Ноцицепторы взаимодействуют с целым рядом тканевых веществ, возникающих в процессе повреждения, воспаления, которые сенситизируют ноцицепторы, т.е. повышают болевую чувствительность (вызывают гипералгезию), понижают пороги болевого раздражения. Такие вещества называются алгогенами. Явление сенситизации болевых рецепторов тканевыми алгогенами означает, что адаптации болевых рецепторов не бывает.
Алгогенные вещества. Биохимическая и физиологическая
характеристика.
Возбуждаются (деполяризуются) ноцицепторы при активации натриевых
каналов и потенциалзависимых кальциевых каналов, чувствительных к
соматостатину,
разнообразным агентам:
норадреналину,
дофамину,
капсаицину. Калиевые каналы, наоборот, гиперполяризуют ноцицепторы.[6]
Ноцицепторы, кроме того, взаимодействуют с целым рядом тканевых
веществ, возникающих в процессе повреждения, воспаления, которые
сенситизируют ноцицепторы, т.е. повышают болевую чувствительность
(вызывают гипералгезию), понижают пороги болевого раздражения. Такие
вещества называются алгогенами. Явление сенситизации болевых рецепторов
тканевыми алгогенами означает, что адаптации болевых рецепторов не
бывает.
К алгогенам относятся:
- простагландины группы Е (рис.1),
Простагландины ( ПГ ) представляют собой ненасыщенные жирные
кислоты с 20 углеродными атомами, окружающими скелет молекулы
простаноевой кислоты. Особый интерес в физиологии представляют
соединения серии Е.
Это биологически активные липиды,
представляющие собой
производные гипотетич. простановой кты с заместителем и двойными
связями в циклопентановом кольце и боковых цепях. По химическому
строению простагландины относятся к 20–углеродным жирным кислотам,
содержащим 5–углеродное лактонное кольцо.
Источником для образования ПГ являются ненасыщенные жирные
кислоты (НЖК), которые поступают в организм с пищей и в
фосфорилированном виде входят в структуру мембран клеток. При
нарушении структуры клеточных мембран из НЖК под воздействием
фосфолипазы А2 образуется арахидоновая кислота. К увеличениюсодержания свободных НЖК приводят различные патогенные воздействия,
вызывающие разрушение клеточных мембран. В физиологических условиях
выходу НЖК способствуют катехоламины, брадикинин, ангиотензин II и
другие факторы, активизирующие фосфолипазы. Под воздействием фермента
циклооксигеназы из арахидоновой кислоты образуются первичные
эндоперекиси (нестабильные простагландины). В последующем под влиянием
антиокислительных систем клеток к кислороду присоединяется водород с
образованием гидроксилов в кольце и боковых цепях и формируется
молекула ПГ. Простагландины быстро метаболизируются во всех тканях,
особенно в легких, печени, почках, селезенке.
В химически чистом виде впервые были получены в 1957 г.
Бергстремом и соавторами. Предшественниками простагландинов в организме
являются полиненасыщенные жирные кислоты, в частности линолевая и
арахидоновая. Из семенников и ряда других органов выделен также
специфический фермент простагландинсинтаза, катализирующая синтез
этих веществ.
Простагландины и их производные обнаружены практически во всех
клетках млекопитающих [впервые выделены из пузырьковидной
(везикулярной) железы]. Найдены также у многих других позвоночных и
беспозвоночных (напр., у птиц, лягушек, карпов, акул, крабов, коралловых
полипов, у некоторых насекомых) и в ряде растений. Их содержание в
большинстве тканей невелико (несколько мкг/г и менее). Единственный
богатый природный источник простагландиновгоргониевые кораллы
(Plexaura homomalla), в которых содержание достигает 1,52% от сухого
веса.
простагландиноподобные вещества (простаноиды),
В кораллах найдены также биологически активные
отличающиеся от
простагландинов расположением функциональных групп, например клавулон
I(ХIII) и пунагландин (XIV).[7]Рисунок 1 Химическая формула простогландина Е
- простациклин (рис.2),
Простациклин (простагландинI2)
—
высокоактивный
метаболит арахидоновой кислоты.
Вместе с простагландинами и тромбоксанами образуют подкласс
простаноидов, которые вместе с лейкотриенами, входят в класс эйкозаноидов.
Фосфолипаза A2 отщепляет арахидоновую кислоту от
Синтез:
фосфолипидов цитоплазматической мембраны.
ЦОГ катализирует превращение арахидоновой кислоты в нестабильное
соединение ПГG2, которое спонтанно превращается в ПГH2.
Простациклинсинтаза катализирет превращение ПГН2 в ПГI2
(простациклин).
Простациклин
синтезируется
преимущественно
в эндотелии кровеносных сосудов, а также поступает в кровеносное русло из
легких.
