1. Общее понятие о катализаторах белковой природы, роль в обмене веществ. Особенности.
2. Свойства ферментов.
3. Строение ферментов. Центры ферментов.
4. Классификация и номенклатура ферментов.
5. Механизм действия, регуляция активности. Практическое применение.
6. Витамины, классификация, биологическая роль.
Ферменты или энзимы – катализаторы белковой природы (от лат. Fermentum закваска, enzyme- в дрожжах). Ферменты были открыты в процессе изучения механизмов брожения.
Й.Я. Берцелиус (шведский химик) выдвинул теорию химического катализа и первым отметил высокую производительность биологических катализаторов.
Л Пастер показал, что сбраживание дрожжами сахаров в спирт катализируется веществами белковой природы - ферментами.
Э. Бухнер доказал, что в водных экстрактах дрожжевых клеток находится набор ферментов, катализирующих превращение сахара в спирт т.е. ферменты отделимы от живых клеток.
В 20 годах ХХ в. Р. Вильштеттер получил ферменты в высокоочищенном состоянии.
В настоящее время энзимология развивается в различных направлениях, получены в чистом виде сотни ферментов, изучена их химическая природа, установлены механизмы действия.
Лекция № 3
Тема. Ферменты: структура, свойства, классификация. Механизм действия, регуляция
активности. Практическое применение. Витамины
ПЛАН
1. Общее понятие о катализаторах белковой природы, роль в обмене веществ. Особенности.
2. Свойства ферментов.
3. Строение ферментов. Центры ферментов.
4. Классификация и номенклатура ферментов.
5. Механизм действия, регуляция активности. Практическое применение.
6. Витамины, классификация, биологическая роль.
Ферменты или энзимы – катализаторы белковой природы (от лат. Fermentum закваска,
enzyme в дрожжах). Ферменты были открыты в процессе изучения механизмов брожения.
Й.Я. Берцелиус (шведский химик) выдвинул теорию химического катализа и первым отметил
высокую производительность биологических катализаторов.
Л Пастер показал, что сбраживание дрожжами сахаров в спирт катализируется веществами
белковой природы ферментами.
Э. Бухнер доказал, что в водных экстрактах дрожжевых клеток находится набор ферментов,
катализирующих превращение сахара в спирт т.е. ферменты отделимы от живых клеток.
В 20 годах ХХ в. Р. Вильштеттер получил ферменты в высокоочищенном состоянии.
В настоящее время энзимология развивается в различных направлениях, получены в чистом
виде сотни ферментов, изучена их химическая природа, установлены механизмы действия.
Функции ферментов сводятся к ускорению превращения определенного вещества (субстрата)
при определенных условиях.
Особенности ферментов. Благодаря уникальной структуре фермента процесс
ферментативного катализа представлен серией элементарных превращений вещества, строжайшим
образом организованным в пространстве и времени. Кооперативность
и жесткая
запрограммированность этапов действия главная особенность ферментов.
В живой клетке множество разнообразных соединений, но реакции между ними образуют
строго определенные метаболические пути, характерные для данной клетки. Индивидуальность
клетки в большой степени определяется уникальным набором ферментов, который генетически
запрограммирован. Отсутствие даже одного фермента или какойнибудь его дефект может
привести к серьезным отклонениям в организме.Ферменты отличаются от других катализаторов уникальными свойствами:
специфичностью действия;
высокой эффективностью действия;
способностью к регуляции;
термолабильностью и неустойчивостью по отношению к кислотам и щелочам;
способностью ускорять реакции при мягких условиях (нормальном давлении, невысокой
температуре и рН).
Специфичность действия или избирательность действия ферментов.
Стереохимическая специфичность: определенный фермент катализирует реакцию с
участием только одной формы субстрата: например лактатдегидрогеназа катализирует
превращение молочной кислоты только L – формы и инертна по отношению к Д – форме кислоты.
Абсолютная специфичность – фермент катализирует только одну реакцию например:
фермент аргиназа расщепляет аргинин на орнитин и мочевину, уреаза расщепляет мочевину на
аммиак и углекислый газ.
Относительная или групповая специфичность – ферменты действуют на группу близких по
строению субстратов, например: гидролиз белков под действием протеаз (расщепление пептидной
связи), но при этом ферменты проявляют дополнительную специфичность – пепсин осуществляет
гидролиз связей, образованных с участием аминогрупп тирозина и фенилаланина, химотрипсин
разрушает пептидные связи тех же аминокислот, образованные карбоксильными группами.
Строение ферментов
Ферменты являются глобулярными белками. Различают простые и сложные ферменты.
Простые ферменты представлены аминокислотными цепями и при гидролизе распадаются только на
аминокислоты. К ним относятся гидролитические ферменты — пепсин, трипсин, папаин, уреаза, лизоцим,
рибонуклеаза, фосфатаза и др.
