодона на рибосоме прерывает процесс трансляции. Эта стадия трансляции получила название терминации.
На следующем этапе полипептидные цепи транспортируются к специфическим органеллам клетки и модифицируются с образованием зрелого, функционально активного белка.
Таким образом, процесс передачи и реализации наследственной информации с молекулы ДНК осуществляется путем образования различных белков с участием различных типов РНК.
В настоящее время успехи молекулярной биологии достигли такого уровня, что появилась возможность определять последовательности оснований в целых генах и удалось даже расшифровать весь генетический «текст» одного организма. Это серьезная веха в развитии науки, поскольку теперь можно искусственно синтезировать целые гены, что уже нашло применение в генной инженерии.




Материалы на данной страницы взяты из открытых источников либо размещены пользователем в соответствии с договором-офертой сайта. Вы можете сообщить о нарушении.