Простациклин во многом является антагонистом тромбоксана А2, так
как снижает агрегацию тромбоцитов и вызывает вазодилатацию, усиливает
антикоагулянтное действие гепарина. Его антиагрегационное действие
связано с активацией аденилатциклазы и повышением уровня цАМФ в
тромбоцитах.Рисунок 2 Химическая формула простациклина
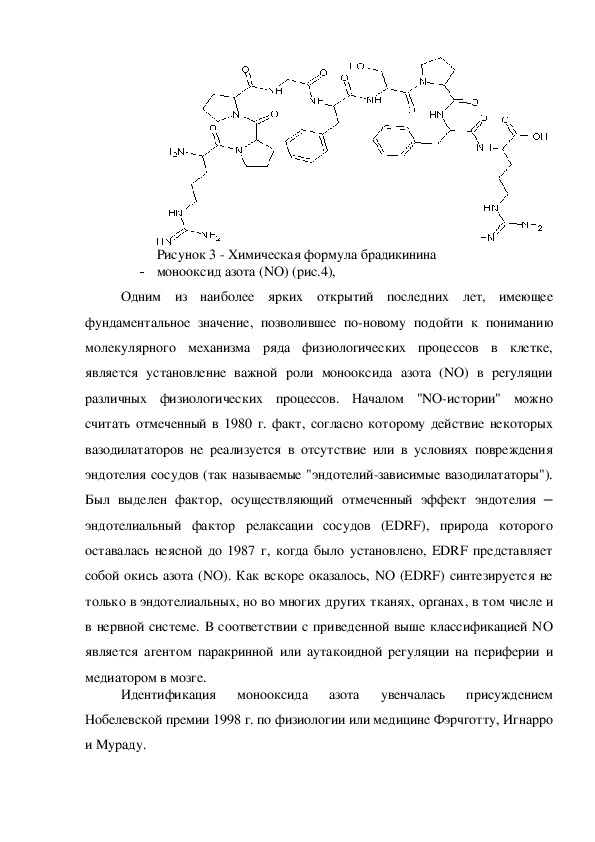
- кинины (брадикинин) (рис.3),
Брадикинин — пептид, расширяющий кровеносные сосуды и потому
снижающий артериальное давление. Ингибиторы АПФ, которые используются
для снижения артериального давления, повышают уровень брадикинина.
Брадикинин воздействует на кровеносные сосуды,
простациклин, оксид азота(II).
высвобождая
Состав брадикинина: ApгПроПроГлиФенСерПроФенApг.
Брадикинин — это физиологически и фармакологически активный
пептид из кининовой группы белков, состоящий из девяти аминокислот.
Брадикинин открыт в 1948 году коллективом бразильских учёных,
руководил которым М. Роша э Силва (англ.)русск., они же исследовали
свойства пептида, включая гипотензивные.
Пространственная структура брадикинина была теоретически
рассчитана группой учёных под руководством Станислава Галактионова в
середине 1970х годов в Минске. Экспериментальное исследование в
лаборатории Института биоорганической химии АН СССР впоследствии
подтвердило точность этих расчётов.
Метаболизм: в человеческом организме брадикинин разрушается тремя
аминопептидазой
киназами: ангиотензинпревращающим ферментом,
P, карбоксипептидазой N, которые расщепляют связи 78, 12 и 89
соответственно.Рисунок 3 Химическая формула брадикинина
- монооксид азота (NO) (рис.4),
Одним из наиболее ярких открытий последних лет, имеющее
фундаментальное значение, позволившее поновому подойти к пониманию
молекулярного механизма ряда физиологических процессов в клетке,
является установление важной роли монооксида азота (NO) в регуляции
различных физиологических процессов. Началом "NOистории" можно
считать отмеченный в 1980 г. факт, согласно которому действие некоторых
вазодилататоров не реализуется в отсутствие или в условиях повреждения
эндотелия сосудов (так называемые "эндотелийзависимые вазодилататоры").
Был выделен фактор, осуществляющий отмеченный эффект эндотелия ─
эндотелиальный фактор релаксации сосудов (EDRF), природа которого
оставалась неясной до 1987 г, когда было установлено, EDRF представляет
собой окись азота (NO). Как вскоре оказалось, NO (EDRF) синтезируется не
только в эндотелиальных, но во многих других тканях, органах, в том числе и
в нервной системе. В соответствии с приведенной выше классификацией NO
является агентом паракринной или аутакоидной регуляции на периферии и
медиатором в мозге.
Идентификация монооксида азота увенчалась присуждением
Нобелевской премии 1998 г. по физиологии или медицине Фэрчготту, Игнарро
и Мураду.Монооксид азота по своей химической природе относится к
двухатомным нейтральным молекулам. Малые размеры и отсутствие заряда
обеспечивают этой молекуле высокую проницаемость через мембраны клеток
и субклеточных структур. Наличие одного электрона с неспаренным спином
придает молекуле NO достаточно высокую реакционную способность.
Взаимодействуя с другими свободными радикалами, NO способна
образовывать ковалентные связи. Благодаря этой способности NO может как
активировать цепные свободнорадикальные реакции, так и ингибировать их.
Кроме того, NO может взаимодействовать с белками и низкомолекулярными
соединениями, содержащими в активном центре ионы переменных металлов, а
также вступает в окислительновосстановительные превращения, образуя
многочисленные азотсодержащие соединения.
