Поделиться

Тема. Липиды: классификация, структура, функции. Роль в построении мембран. Обмен липидов.
План:
1. Липиды, общая характеристика и классификация липидов.
2. Простые липиды, жиры, воски, стериды. Сложные липиды: фосфолипиды, гликолипиды.
3. Биологические функции липидов. Роль липидов в построении мембран.
4. Обмен липидов.
5. Энергетический эффект распада жиров.
Различают следующие основные классы липидов:
Простые липиды: сложные эфиры жирных кислот с различными спиртами:
- Глицериды (ацилглицерины, или ацилглицеролы - по международной номенклатуре) представляют собой сложные эфиры трехатомного спирта глицерина и высших жирных кислот.
- Воска: сложные эфиры высших жирных кислот и одноатомных или двухатомных спиртов.
Стериды – сложные эфиры высших жирных кислот и спирта стерола
Сложные липиды: сложные эфиры жирных кислот со спиртами, дополнительно содержащие и другие группы.
- Фосфолипиды: липиды, содержащие, помимо жирных кислот и спирта, остаток фосфорной кислоты. В их состав входят азотистые основания и другие компоненты: глицерофосфолипиды (спирт - глицерол); сфинголипиды (спирт - сфингозин).
- Гликолипиды (гликосфинголипиды).
- Стероиды.
- Липопротеины
Воска - сложные эфиры высших жирных кислот и высших одноатомных или двухатомных спиртов с числом углеродных атомов от 16 до 22. Общие формулы можно представить так:
Воска входят в состав жира, покрывающего кожу, шерсть, перья. У растений 80% от всех липидов, образующих пленку на поверхности листьев и плодов, составляют воска. Воска являются нормальными метаболитами некоторых микроорганизмов. Природные воска (например, пчелиный воск, спермацет, ланолин) обычно содержат, кроме указанных сложных эфиров, некоторое количество свободных жирных кислот, спиртов и углеводородов с числом углеродных атомов 21-35.
Фосфолипиды представляют собой сложные эфиры многоатомных спиртов: глицерина или сфингозина с высшими жирными кислотами и фосфорной кислотой. В состав фосфолипидов входят также азотсодержащие соединения: холин, этаноламин или серин.
Фосфолипиды (глицерофосфолипиды). Глицерофосфолипиды являются производными фосфатидной кислоты. В их состав входят глицерин, жирные кислоты, фосфорная кислота и азотсодержащие соединения. Общая формула глицерофосфолипидов:
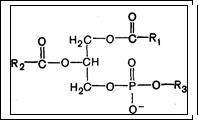
В этих формулах R1 и R2- радикалы высших жирных кислот, а R3- чаще радикал азотистого соединения - этаноламин (НО — СН2 — СН2 — N +Н3), холин [НО — СН2 — СН2 — Na+(СН3)3]. -
. Для всех глицерофосфолипидов характерно, что одна часть их молекул (радикалы R1 и R2-) обнаруживает резко сраженную гидрофобность, тогда как другая часть гидрофильна благодаря отрицательному заряду фосфорной кислоты и положительному заряду радикала R3.
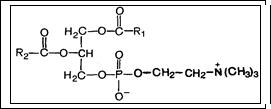
Фосфатидилхолины (лецитины). В состав молекулы входят глицерин, высшие жирные кислоты, фосфорная кислота и холин:
Биологические функции липидов:
Энергетическая - эту функцию выполняют жирные кислоты, освобождающиеся после распада жиров.
Структурную - фосфолипиды, гликолипиды и холестерин участвуют в образовании клеточных мембран.
Холестерин является структурным компонентом мембран, а также предшественником желчных кислот и стероидных гормонов.
Регуляторная функция - производные некоторых полиненасыщенных жирных кислот биологически активные вещества - (простагландины), Поэтому эти жирные кислоты представляют собой незаменимые пищевые факторы.
Транспортную - вместе с жирами при всасывании в организм поступают жирорастворимые витамины (А, Е, D, К).
Липиды плохо растворимы в воде (гидрофобность)
Некоторые (гликолипиды, фосфолипиды, желчные кислоты) амфифильны, так как имеют в своем составе гидрофильные и гидрофобные части.
Расщепление липидов в желудочно-кишечном тракте. Расщепление липидов происходит в 12-перстной кишке, куда поступают липаза с соком поджелудочной железы и конъюгированные желчные кислоты в составе желчи.
Эмульгирование жира - обязательное условие для переваривания, так как делает гидрофобный субстрат более доступным для действия гидролитических ферментов - липаз.
Эмульгирование происходит при участии желчных кислот , которые из-за своей амфифильности, окружают каплю жира и снижают поверхностное натяжение, что приводит к дроблению капли.
Гидролиз жира осуществляется при участии панкреатической липазы , которая, сорбируясь на поверхности капель жира, расщепляет эфирные связи в триацилглицеринах (ТАГ).
Всасывание происходит при участии желчных кислот, которые образуют вместе с моноацилглицеринами, холестерином и жирными кислотами смешанные мицеллы - растворимые комплексы, обеспечивающие переход продуктов гидролиза в клетки слизистой кишечника.