Большая часть природных ферментов является сложными белками и, помимо полипептидных цепей,
содержит небелковый компонент (кофактор), являющийся абсолютно необходимым для белковой
активности.
Кофакторы имеют различную химическую природу и различаются по прочности связи с
полипептидной цепью.
Полипептидная часть фермента называется апоферментом,
Небелковая, легко диссоциирущая с белковой частью – коферментом,
прочно связанная с белком простетической группой (кофактор), молекула в целом –холофермент.
Коферменты и кофакторы активно участвуют и химических реакциях, играя роль переносчиковпротонов, электронов или различных функциональных групп. Эти группы определяют специфичность
действия ферментов, участвуют в связывании фермента с субстратом , стабилизируют белковую часть
фермента.
Коферменты и кофакторы связаны с апоферментом водородными, ионными гидрофобными
связями.
Кофактором называется любой фактор, абсолютно необходимый для выполнения белком его
биологической роли. Им могут быть любые небелковые органические молекулы или ионы металлов.
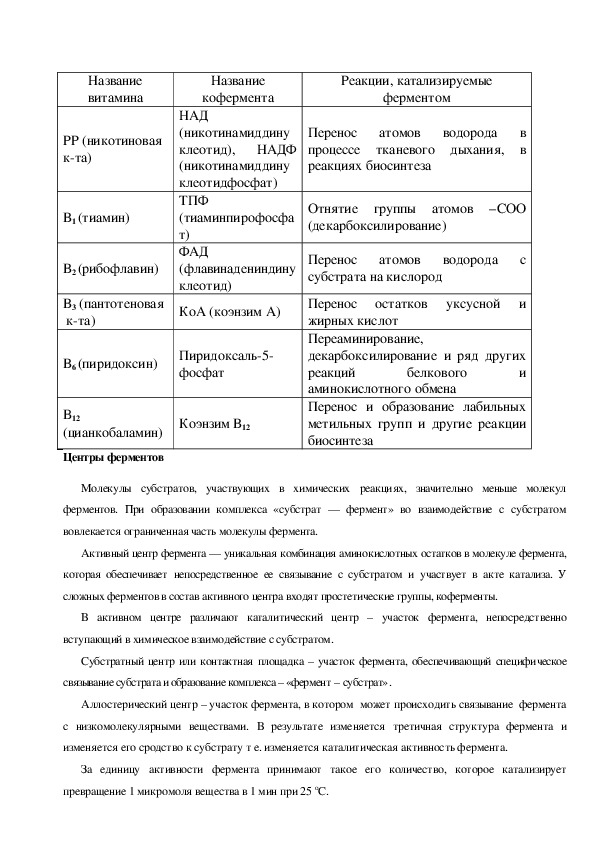
Коферменты представлены водорастворимыми витаминами ( группы В, витамин Н, РР),
нуклеотидные производные, липоевая кислота, фосфорные эфиры некоторых полисахаридов.Название
витамина
РР (никотиновая
кта)
В1 (тиамин)
В2 (рибофлавин)
В3 (пантотеновая
кта)
Название
кофермента
НАД
(никотинамиддину
клеотид),
НАДФ
(никотинамиддину
клеотидфосфат)
ТПФ
(тиаминпирофосфа
т)
ФАД
(флавинадениндину
клеотид)
КоА (коэнзим А)
В6 (пиридоксин)
Пиридоксаль5
фосфат
B12
(цианкобаламин)
Центры ферментов
Коэнзим В12
Реакции, катализируемые
ферментом
Перенос атомов водорода в
процессе тканевого дыхания, в
реакциях биосинтеза
Отнятие группы атомов –СОО
(декарбоксилирование)
Перенос атомов водорода с
субстрата на кислород
Перенос остатков уксусной и
жирных кислот
Переаминирование,
декарбоксилирование и ряд других
реакций
и
аминокислотного обмена
Перенос и образование лабильных
метильных групп и другие реакции
биосинтеза
белкового
Молекулы субстратов, участвующих в химических реакциях, значительно меньше молекул
ферментов. При образовании комплекса «субстрат — фермент» во взаимодействие с субстратом
вовлекается ограниченная часть молекулы фермента.
Активный центр фермента — уникальная комбинация аминокислотных остатков в молекуле фермента,
которая обеспечивает непосредственное ее связывание с субстратом и участвует в акте катализа. У
сложных ферментов в состав активного центра входят простетические группы, коферменты.
В активном центре различают каталитический центр – участок фермента, непосредственно
вступающий в химическое взаимодействие с субстратом.