NO синтезируется из Lаргинина при участии фермента NOсинтазы
(NOS) путем присоединения молекулярного кислорода к концевому атому
азота в гуанидиновой группе Lаргинина. Эта простейшая сигнальная
молекула продуцируется одним из наиболее сложных ферментов, для
проявления активности которого требуется 5 кофакторов: NADPH, FAD,
FMN, тетрагидробиоптерин и Са2+/кальмодулин. Кроме того, NOS содержит
гем. Выделены три изоформы NOсинтаз: эндотелиальная (eNOS), нейронная
(nNOS) и индуцибельная (iNOS, содержится в марофагах). Активность
различных изоформ NOсинтаз колеблется в широком пределе: нейрональная
NOсинтаза имеет максимальное значение около 300, макрофагальная – до
1000, эндотелиальная около 15 нмоль/мг/мин. Нейронная NOсинтаза имеет
молекулярный вес около 160000, тогда как макрофагальная и эндотелиальная
около 130000. Все изоформы NOS обнаруживают высокую степень гомологии
с цитохром Р450 редуктазой. Эта гомология отражает родственную природу
систем окислительного биосинтеза NO и микросомального окисления.
Удивительно, но этим сходство не ограничивается. Недавно цитохром Р450
редуктаза была обнаружена в мозге, где она является донором электронов длягемоксигеназы. В результате гемоксигеназной реакции кроме продуктов
распада гема образуется угарный газ СО, который, как оказалось, тоже играет
сигнальную роль в нервной системе.
eNOS обнаружена в эндотелии всех сосудов (включая церебральные), в
сердце (эндокарде и миокарде) и в тромбоцитах. Хотя эндотелиальную NO
синтазу относят к конститутивным изоформам, уровень ее генной экспресии
может изменяться под влиянием некоторых факторов и при патологических
состояниях.
Вторая конститутивная форма NOсинтазы
нейронная,
экспрессирована в нейронах. Клетки, содержащие nNOS, характеризуются
─
сильноразветвленной системой отростков и составляют около 2% общего
количества нейронов. Наиболее высока активность nNOS в обонятельной
луковице и мозжечке, несколько ниже ее уровень в среднем мозге, стриатуме,
коре, гиппокампе и продолговатом мозге. NO играет важную роль в регуляции
мозгового кровообращения.
Большое число NOергических нейронов находится в вегетативной
нервной системе, где они управляют важнейшими вегетативными функциями.
Нервные клетки содержащие NOсинтазу найдены по всей длине
пищеварительного тракта в т.н. энтеральной нервной системе, где они
расслабляют гладкие мышцы пищевода, желудка, кишечника и практически
всех сфинктеров (кардиального, пилорического, анального и др.). Монооксид
азота выделяется в окончаниях парасимпатических нервов, иннервирующих
половые органы, за счет чего происходит переполнение кровью пещеристых
тел и эрекция. Нейронная NOS найдена в скелетной мускулатуре, где NO
наряду с другими факторами (рН, аденозин, К+) участвует в обеспечении
рабочей гиперемии (в покое мышцы отличаются незначительным
кровоснабжением).
В отличие от эндотелиальной и нейронной NOсинтаз, которые
экспрессированы постоянно (конститутивно), индуцибельная изоформа NO
синтазы не представлена в клетках в обычных состояниях. Она играетведущую патофизиологическую роль в реализации цитотоксических функций
активированных макрофагов. Основная функция NO, вырабатываемого
─
цитотоксическое действие на клеткимишени (опухолевые, а
макрофагами,
также бактерии, вирусы и др.). Макрофагальная NOS найдена и мозге, где ее
экспрессируют особые клетки глии – т.н. микроглия.
Монооксид азота активирует растворимую форму (т.е. не связанную с
мембраной) фермента гуанилатциклазы. Это основная мишень действия NO в
нервных клетках. Гуанилатциклаза катализирует синтез цГМФ из ГТФ. цГМФ
опосредует (через изменение уровня кальция, активности цГМФзависимых
протеинкиназ, фосфодиэстераз и др.) регуляторные эффекты NO. Кроме
этого, монооксид азота является реакционноспособной молекулой и может
сам по себе, без участия каких либо ферментов модифицировать различные
белки (например, нитрозилировать по SH группам цистеина), в том числе,
белки ионных каналов. Нитрозилированные белки натриевых каналов
инактивируются и теряют способность участвовать в проведении нервных
импульсов. Недавно установлено, что нитрозилированию подвергается
аденилатциклаза в нервных клетках, что приводит к потере способности этого
фермента активироваться сигнальными молекулами. Нитрозилирование
аденилатциклазы и ионных каналов обратимо в восстанавливающих условиях,
что свидетельствует о подлинной регуляторной роли этого механизма в
управлении активностью ферментов и ионных каналов.
В нейронах гиппокампа NO синтезируется в постсинаптических
нейронах в ответ на активацию NMDAрецепторов глутаматом. Вход кальция
в клетку при этом активирует nNOS, которая синтезирует NO из Lаргинина.
Образующийся в ответ на этот импульс NO в свою очередь диффундирует из
постсинаптического нейрона в пресинаптическое окончание (ретроградно),
где оказывает облегчающее влияние на выделение нейромедиатора глутамата.
Этот механизм лежит в основе одной из наиболее важных физиологическихфункций NO в головном мозгу: NO участвует в процессах долговременной
синаптической потенциации феномена, связанного с образованием памяти.
Будучи липофильной молекулой, NO свободно переходит через
поверхностные мембраны эндотелиальных и гладкомышечных клеток сосудов,
стимулируя образование в них цГМФ, который, при посредстве
протеинкиназы G и цАМФспецифичной цГМФзависимой фосфодиэстеразы
(ФДЭ II) уменьшает концентрацию ионизированного кальция в цитоплазме.