Ресинтез триацилглицеринов из продуктов расщепления происходит в клетках слизистой кишечника.
Транспорт ресинтезированного жира через лимфатическую систему и кровоток возможен только после включения его в состав липопротеинов.
В кишечнике образуются два типа липопротеинов: хиломикроны - ХМ и в небольшом количестве липопротеины очень низкой плотности - ЛОНП. В составе хиломикронов экзогенные жиры доставляются в органы и ткани.
Потребность в жирах составляет 50-100 г в сутки - в зависимости от характера питания и энергетических затрат.
Депонирование и мобилизация жиров
Жиры, как и гликоген, являются формами депонирования энергетического материала. Жиры более эффективные источники энергии . При голодании запасы жира у человека истощаются за 5-7 недель, тогда как гликоген полностью расходуется примерно за сутки. Если поступление жира превышает потребности организма в энергии, то жир депонируется в адипоцитах - специализированных клетках жировой ткани.
1. Поступают из хиломикронов, которые приносят экзогенные жиры из кишечника
2. Поступают из ЛОНП, которые транспортируют эндогенные жиры, синтезированные в печени из глюкозы
3. Образуются из глюкозы в самих клетках жировой ткани.
Под действием липазы жиры гидролизуются с образованием глицерина и высших жирных кислот.
Глицерин, не зависимо от того, поступит ли он на ресинтез жиров или будет распадаться далее, подвергается фосфорилированию
Глицерокиназа глицерофосфатдегидрогеназа
![]()
![]()
![]()
![]() Глицерин + АТФ
глицерофосфат диоксиацетонфосфат
Глицерин + АТФ
глицерофосфат диоксиацетонфосфат
НАД НАДН
Диоксиацетонфосфат изомеризуется в 3-фосфоглицериновый альдегид и вступает в реакции обмена.
Жирные кислоты транспортируются кровью в виде комплексов с сывороточными альбуминами в разные органы и ткани, где включаются в процесс окисления.
Окисление жирных кислот. Окисление жирных кислот происходит путем β -окисления
Ненасыщенные жирные кислоты предварительно восстанавливаются до предельных.
Процесс распада осуществляется ступенчато, путем отщепления от молекулы кислоты двухуглеродных фрагментов в виде ацетил - КоА.
Жирная кислота поступает из кровотока в цитозоль и активируется путем конденсации с коферментом А (КоА), образуется тиоэфир.
β -окисление происходит в митохондриях клетки.
Для того чтобы пройти через митохондриальную мембрану, жирной кислоте необходим специальный переносчик - карнитин . Поэтому сначала ацильная группа переносится с ацил-СоА на карнитин. Ацил-карнитин пересекает мембрану и отдает свой ацильный фрагмент снова на кофермент А. Ферментом этой обратимой реакции служит ацил-карнитинтрансфераза .
Первая фаза – активирование высшей жирной кислоты путем образования ацил КоА
![]() С17Н35СООН
+ НSКоА + АТФ С17Н35СО~
SКоА + АМФ + Н4Р2О7
С17Н35СООН
+ НSКоА + АТФ С17Н35СО~
SКоА + АМФ + Н4Р2О7
Стеариновая кислота
Фермент – ацилкоэнзим-А синтаза.
Вторая фаза состоит в окислении ацилКоА при участии ФАД -зависимой дегидрогеназы
![]()
![]() С15
Н31-СН2-СН2- СО~ SкоА
С15 Н31-СН = СН- СО~ SКоА
С15
Н31-СН2-СН2- СО~ SкоА
С15 Н31-СН = СН- СО~ SКоА
Стеарил КоА α, β дегидростеарил КоА
ФАД ФАДН2
Третья фаза – присоединение воды по месту двойной связи
![]() С15 Н31-СН=СН- СО~ SКоА
+ Н2О С15 Н31-СН–СН2-
СО~ SКоА
С15 Н31-СН=СН- СО~ SКоА
+ Н2О С15 Н31-СН–СН2-
СО~ SКоА
![]()
ОН
α, β оксистеарил КоА
фермент гидратаза.
Четвертая фаза – окисление – дегидрирование с участием НАД-зависимых дегидрогеназ
![]()
![]()
![]() С15 Н31-СН–СН2- СО~ SКоА
С15 Н31-С–СН2- СО~ SКоА
С15 Н31-СН–СН2- СО~ SКоА
С15 Н31-С–СН2- СО~ SКоА
![]()
![]()
ОН НАД НАДН2 О
β кетостеарил КоА
Пятая фаза распада- перенос новой ацильной группировки на НSКоА
![]()
![]()
![]() С15 Н31-С–СН2-
СО~ SКоА + НSКоА С15Н31СО~
SкоА
С15 Н31-С–СН2-
СО~ SКоА + НSКоА С15Н31СО~
SкоА
О
Пальмитил КоА
+ СН3СО~ SКоА
ацетил КоА
Фермент – тиолаза.