Субстратный центр или контактная площадка – участок фермента, обеспечивающий специфическое
связывание субстрата и образование комплекса – «фермент – субстрат».
Аллостерический центр – участок фермента, в котором может происходить связывание фермента
с низкомолекулярными веществами. В результате изменяется третичная структура фермента и
изменяется его сродство к субстрату т е. изменяется каталитическая активность фермента.
За единицу активности фермента принимают такое его количество, которое катализирует
превращение 1 микромоля вещества в 1 мин при 25 оС.Формирование активного центра фермента начинается на ранних этапах его синтеза, когда линейная
полипептидная цепь превращается в трехмерное тело с определенной конфигурацией. Белок со
сформированной третичной структурой приобретает функциональную (в частности, каталитическую)
информацию. Любые воздействия, приводящие к денатурации фермента (нарушению его третичной
структуры), нарушают структуру активного центра и ведут к потере ферментом каталитических свойств.
При восстановлении нативной трехмерной структуры белкафермента (ренатурации) восстанавливается и
его каталитическая активность.
Важнейшие аминокислотные остатки в активном центре
лизоцима
У некоторых ферментов в активном центре располагается и кофактор. Одни R
группы активного центра принимают участие в связывании субстрата, другие в катализе.
Мультимолекулярные (надмолекулярные) ферментные системы — комплексы, состоящие из
различных по строению ферментов, катализирующих последовательные ступени превращения
определенного субстрата. Их особенностями является прочность ассоциации ферментов и определеннаяпоследовательность прохождения промежуточных стадий во времени, определяемая порядком расположения
ферментов в пространстве.
Молекулярная масса таких комплексов колеблется от 2,3 X 106 до10 X 10б. Образование
подобных систем резко сокращает расстояния, на которые перемещаются молекулы субстратов.
Мультимолекулярные ферментные системы могут быть структурно связанными с какойлибо органеллой
(рибосомой, митохондрией) или с биомембраной, такие системы обеспечивают жизненно важные для
клетки функции (например, тканевое дыхание, окисление жирных кислот, ЦТК). К мультиферментным
системам относятся пируватдегидрогеназный и аетоглутаратдегидрогеназный комплексы..
Классификация и номенклатура ферментов.
В основу принятой классификации ферментов положен принцип типа катализируемой реакции.
Согласно Международной классификации ферменты делят на шесть главных классов.
1.Оксидоредуктазы — ферменты, катализирующие окислительновосстановительные реакции.
Систематические названия составляются по схеме: «донор: акцептор оксидоредуктаза» (например,
лактат: НАД+ оксидоредуктаза). Среди оксидоредуктаз различают аэробные дегидрогеназы, или
оксидазы (катализируют перенос протонов или электронов на кислород), анаэробные дегидрогеназы
(катализируют перенос протонов или электронов на промежуточный субстрат), цитохромы
(катализируют перенос только электронов). К оксидоредуктазам относятся гемсодержащие
ферменты: каталаза и пероксидаза.
2.Трансферазы — ферменты, катализирующие реакции межмолекулярного переноса атомов, групп атомов
и радикалов. Систематические названия составляются по схеме: «донор: транспортируемая группа —
трансфераза». Среди трансфераз выделяют катализирующие перенос одноуглеродных остатков,
ацильных, гликозильных, альдегидных или кетонных, нуклеотидных остатков, азотистых групп,
остатков кислот.
3.Гидролазы — ферменты, катализирующие расщепление внутримолекулярных связей при участии
молекул воды. Систематические названия составляются по схеме: «субстрат — гидролаза». К
гидролазам относятся
эстеразы (катализируют реакции гидролиза и синтеза сложных эфиров),
гликозидазы (катализируют разрыв гликозидных связей),
фосфатазы,
4.Лиазы — ферменты, катализирующие разрыв связей С—О,
пептидазы и амидазы (катализируют разрыв фосфоангидридных, пептидных и амидных связей).
С—С, С—N, реакции
негидролитического отщепления различных групп от субстратов. Реакции сопровождаются
образованием двойной связи или присоединением к месту разрыва двойной связи. Систематические
названия составляются по схеме: «субстрат — лиаза».
5.Изомеразы — ферменты, катализирующие геометрическую и оптическую изомеризацию.Систематические названия составляются по схеме: «субстратцистрансизомераза». При внутри
молекулярном переносе группы фермент называется «мутаза».
6.Лигазы (синтетазы) — ферменты, катализирующие синтез органических веществ из двух исходных
молекул с использованием энергии макроэрга (АТФ или др.). Систематические названия
составляются по схеме: «X : Y лигаза», где X и Y — исходные вещества.
Список ферментов содержит для каждого фермента кодовый номер (шифр), систематическое
(рациональное) название, рекомендуемое (рабочее) название, химическую реакцию, катализируемую им,
и примечания о специфичности действия.