Первый фермент активирует кальциевую помпу в саркоплазматическом
ретикулуме, второй снижает уровень цАМФ, что угнетает проводимость
кальциевых каналов плазмалеммы.
цГМФзависимая
протеинкиназа G активирует фосфатазу легких цепей миозина,
Кроме этого,
дефосфорилирующую головки миозина. Дефосфорилированный миозин
теряет способность образовывать поперечные мостики с актином. В
результате тонус гладкомышечных клеток падает, сосуды расслабляются,
сопротивление кровотоку в них понижается. Недавно обнаружен и цГМФ
независимый механизм расслабления гладких мышц, обусловленный прямым
влиянием NO
на сократительные белки
(нитрозилированием)
гладкомышечных клеток, что активирует фосфатазы, ответственных за
дефосфорилирование легких цепей миозина
Небольшие количества NO,
продуцируемые неповрежденным
эндотелием под влиянием конститутивной NOS поддерживают артериальные
сосуды в состоянии дилатации. Ингибиторы синтеза NO вызывают сужение
сосудов и существенное повышение системного артериального давления.
Недавно были проведены испытания действия ингибиторов NOсинтазы на
здоровых нормотензивных добровольцах. Оказалось, что ингибиторы
вызывают весьма значительный подъем артериального давления у людей.
Наоборот, назначение больным первичной гипертензией предшественника
образования NO в организме Lаргинина в виде пищевой добавки по 500 мг в
сутки в течение 1 месяца повышало на 50100% образование NO у этихбольных, что сопровождалось снижением артериального давления до
нормальных величин.
Малые физиологические концентрации NO (<106 М) оказывают цито и
нейропротекторное действие. Связано это с тем, что монооксид азота
является относительно долгоживущим слабым радикалом, который
эффективно перехватывает гораздо более опасные радикалы, такие как
супероксидный (O2.), гидроксильный (.OH), тиильный (напр. .GS)
липопероксильный (.LOO) и др. Однако, относительно большие концентрации
NO в тканях (>106 М), выделяющиеся при интенсивном возбуждении
нейронов либо при активации макрофагальной NOS чужеродными агентами
могут вызывать повреждение собственных клеток. В первую очередь при этом
ингибируются дыхательные ферменты митохондрий, затем происходит
фрагментация ДНК, нитрованиебелков и биогенных моноаминов, а также
перекисное окисление липидов. Во многом эти процессы обусловлены
образованием весьма токсичного пероксинитрита (ONOO). Пероксинитрит,
образующийся при взаимодействии NO с О2 дает начало большому
количеству оксидантов, играющих значительную роль в гибели нейронов.
Важным фактором, препятствующим развитию NOзависимых окислительных
процессов в клетках является СОД. При повышении концентрации NO. до 2—
4 мкМ, как это имеет место при ишемии мозга, или даже до 10 мкМ, как это
бывает в непосредственной близости от активированных макрофагов
способность супероксиддисмутазы конкурировать с NO. за супероксид анион
резко падает и синтез ONOO увеличивается.
N=O
Рисунок 4 Химическая формула монооксида азота.
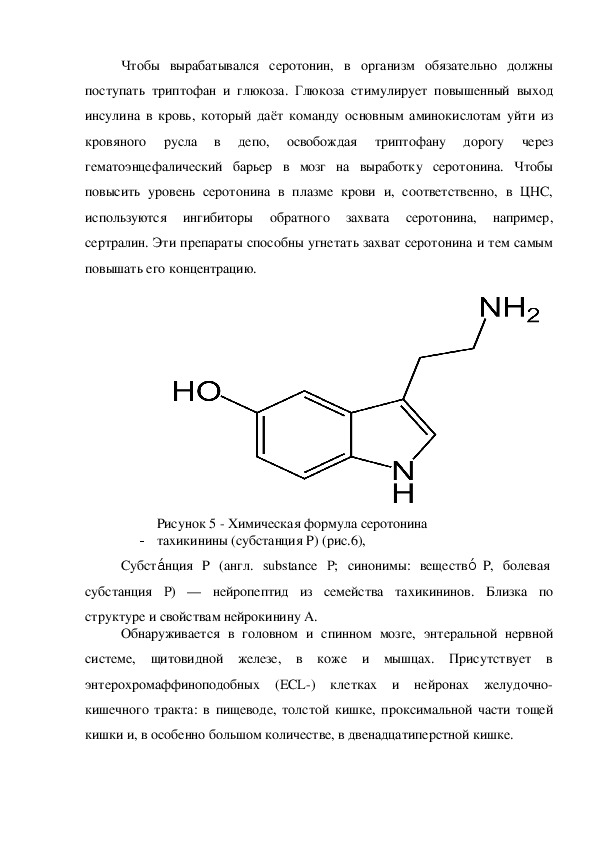
- Серотонин (рис.5),ии
Серотон н, 5гидрокситриптамин, 5НТ — один из основных
нейромедиаторов. По химическому строению серотонин относится к
биогенным аминам, классу триптаминов. Серотонин часто называют
«гормоном хорошего настроения» и «гормоном счастья».
Физиологические функции серотонина чрезвычайно многообразны.
Серотонин «руководит» очень многими функциями в организме. Например,
очень интересны исследования его влияния на проявление боли.
Доктором Виллисом доказано, что при снижении серотонина
повышается чувствительность болевой системы организма, то есть даже самое
слабое раздражение отзывается сильной болью.
Серотонин как нейромедиатор
Серотонин играет роль нейромедиатора в центральной нервной системе.
Серотонинергические нейроны группируются в стволе мозга: в варолиевом
мосту и ядрах шва. От моста идут нисходящие проекции в спинной мозг,
нейроны ядер шва дают восходящие проекции к мозжечку, лимбической
системе, базальным ганглиям, коре. При этом нейроны дорсального и
медиального ядер шва дают аксоны, различающиеся морфологически,
электрофизиологически, мишенями иннервации и чувствительностью к
некоторым нейротоксичным агентам, например, метамфетамину.
«Круговорот» серотонина
Синтезированный нейроном серотонин закачивается в везикулы. Этот
процесс является протонсопряжённым транспортом. В везикулу с помощью
протонзависимой АТФазы закачиваются ионы H+. При выходе протонов по
градиенту в везикулу поступают молекулы серотонина.
Далее, в ответ на деполяризацию терминали, серотонин выводится в
синаптическую щель. Часть его участвует в передаче нервного импульса,
воздействуя на клеточные рецепторы постсинаптической мембраны, а часть
возвращается в пресинаптический нейрон с помощью обратного захвата.
Ауторегуляция выхода серотонина обеспечивается путём активации
пресинаптических 5НТ рецепторов, запускающих каскад реакций, которые
регулируют вход ионов кальция внутрь пресинаптической терминали. Ионыкальция, в свою очередь, активируют фосфорилирование фермента 5
триптофангидроксилазы, обеспечивающей превращение триптофана в
серотонин, что приводит к усилению синтеза серотонина.
Обратный захват производится транспортером серотонина,
двенадцатидоменным белком, производящим натрийкалийсопряжённый
транспорт. Вернувшийся в клетку медиатор расщепляется с помощью
моноаминооксидазы до 5гидроксилиндолилуксусной кислоты.
Химизм транспортных систем серотонина также подобен таковым
норадреналина.
Функции серотонина
Серотонин облегчает двигательную активность, благодаря усилению
секреции субстанции Р в окончаниях сенсорных нейронов путём воздействия
на ионотропные и метаботропные рецепторы.
Серотонин наряду с дофамином играет важную роль в механизмах
гипоталамической регуляции гормональной функции гипофиза. Стимуляция
серотонинергических путей, связывающих гипоталамус с гипофизом,
вызывает увеличение секреции пролактина и некоторых других гормонов
передней доли гипофиза — действие, противоположное эффектам
стимуляции дофаминергических путей.
Серотонин также участвует в регуляции сосудистого тонуса.
Серотониновый синдром
Избыток серотонина может быть потенциально опасен, вызывая
последствия, известные как серотониновый синдром. Такая критическая
концентрация серотонина зачастую является следствием параллельного
применения антидепрессантов классов ингибиторов моноаминооксидазы и
селективных ингибиторов обратного захвата серотонина.
Серотонин как гормон
Повышение свёртываемости крови
Серотонин играет важную роль в процессах свёртывания крови.
Тромбоциты крови содержат значительные количества серотонина и обладают
способностью захватывать и накапливать серотонин из плазмы крови.Серотонин повышает функциональную активность тромбоцитов и их
склонность к агрегации и образованию тромбов. Стимулируя специфические
серотониновые рецепторы в печени, серотонин вызывает увеличение синтеза
печенью факторов свёртывания крови. Выделение серотонина из
повреждённых тканей является одним из механизмов обеспечения
свёртывания крови по месту повреждения.
Влияние на аллергические и воспалительные реакции
Серотонин участвует в процессах аллергии и воспаления. Он повышает
проницаемость сосудов, усиливает хемотаксис и миграцию лейкоцитов в очаг
воспаления, увеличивает содержание эозинофилов в крови, усиливает
дегрануляцию тучных клеток и высвобождение других медиаторов аллергии и
воспаления.
Местное (например, внутримышечное) введение экзогенного серотонина
вызывает сильную боль в месте введения. Предположительно серотонин
наряду с гистамином и простагландинами, раздражая рецепторы в тканях,
играет роль в возникновении болевой импульсации из места повреждения или
воспаления.
Влияние на пищеварение
Также большое количество серотонина производится в кишечнике.
Серотонин играет важную роль в регуляции моторики и секреции в
желудочнокишечном тракте, усиливая его перистальтику и секреторную
активность. Кроме того, серотонин играет роль фактора роста для некоторых
усиливает бактериальный
видов симбиотических микроорганизмов,
метаболизм в толстой кишке. Сами бактерии толстой кишки также вносят
некоторый вклад в секрецию серотонина кишечником, поскольку многие виды
симбиотических бактерий обладают способностью декарбоксилировать
триптофан. При дисбактериозе и ряде других заболеваний толстой кишки
продукция серотонина кишечником значительно снижается.
Массовое высвобождение серотонина из погибающих клеток слизистой
желудка и кишечника при воздействии цитотоксических химиопрепаратовявляется одной из причин возникновения тошноты и рвоты, диареи при
химиотерапии злокачественных опухолей. Аналогичное состояние бывает при
некоторых злокачественных опухолях,
серотонин.