Код каждого фермента состоит из четырех цифр, разделенных точками: номер главного класса
фермента, номер подкласса, номер подгруппы в подклассе и порядковый номер фермента
В живой клетке множество разнообразных соединений, но реакции между ними образуют
строго определенные метаболические пути, характерные для данной клетки. Индивидуальность
клетки в большой степени определяется уникальным набором ферментов, который генетически
запрограммирован. Отсутствие даже одного фермента или какойнибудь его дефект может
привести к серьезным отклонениям в организме.
Механизмы действия ферментов
На первоначальном этапе происходит специфическое связывание фермента с лигандом
субстратом (S). Это происходит в области активного центра, который формируется из нескольких
специфических радикалов аминокислот, определенным образом ориентированных в пространстве.
Рассмотрим строение фермента лизоцима.
В процессе ферментативной реакции различают 4 этапа:
1 – присоединение молекулы субстрат к ферменту и образование ферментсубстратного
комплекса;
2 – изменение субстрата под действием фермента, делающее его доступным для химической
реакции, т.е. активизация субстрата;
3 – химическая реакция;
4 – отделение продуктов реакции от фермента.
Это можно записать в виде схемы:
1 2 3 4
E + S ES ES* EP E + P
Начальная скорость (Vo). При обычных условиях, когда [S] >> [E], начальная скорость
прямо пропорциональна концентрации фермента.Максимальная скорость (Vmax). При фиксированной концентрации фермента, скорость
реакции стремится к конечному максимальному значению, в то время как концентрация субстрата
растет. Насыщение фермента субстратом наступает, когда весь фермент включен в фермент
субстратный комплекс.
Константа Михаэлиса (Km). В случае, когда все активные центры заняты, и свободные
молекулы фермента отсутствуют, Vo=Vmax. При таком условии говорят о 100% насыщении. При
50% насыщении, когда Vo=1/2 Vmax из уравнения Михаэлиса Ментен следует:
Vmax / 2 = Vmax [S] / Km + [S],
или в преобразованном виде: Km + [S] = 2 [S]; Km = [S].
Следовательно, Km имеет размерность концентрации. Таким образом, Кm – это такая
концентрация субстрата, которая необходима для связывания половины имеющегося
фермента и достижения половины максимальной скорости.
Из этого определения следует, что Km можно использовать для оценки сродства
фермента по отношению к данному субстрату. Оценить субстратную специфичность можно по
такому правилу: чем ниже значение Km, тем лучше (предпочтительнее) субстрат для данного
фермента.
Витамины, роль в обмене веществ.
Витамины это низкомолекулярные органические вещества разнообразного строения.
Объединены в одну группу по следующим признакам.
1. Витамины абсолютно необходимы организму и в очень небольших количествах.
2. Витамины не синтезируются в организме и должны поступать извне или синтезироваться
микрофлорой кишечника.
Витамины играют одинаковую роль во всех формах жизни, но высшие животные утратили
способность к их синтезу. Например, аскорбиновая кислота (витамин "С") не синтезируется в
организмах человека, обезьян и морской свинки, так как в процессе эволюции была утеряна
ферментная система синтеза этого витамина из глюкозы.
2. Классификация витаминов
1. Водорастворимые витамины. К этой группе относят витамины С, Р, В1, В2, В3, ВC, В6, В12,
РР, Н.
2. Жирорастворимые витамины: А, Д, Е, К.
Большинство водорастворимых витаминов должно поступать регулярно с пищей, так как
они быстро выводятся или разрушаются в организме.
Жирорастворимые витамины могут депонироваться в организме. Кроме того, они плохо
выводятся, поэтому иногда при избытке жирорастворимых витаминов наблюдаются
гипервитаминозы заболевания, связанные с интоксикацией организма высокими дозами
жирорастворимых витаминов. Такие заболевания описаны для витаминов А и Д.
Для большинства витаминов известно, что их производные входят в состав коферментов и
простетических групп ферментов.
Витамин С,Е известны своей антиоксидантной активностью и участием в многообразных
биохимических процессах.
Авитаминоз это заболевание, которое развивается при полном отсутствии того или иного
витамина в организме. В настоящее время авитаминозы обычно не встречаются, а бывают
гиповитаминозы при недостатке витамина в организме.
Причины развития гипо и авитаминозовВсе причины можно разделить на внешние и внутренние.
Внешние причины гиповитаминозов
1. Недостаточное содержание витамина в пище (при неправильной обработке пищи, при
неправильном хранении пищевых продуктов).
2. Состав рациона питания (например, отсутствие в рационе овощей и фруктов).