эктопически продуцирующих
Влияние на процессы в матке
Большое содержание серотонина также отмечается в матке. Серотонин
играет роль в паракринной регуляции сократимости матки и маточных труб и
в координации родов. Продукция серотонина в миометрии возрастает за
несколько часов или дней до родов и ещё больше увеличивается
непосредственно в процессе родов. Также серотонин вовлечён в процесс
овуляции — содержание серотонина (и ряда других биологически активных
веществ) в фолликулярной жидкости увеличивается непосредственно перед
разрывом фолликула, что, повидимому, приводит к увеличению
внутрифолликулярного давления.
Влияние на половую систему
Серотонин оказывает значительное влияние на процессы возбуждения и
торможения в системе половых органов. Например, увеличение концентрации
серотонина у мужчин задерживает наступление эякуляции.
Один из гормонов удовольствия
Серотонин часто называют «гормоном счастья», он вырабатывается в
организме в моменты экстаза, его уровень повышается во время эйфории и
понижается во время депрессии. Для выработки серотонина обязательно
нужен ультрафиолет, недостаток ультрафиолета в зимнее время года и
является причиной столь распространённой сезонной депрессии.
Изменение уровня серотонина
На уровень серотонина в организме можно влиять:
-
с помощью физических упражнений, изменения ритма и глубины
дыхания
- диетами
- натуральными и химическими лекарственными препаратами
-
солнечным светом.Чтобы вырабатывался серотонин, в организм обязательно должны
поступать триптофан и глюкоза. Глюкоза стимулирует повышенный выход
инсулина в кровь, который даёт команду основным аминокислотам уйти из
кровяного русла в депо, освобождая триптофану дорогу через
гематоэнцефалический барьер в мозг на выработку серотонина. Чтобы
повысить уровень серотонина в плазме крови и, соответственно, в ЦНС,
используются ингибиторы обратного захвата серотонина, например,
сертралин. Эти препараты способны угнетать захват серотонина и тем самым
повышать его концентрацию.
-
Рисунок 5 Химическая формула серотонина
тахикинины (субстанция Р) (рис.6),
аи
Субст нция Р (англ. substance P; синонимы: веществ Р, болевая
ои
субстанция P) — нейропептид из семейства тахикининов. Близка по
структуре и свойствам нейрокинину А.
Обнаруживается в головном и спинном мозге, энтеральной нервной
системе, щитовидной железе, в коже и мышцах. Присутствует в
энтерохромаффиноподобных (ECL) клетках и нейронах желудочно
кишечного тракта: в пищеводе, толстой кишке, проксимальной части тощей
кишки и, в особенно большом количестве, в двенадцатиперстной кишке.Субстанция Р состоит из 11 аминокислотных остатков. Её структура:
ArgProLysProGlnGlnPhePheGlyLeuMet.
Субстанция Р обладает широким спектром биологической активности:
оказывает сосудорасширяющее действие, влияет на артериальное давление
крови, увеличивает капиллярную проницаемость, способствует дегрануляции
тучных клеток, является хемоаттрактантом для лейкоцитов, активирует
синтез и высвобождение медиаторов воспаления, вызывает сокращение
гладкой мускулатуры, оказывает секретогенное действие, стимулирует
высвобождение пролактина и пищеварительных гормонов.
Отвечает за передачу болевых импульсов в центральную нервную
систему. Клетки кожи голого землекопа лишены субстанции Р, поэтому голые
землекопы не чувствуют порезов и ожогов. Однако после инъекции данного
нейротрансмиттера болевые ощущения появляются.
Рисунок 6 Химическая формула субстанции Р
- цитокины,
Цитокины (cytokines) [греч. kytos — сосуд, здесь — клетка и kineo —
двигаю, побуждаю] — большая и разнообразная группа небольших по
размерам (молекулярная масса от 8 до 80 кДа) медиаторов белковой природы
— молекулпосредников («белков связи»), участвующих в межклеточной
передаче сигналов преимущественно в иммунной системе.
К цитокинам относят фактор некроза опухоли, интерфероны, ряд
интерлейкинов и др. Цитокины, которые синтезируются лимфоцитами иявляются регуляторами пролиферации и дифференцировки, в частности
гематопоэтических клеток и клеток иммунной системы, называют
лимфокинами.
Все клетки иммунной системы имеют определенные функции и
работают в четко согласованном взаимодействии, которое обеспечивается
специальными биологически активными веществами цитокинами
регуляторами иммунных реакций. Цитокины это специфические белки, с
помощью которых разнообразные клетки иммунной системы могут
обмениваться друг с другом информацией и осуществлять координацию
действий.
Набор и количества цитокинов, действующих на рецепторы клеточной
поверхности, "цитокиновая среда" представляют собой матрицу
взаимодействующих и часто меняющихся сигналов. Эти сигналы носят
сложный характер изза большого разнообразия цитокиновых рецепторов и из
за того, что каждый из цитокинов может активировать или подавлять
несколько процессов, включая свой собственный синтез и синтез других
цитокинов, а также образование и появление на поверхности клеток
цитокиновых рецепторов.