3. Не учитывается потребность в том или ином витамине. Например, при белковой диете
возрастает потребность в витамине "РР" (при обычном питании он может частично
синтезироваться из триптофана). Если человек потребляет много белковой пищи, то может
увеличиться потребность в витамине "В6" и снизиться потребность в витамине РР.
1. Социальные причины: урбанизация населения, питание исключительно высокоочищенной и
консервированной пищей; наличие антивитаминов в пище. Социальные причины развития
авитаминозов существуют в мире. Например, в отдаленных районах Севера, в рационе
людей мало овощей и фруктов.
В крупных городах люди недостаточно обеспечены солнечным светом поэтому может быть
гиповитаминоз Д.
Внутренние причины гиповитаминозов
1. Физиологическая повышенная потребность в витаминах, например, в период
беременности, при тяжелом физическом труде.
2. Длительные тяжелые инфекционные заболевания, а также период выздоровления.
3. Нарушение всасывания витаминов при некоторых заболеваниях ЖКТ, например, при
желчнокаменной болезни нарушается всасывание жирорастворимых витаминов.
4. Дисбактериоз кишечника. Имеет значение, так как некоторые витамины синтезируются
полностью микрофлорой кишечника (это витамины В3, Вc, В6, Н, В12 и К).
2. Генетические дефекты некоторых ферментативных систем. Например, витамин Д
резистентный рахит развивается у детей при недостатке ферментов, участвующих в
образовании активной формы витамина Д (1,25диоксихолекальциферола).
Витамин А (ретинол, антиксерофтальмический)
Наиболее ранний и специфический признак гиповитаминоза А гемералопия (куриная
слепота) нарушение сумеречного зрения. Возникает изза недостатка зрительного пигмента
родопсина. Родопсин содержит в качестве активной группы ретиналь (альдегид витамина А)
находится в палочках сетчатки. Эти клетки (палочки) воспринимают световые сигналы низкой
интенсивности.
Родопсин = опсин (белок) + цисретиналь.
При возбуждении родопсина светом, цисретиналь, в результате ферментативных
перестроек внутри молекулы переходит в полностьютрансретиналь (на свету). Это приводит к
конформационной перестройке всей молекулы родопсина. Родопсин диссоциирует на опсин и
трансретиналь, что является пусковым механизмом, возбуждающим в окончаниях зрительного
нерва импульс, который затем передается в мозг.
В темноте, в результате ферментативных реакций трансретиналь вновь превращается в
цисретиналь и, соединяясь с опсином, образует родопсин.Витамин А также влияет на процессы роста и развития покровного эпителия. Поэтому при
авитаминозе наблюдается поражение кожи, слизистых оболочек и глаз, которое проявляется в
патологическом ороговении кожи и слизистых. У больных развивается ксерофтальмия сухость
роговой оболочки глаза, так как происходит закупорка слезного канала в результате ороговения
эпителия. Так как глаз перестает омываться слезой, которая обладает бактерицидным действием,
развиваются конъюнктивиты, изъязвление и размягчение роговицы кератомаляция.
При авитаминозе А может быть также поражение слизистой ЖКТ, дыхательных и
мочеполовых путей. Нарушается устойчивость всех тканей к инфекциям. При развитии
авитаминоза в детстве задержка роста.
В настоящее время показано участие витамина А в защите мембран клеток от окислителей
т. е. витамин А обладает антиоксидантной функцией.
Витамин А запасается в печени.
Пищевые источники печень морских рыб и млекопитающих, желток яиц, цельное молоко,
рыбий жир. Овощи и фрукты краснооранжевого цвета (томаты, морковь и др.) содержат много
каротина водорастворимого предшественника витамина А, имеющего в молекуле 2 иононовых
кольца.
Высокие дозы витамина А могут приводить к токсическим эффектам. Характерные
проявления гипервитаминоза воспаление глаз, гиперкератоз, выпадение волос.
Суточная потребность в витамине А 12,5 мг, в каротине в 2 раза больше.
Витамин К (филлохинон)
Витамин К необходим для
нормального синтеза протромбина (фактор
II) предшественника одного из белков
системы свертывания тромбина. Тромбин
это фермент,
который катализирует
реакцию превращения фибриногена в
фибрин основу кровяного сгустка при
активации системы свертывания крови.
При недостатке витамина К синтезируется дефектная молекула протромбина и ряда
других факторов свертывания крови. Причина нарушение ферментативного карбоксилирования
глутаминовой кислоты, необходимой для связывания Са2+ белками системы свертывания.
Основное проявление недостаточности нарушение свертывания крови, в результате которого
происходят самопроизвольные паренхиматозные и капиллярные кровотечения.
Авитаминоз, как правило, связан с нарушением выделения желчи в ЖКТ (при
желчнокаменной болезни).
Пищевые источники ягоды рябины, капуста, арахисовое масло и др. растительные масла.
Витамин К также синтезируется микрофлорой кишечника, поэтому одна из причин
гиповитаминозов при недостатке витамина в пище дизбактериоз кишечника.
Если больной страдает гиповитаминозом К, например, при некоторых видах желтух, то
операции даже удаление зуба могут сопровождаться длительным кровотечением.
Синтезирован водорастворимый аналог витамина К викасол, который используют при
лечении гиповитаминозов, связанных с нарушением всасывания витамина К из кишечника.
Известны природные антивитамины К например, дикумарин, салициловая кислота,
которые применяют при лечении тромбозов, так как антивитамины К способны снижать
количество протромбина в крови.
Суточная потребность точно не установлена, так как витамин синтезируется микрофлорой.
Считают, что в сутки потребность около 1 мг.
Витамин Е.Является антиоксидантом. При недостаточности витамина Е дегенеративные изменения в
печени, нарушение функций биологических мембран. Витамин Е предохраняет липиды клеточных
мембран от окисления активными формами кислорода.
Авитаминоз проявляется при очень длительном голодании или при стойком нарушении
желчевыделительной функции печени (нарушение всасывания жиров). При этом наблюдаются
шелушение кожи, мышечная слабость, стерильность нарушением функции размножения.
Поскольку витамин Е широко распространен в природе (растительные масла, семена пшеницы и
др. злаков, сливочное масло), то авитаминоз встречается редко.
Суточная потребность около 1030 мг.
Витамин Д (холекальциферол, антирахитный)
Сам витамин Д не обладает витаминной активностью, но он служит предшественником
1,25дигидроксихолекальциферола (1,25дигидроксивитамина Д3).
Синтез активной формы протекает в два этапа в печени присоединяется оксигруппа в
положении 25, а затем в почках оксигруппа в положении 1. Из почек активный витамин Д3
переносится в другие органы и ткани главным образом в тонкий кишечник и в кости, где витамин
Д участвует в регуляции обмена Са и Р.
Недостаток витамина Д приводит к развитию нарушений фосфорнокальциевого обмена и
процессов окостенения. В результате у детей развивается рахит, связанный с недостатком Са и Р.
Характерные признаки рахита остеомаляция ("размягчение" костей запаздывание окостенения),
запаздывание закрытия родничков, деформации грудной клетки, позвоночника, конечностей. У
таких детей снижен мышечный тонус, наблюдается раздражительность, потливость, выпадение
волос.
У взрослых при недостатке витамина Д наблюдается остеопороз разрежение костной
ткани в результате вымывания солей кальция из скелета.
Потребность в витамине Д повышается у беременных.
При благоприятных условиях витамин Д может синтезироваться в организме человека из
предшественника 7дегидрохолестерина под действием ультрафиолетовых лучей
(фотохимическая реакция) в результате разрыва связи в кольце В.
Пищевые источники рыба, рыбий жир, печень, сливочное масло, желток яиц.
Суточная доза витамина Д3 1020 мкг. Высокие дозы витамина Д (выше 1,5 мг в сутки)
крайне токсичны. При гипервитаминозе кроме интоксикации наблюдается отложение
гидроксиапатита в некоторых внутренних органах (кальцификация почек, кровеносных сосудов).Витамин С (аскорбиновая кислота, антицинготный, антискорбутный)
В 1932 г. впервые выделен из
сока лимона, через два года искусственно синтезирован. Важное свойство способность
аскорбиновой кислоты легко окисляться.
Биологическая роль витамина С (связана с его участием в окислительновосстановительных
реакциях).
1. Витамин С, являясь сильным восстановителем, играет роль кофактора в реакциях
окислительного гидроксилирования, что необходимо для окисления аминокислот пролина и
лизина в оксипролин и в оксилизин в процессе биосинтеза коллагена. Коллаген может
синтезироваться и без участия витамина С, но такой коллаген не является полноценным (не
формирутся его нормальная структура). Поэтому при недостатке витамина С ткани, содержащие
много коллагена, становятся непрочными, ломкими. В первую очередь нарушается структура
стенок сосудов, повышается их проницаемость, наблюдаются кровоизлияния под кожу и под
слизистые оболочки.
2. Участвует в синтезе стероидных гормонов надпочечников.
3. Необходим для всасывания железа.
4. Участвует в неспецифической иммунной защите организма.
Авитаминоз "С" цинга. Проявления цинги: болезненность, рыхлость и кровоточивость
десен, расшатывание зубов, нарушение целостности капилляров подкожные кровоизлияния,
отечность и болезненность суставов, нарушение заживления ран, анемия. Иногда цинга
развивается у новорожденных на искусственном вскармливании пастеризованным молоком, в
которое не добавлен витамин С.