Межклеточная сигнализация в иммунной системе осуществляется путем
непосредственного контактного взаимодействия клеток или с помощью
медиаторов межклеточных взаимодействий. При изучении дифференцировки
иммунокомпетентных и гемопоэтических клеток, а также механизмов
межклеточного взаимодействия, формирующих иммунный ответ, и была
открыта большая и разнообразная группа растворимых медиаторов белковой
природы молекулпосредников ("белков связи"), участвующих в
межклеточной передаче сигналов цитокинов .
Гормоны обычно исключают из этой категории на основании
эндокринного (а не паракринного или аутокринного) характера их действия.
Вместе с гормонами и нейромедиаторами они составляют основу языкахимической сигнализации, путем которой в многоклеточном организме
регулируется морфогенез и регенерация тканей.
В положительной и отрицательной регуляции иммунного ответа им
принадлежит центральная роль. К настоящему времени у человека
обнаружено и изучено в той или иной степени, как уже упоминалось выше,
более ста цитокинов, и постоянно появляются сообщения об открытии новых.
Для некоторых получены генноинженерные аналоги. Цитокины действуют
через активацию рецепторов цитокинов.
Классификация цитокинов.
Основные группы цитокинов: Интерфероны, Интерлейкины, Фактор
некроза опухолей, Колониестимулирующиефакторы, Ростовые факторы. В
зависимости от типа клеток, продуцирующих цитокины, их делят на
монокины (ИЛ1, ФНО), синтезируемые моницитами, макрофагами, и
лимфокины (ИЛ2, ИЛ4, ИЛ5, ИЛ10 и др.), синтезируемые лимфоцитами.
Биосинтез и механизм действия питокинов.
Цитокины синтезируются клеткамипродуцентами после их
взаимодействия со стимуляторами. Молекулы стимуляторов связываются с
мембранными рецепторами клетки, вызывая тем самым ее активацию
(изменение биохимических процессов, экспрессию определенных генов, в том
числе и кодирующих цитокины), биосинтез и секрецию определенного спектра
цитокинов. Биологические эффекты цитокинов весьма разнообразны.
В ответ на адсорбцию на мембране клетокмишенейвирусных частиц
или их стимуляцию вирусной нуклеиновой кислотой, а также другими
стимуляторами интерферона секретируется мононуклеарными клетками
(моноцитами и лимфоцитами).
Особую роль в этом играют Тхелперы первого типа (Тп1). Поскольку
интерфероны проявляют неспецифическую противовирусную активность, то
они являются наиболее важным элементом естественного иммунитета
организма. Интерфероны, продуцируемые инфицированными клетками
(обычно ИНФ Р), индуцируют состояние противовирусной резистентностиокружающих клеток и организма в целом. Основное противовирусное дей
ствие интерферонов заключается ингибиции трансляции вирусной мРНК.
Интерферон связывается с соответствующим рецептором на мембране
клеток мишеней. Это связывание вызывает дерепрессию нескольких генов,
которые детерминируют вирусные ингибиторные белки, которые блокируют
трансляцию вирусной мРНК, но не мРНК клетки хозяина. Противовирусные
эффекты интерферонов а и Рг очень близки.
Он синтезируется в процессе иммунного ответа в результате
антигенспецифического распознавания и активации соответствующих клонов
Тлимфоцитов хелперов первого типа. Данная субпопуляция Тхелперов
является наиболее важным элементом клеточного иммунного ответа и
отвечает за реализацию реакций гиперчувствительности замедленного типа
(ГЗТ). Этот механизм формирования специфической резистентности особенно
важен для защиты организма от внутриклеточных патогенов, таких как
микобактерии, листерии и, безусловно, вирусы, у интерферон способен
стимулировать противомикробную активность макрофагов и естественных
киллеров.
Рецепторы цитокинов
Цитокины — гидрофильные сигнальные вещества, действие которых
опосредовано специфическими рецепторами на внешней стороне
плазматической мембраны. Связывание цитокинов с рецептором приводит
через ряд промежуточных стадий к активации транскрипции определенных
генов.
Сами цитокиновые рецепторы не обладают тирозинкиназной
активностью (за немногими исключениями). После связывания с цитокином
молекулы рецептора ассоциируют, образуя гомодимеры. Кроме того, они
могут образовывать гетеродимеры за счет ассоциации с белками
переносчиками сигнала [БПС (STP)] или стимулировать димеризацию самих
БПС.
Цитокиновые рецепторы класса I могут агрегировать с тремя типами
БПС: белками GP130, с или с. Эти вспомогательные белки сами не
β
γспособны связывать цитокины, но они осуществляют передачу сигнала на
тирозинкиназы, Одинаковые спектры биологической активности многих
цитокинов объясняются тем, различные цитокинрецепторные комплексы
могут активировать одни и те же БПС.
В качестве примера передачи сигнала от цитокинов, рецептор ИЛ6 (IL
6) после связывания с лигандом стимулирует димеризацию GP130. Димер
мембранного белка GP130 связывает и активирует цитоплазматическую
тирозинкиназу ЯКсемейства (Янускиназы, имеющие два активных центра).