В основе всех изменений при цинге, за исключением анемии, лежит нарушение синтеза
коллагена. Анемия связана с нарушением всасывания железа.
В настоящее время цинга не распространена, но весной у многих людей наблюдается
недостаток (гиповитаминоз) витамина "С", что проявляется, например, повышенной
утомляемостью, понижением иммунитета.
Основные источники витамина С: свежие зеленые овощи и фрукты.
Следует помнить, что витамин С легко разрушается при нагревании, особенно в щелочной
среде в присутствии кислорода, ионов железа и меди. Хорошо сохраняется в кислой среде (в
квашеной капусте, в клюкве, в ягодах черной смородины и плодах шиповника). При длительном
хранении овощей и фруктов содержание в них витамина "С" уменьшается.
Источником витамина С является также хвоя ели и сосны.
Суточная потребность около 100 мг в сутки.
Витамин Р (рутин, витамин проницаемости)
Биологическая роль стабилизация основного вещества соединительной ткани, путем
ингибирования фермента гиалуронидазы.При недостатке витамина Р у людей повышается проницаемость кровеносных сосудов,
которое сопровождается кровоизлияниями и кровотечениями. Витамин Р усиливает действие
витамина С (снижает потребность в нем)
Пищевые источники: зеленые овощи и фрукты, кожура лимона.
Суточная потребность не установлена.
Витамины группы В
Витамин B1 (тиамин, антиневритный).
Производное витамин В1 ТДФ (ТПФ) является коферментом пируватдегидрогеназного
комплекса (фермента пируваткарбоксилазы),
кетоглутаратдегидрогеназного комплекса и
фермента транскетолазы, а также входит в состав кофермента транскетолаз ферментов
неокислительного этапа ГМФпути..
При недостаточности витамина В1 может возникнуть болезнь берибери, характерная для
тех стран Востока, где основным продуктом питания служит очищенный рис и кукуруза.
Для этого заболевания характерна мышечная слабость, нарушение моторики кишечника,
потеря аппетита, истощение, периферический неврит (характерный признак человеку больно
вставать на стопу больные ходят "на цыпочках"), спутанность сознания, нарушения работы
сердечнососудистой системы. При "берибери" повышается содержание пирувата в крови.
Пищевые источники витамина В1 ржаной хлеб. В кукурузе, рисе, пшеничном хлебе
витамин В1 практически отсутствует. Это объясняется тем, что в зерне ржи тиамин распределен
по всему зерну, а в других злаках он содержится только в оболочке зерен.
Суточная потребность 1.5 мг/сутки.
Витамин В2
(рибофлавин) входит в состав флавинмононуклеотида (ФМН) и
флавинадениндинуклеотида (ФАД) простетических групп флавиновых ферментов.
Его биологическая функция в организме участие в окислительновосстановительных
реакциях в составе флавопротеидов (ФП).
Недостаточность этого витамина часто встречается в России. Особенно часто бывает у
людей, которые не употребляют в пищу черный ржаной хлеб. Проявление гиповитаминоза:
ангулярные дерматиты в углах рта ("заеда"), глаз. Часто это сопровождается кератитами
(воспаление роговицы). В очень тяжелых случаях бывает анемия. Очень часто сочетаются
сочетанные гиповитаминозы витаминов "В2" и "РР",так как эти витамины содержатся в одних и
тех же продуктах.Пищевые источники: ржаной хлеб, молоко, печень, яйца, овощи желтого цвета, дрожжи.
Суточная потребность: 24 мг/сутки.
Фолиевая кислота (ВC).
В составе 3 структурных единицы: птеридин, ПАБК (парааминобензойная кислота) и
глутаминовая кислота.
Часто ПАБК (парааминобензойную кислоту) тоже называют витамином. Но это неверно.
ПАБК это фактор роста для микроорганизмов, которые синтезируют фолиевую кислоту.
Активный С1 извлекается из глицина или серина с помощью фермента, в небелковой части
фолиевая кислота. Фолиевая кислота дважды
которого содержится витамин Вc
восстанавливается в организме (к ней присоединяется водород).
ТГФК является коферментом ферментов, переносящих одноуглеродные радикалы.
Из метиленТГФК могут образовываться все другие формы активного С1: формилТГФК,
метилТГФК, метенТГФК, оксиметилТГФК в результате реакций окисления или восстановления
метиленТГФК.
Фолиевая кислота в виде тетрагидрофолиевой кислоты является коферментом,
участвующим в ферментативных реакциях, связанных с переносом активных одноуглеродных
радикалов. Например: биосинтез пуриновых и пиримидиновых мононуклеотидов.
При авитаминозе у человека наблюдается макроцитарная анемия, при которой нарушен
синтез ДНК в клетках красного костного мозга, для больных характерна потеря веса.