Янускиназы фосфорилируют цитокиновые рецепторы, БПС и
различные цитоплазматические белки, которые осуществляют дальнейшую
передачу сигнала; они также фосфорилируют факторы транскрипции —
переносчики сигнала и активаторы транскрипции [ПСАТ (STAT, от англ.
signal transducers and activators of transcription)]. Эти белки относятся к
семейству БПС, имеющих в структуре SH2домен, узнающий остатки
фосфотирозина. Поэтому они обладают свойством ассоциировать с
фосфорилированным цитокиновым рецептором. Если затем происходит
фосфорилирование молекулы ПСАТ, фактор переходит в активную форму и
образует димер. После транслокации в ядро димер в качестве фактора
транскрипции связывается с промотором инициируемого гена и индуцирует
его транскрипцию.
Некоторые цитокиновые рецепторы могут за счет протеолиза
утрачивать экстрацеллюлярный лигандсвязывающий домен. Домен поступает
в кровь, где конкурирует за связывание с цитокином, что снижает
концентрацию цитокина в крови.
Цитокины и регуляция иммунного ответа
В ответ на вирусную, бактериальную или паразитарную инфекцию
клетки иммунной системы вырабатывают цитокины — сигнальные молекулы,
активирующие различные защитные механизмы. Цитокины представляют
собой полипептиды с сильной и разносторонней активностью, которые служатмедиаторами межклеточных взаимодействий и обладают системным
действием.
Цитокины играют важную роль во многих биологических процессах, в
том числе в инфекциях, воспалении, иммунных реакциях, кроветворении.
Цитокины вырабатываются макрофагами, моноцитами, лимфоцитами,
фибробластами и эндотелиальными клетками. На сегодняшний день вьщеляют
следующие группы цитокинов: интерфероны, интерлейкины (ИЛ), факторы
некроза опухолей (ФНО), факторы роста (например, эпидермальный фактор
роста) и колониестимулирующие факторы.
Цитокины играют важную роль в иммунном ответе, а их избыточная
продукция (например, ИЛ1 при ревматоидном артрите и ФНОа при сепсисе)
может приводить к повреждению тканей и системной патологии. Поэтому
подавление синтеза или активности некоторых цитокинов может изменить
течение воспалительного процесса. Появление очищенных рекомбинантных
цитокинов и их антагонистов дало толчок к многочисленным клиническим
испытаниям в этом направлении.
- фактор роста нервов
Фактор роста нервов (англ. nerve growth factor, NGF) — небольшой
секретируемый белок, поддерживающий жизнеспособность нейронов,
стимулирующий их развитие и активность. Относится к семейству
нейротрофинов. Был идентифицирован первым из факторов роста. Другие
представители этого семейства, которые хорошо известны, включают в себя
нейротрофический фактор головного мозга, нейротрофин3 и нейротрофин
4/5.
NGF является незаменимым для выживания и развития симпатических
и сенсорных нейронов. Без него эти нейроны подвержены апоптозу. Фактор
роста нервов вызывает рост аксонов: исследования показали, что он
способствует их ветвлению и небольшому удлинению. NGF связывается с по
меньшей мере двумя классами рецепторов: LNGFR и TrkA. Оба они связаны с
нейродегенеративными патологиями.Существует доказательство того, что NGF циркулирует по всему телу и
имеет большое значение для поддержания гомеостаза.
Структура NGF была установлена с помощью методов рентгеновской
кристаллографии и опубликована в 1991 году. NGF образует "цистиновый
узел" — структуру, состоящую из бетаслоев, закрученных друг вокруг друга
и связанных тремя дисульфидными связями между остатками цистеина.
NGF предотвращает или уменьшает дегенерации нейронов у животных с
нейродегенеративными заболеваниями. Эти обнадеживающие результаты на
животных привели к ряду клинических испытаний на людях. Экспрессия NGF
увеличивается при воспалительных заболеваниях, при которых он подавляет
воспаление. Кроме того, NGF появляется при процессе восстановления
миелина.
Снижение уровня NGF предполагается у больных шизофренией, однако
данные противоречивы и осложнены действием медикаментов. С целью
разрешения этого противоречия в 2009 году было проведено первое
ещё не получавших
исследование психиатрических пациентов,
нейролептической терапии, в котором было показано, что уровень NGF в
спинномозговой жидкости и плазме крови пациентов понижен по сравнению с
нормой.
Действуют алгогены:
частично прямо – на мембрану ноцицепторов,
частично – непрямо, через васкулярную систему, процессы
микроциркуляции и окружающую ноцицепторы среду.
Множественность алгогенных веществ обеспечивает различия в
возбуждении ноцицепторов и кодирует генез и интенсивность болевого
раздражения.
Так,
выделение серотонина и брадикинина имеет место при
механическом раздражении, простагландина – при химическом.
Таким образом, рецепторы, волокна и проводящие пути центральной
нервной системы, участвующие в механизмах боли, приспособлены
генерировать и передавать информацию в виде паттернов импульсов, а не в
виде модальноспецифических импульсов. Повреждающее раздражениевозбуждает множество проводящих систем, волокна которых конвергируют
и дивергируют по нескольку раз, так что паттерны импульсов могут
подвергаться изменениям на каждом синаптическом уровне. Нервные
импульсы, передаваемые по тонким и толстым волокнам, которые
конвергируют на клетках задних рогов спинного мозга, модулируются
активностью желатинозной субстанции.
Таким же образом конвергенция волокон на клетках ретикулярной
формации позволяет осуществлять значительную суммацию и взаимодействие
входов от расположенных далеко друг от друга областей тела.