Пищевые источники: зеленые листья овощей, дрожжи, мясо, шпинат.
Авитаминозы встречаются редко, так как потребность в этом витамине компенсируется за
счет микрофлоры кишечника. При некоторых заболеваниях кишечника, когда возникают
дисбактериозы, нарушается всасывание фолиевой кислоты.
Суточная потребность: 0.20.4 мг.
Витамин В6 (пиридоксин) в форме пиридоксальфосфата является простетической группой
трансаминаз и декарбоксилаз аминокислот.
При авитаминозе В6 наблюдаются нарушения обмена аминокислот.
В6 участвует в реакциях синтеза гема гемоглобина (синтез dаминолевулиновой кислоты).
Поэтому при недостатке В6 у человека развивается анемия, наблюдаются дерматиты.
Недостаток В6 может развиться у больных туберкулезом, потому что этих больных лечат
препаратами, синтезированными на основе изониазида это антагонисты витамина В6.
Пищевые источники: ржаной хлеб, горох, картофель, мясо, печень, почки.
Суточная потребность взрослого человека: 0,150,20 мг.
Пантотеновая кислота (витамин В3) Молекула пантотеновой кислоты состоит из бета
аланина и 2,4дигидроксидиметилмасляной кислоты. Важность этого витамина в том, что он
входит в состав HSKoA (кофермента ацилирования).Строение КоА:
а) тиоэтиламин;
б) пантотеновая кислота;
в) 3фосфоаденозин5дифосфат.
HSКоА кофермент ацилирования, т. е. входит в состав ферментов, которые катализируют
перенос ацильных остатков. Поэтому В3 участвует в бетаокислении жирных кислот,
окислительном декарбоксилировании
кетокислот, биосинтезе нейтрального жира, липоидов,
стероидов, гема, ацетилхолина.
При недостатке пантотеновой кислоты при дисбактериозе у человека развиваются
дерматиты, в тяжелых случаях изменения со стороны желез внутренней секреции, в том числе
надпочечников. Также наблюдается депигментация волос, истощение.
Пищевые источники: яичный желток, печень, дрожжи, мясо, молоко.
Суточная потребность: 10мг/сут.
Витамин В12 (кобаламин) (антианемический витамин).
Имеет сложное строение, структура молекулы похожа на гем, но вместо железа кобальт.
В состав В12 входит также нуклеотидная структура, похожая на АМФ.
Производное витамина В12 является коферментом. Этот витамин необходим для синтеза
нуклеиновых кислот. Он обеспечивает переход оксирибонуклеотидов в дезоксирибонуклеотиды
(РНК в ДНК).
Недостаток этого витамина может привести к развитию злокачественной тромбоцитарной
анемии, нарушениям функции центральной нервной системы.
Как правило, встречается сочетанный недостаток витамина В12 и фолиевой кислоты.
Анемия развивается не потому, что В12 мало поступает с пищей, а при отсутствии особого
гликопротеина, который называется "внутренний фактор Кастла" и вырабатывается в желудке.
Фактор Кастла необходим для всасывания витамина В12. При удалении части желудка, гастритах
уменьшается выработка фактора Кастла.
Это единственный витамин, который синтезируется только микрофлорой кишечника.
Это единственный водорастворимый витамин, который депонируется в организме (в
печени).
Суточная потребность: 2,55 мкг.
Витамин РР (антипеллагрический)
Химическое название: никотинамид.
Входит в состав НАД и НАДФ, т. е. входит в состав коферментов никотинамидных
дегидрогеназ. Его роль участие в окислительновосстановительных реакциях. При недостатке РР
развивается пеллагра. При пеллагре наблюдаются три "Д":
1) дерматит;2) диарея;
3) деменция (поражение центральной нервной системы).
Источники РР: мясо, бобовые, орехи, рыба и вообще продукты, богатые белком.
Витамин РР может частично синтезироваться из триптофана.
Если человек съедает много белковой пищи, то потребность в этом витамине снижается. Из
60 г белка может синтезироваться 1 мг витамина РР.
Суточная потребность: 1525 мг/сутки.
Витамин Н (биотин)
В составе молекулы биотина имеются имидазоловое и тиоэфирное кольца, к ним
присоединен радикал валериановая кислота.
Витамин Н входит в состав ферментов карбоксилаз: АцетилКоАкарбоксилазы,
пируваткарбоксилазы и других.
Всасыванию биотина в кишечнике препятствует овидин белок, содержащийся в сырых
яйцах. При термической обработке яиц происходит днатурация овидина.
При авитаминозе наблюдаются дерматиты, поражения ногтей, анемия. Синтезируется
микрофлорой кишечника.