Т.О. кислота укорачивается на 2 углеродных атома. Далее цикл повторяется
Конечным продуктом распада жирных кислот с четным числом атомов – ацетил КоА, с нечетным - ацетил КоА и пропионил КоА.
СН3 ОН ОН
![]()
![]()
![]()
![]()
![]() НS-СН2-СН2-NН-СО-СН2-
СН2-NН-СО- СНОН-С- СН2-О-Р-О-Р-О-СН2-рибозо3фосфат-аденин
НS-СН2-СН2-NН-СО-СН2-
СН2-NН-СО- СНОН-С- СН2-О-Р-О-Р-О-СН2-рибозо3фосфат-аденин
СН3 О О
Коэнзим А – остаток тиоламина, остаток пантотеновой кислоты, остаток 3- фосфоаденозин-5-дифосфата.
Конечным итогом повторяющихся циклов β -окисления будет окисление всей ацильной цепи до ацетил-СоА, который затем может включаться в цитратный цикл для полного окисления.
Распад фосфатидов.1) Гидролитический путь. Фосфатиды распадаются на компоненты: глицерин, высшие жирные кислоты, фосфорную кислоту, азотистые вещества.
Ферменты – фосфолипазы – Классифицируются по месту расщепления и делятся на 4 типа:
А В С Д
С действия одного из этих ферментов начинается распад фосфатидов
|
СН2 |
|
О |
|
СО |
|
R1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
А |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
R2 |
|
|
|
|
|
|
|
|
|
|
СН2 |
|
|
|
|
|
В |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
СН2 |
|
СН2 |
|
|
СН2 |
|
|
С Д холин
Глицерин фосфорилируется и поступает в обменные реакции
ВЖК окисляется до СО2 и Н2О
Фосфорная кислота поступает в метаболитный фонд
Холин взаимодействует с ацетил-КоА – образуется ацетилхолин – вещество , осуществляющее передачу нервных импульсов.
Биосинтез фосфатидов.
Схема синтеза:
![]()
![]()
![]() глицерин
фосфоглицерин фосфатидная кислота
глицерин
фосфоглицерин фосфатидная кислота
![]()
![]() диглицерид ЦДФ
Х (цитидиндифосфатхолин)
диглицерид ЦДФ
Х (цитидиндифосфатхолин)
ФОСФАТИД + ЦМФ
![]() ЦДФ
+ холин ЦДФ-холин
ЦДФ
+ холин ЦДФ-холин
![]() ЦДФ-холин
+ диглицерид фосфатид + ЦМФ
ЦДФ-холин
+ диглицерид фосфатид + ЦМФ
Фермент – 1,2 диацилглицеролхолинфосфотрансфераза.
Биосинтез жирных кислот.
Схема синтеза высших жирных кислот
СО2 + ацетил КоА + АТФ + Н2О
![]()
Фермент – ацетил-КоА карбоксилаза
![]() СООН-
СН2 – СО –SКoA
+ АДФ + Н3РО4
СООН-
СН2 – СО –SКoA
+ АДФ + Н3РО4
малонилКоА
СООН- СН2 – СО –SкoA + ацетил КоА
Фермент трансацилаза
СН3- СО - СН2 – СО –SкoA + НSКоА
β - кетобутирил КоА
![]()
НАДН2 фермент кетоацилредуктаза
НАД
СН3- СНОН - СН2 – СО –SкoA
β- оксибутирил КоА
![]() фермент - дегидротаза
фермент - дегидротаза
Н2О
СН3- СН = СН – СО –SКoA
Кротонил КоА
![]()
![]() НАДФН2
НАДФН2
НАДФ+ фермент еноилредуктаза
СН3- СН2 - СН2 – СО –SКoA
бутирил КоА
Произошло удлинение цепи на два «С» атома, цикл продолжается дальше до образования высшей жирной кислоты.
Фермент ацетил – КоА-трансфераза
Проявляет свою активность при присоединении лимонной кислоты,
Фосфорилирование фермента понижает активность, дефосфорилирование – повышает.
Синтез триглицеридов. Синтез триацилглицеридов осуществляется путем трансацилирования
В синтезе участвуют :
α -фосфоглицерин
Ацил КоА
Ферменты – ацилтрансферазы
1 Образование α –фосфоглицерина
![]() глицерин
+ АТФ глицерофосфат
глицерин
+ АТФ глицерофосфат
2. Образование фосфатидной кислоты
![]() глицерофосфат
+2 R-СО-SКоА
фосфатидная кислота
глицерофосфат
+2 R-СО-SКоА
фосфатидная кислота
фермент ацилтрансфераза
![]() фосфатидная
кислота + Н2О диглицерид + Н3РО4
фосфатидная
кислота + Н2О диглицерид + Н3РО4
фермент – фосфатидатфосфогидролаза
3. Образование триглицерида
![]() диглицерид
+ R-СО-SКоА
триглицерид
диглицерид
+ R-СО-SКоА
триглицерид
фермент – ацилтрансфераза.
Обмен стеридов. При распаде, стериды гидролизуются на жирную кислоту и стерол. Реакция гидролиза ускоряется холестеролэстеразой, действующей также на сложные эфиры других стеролов.
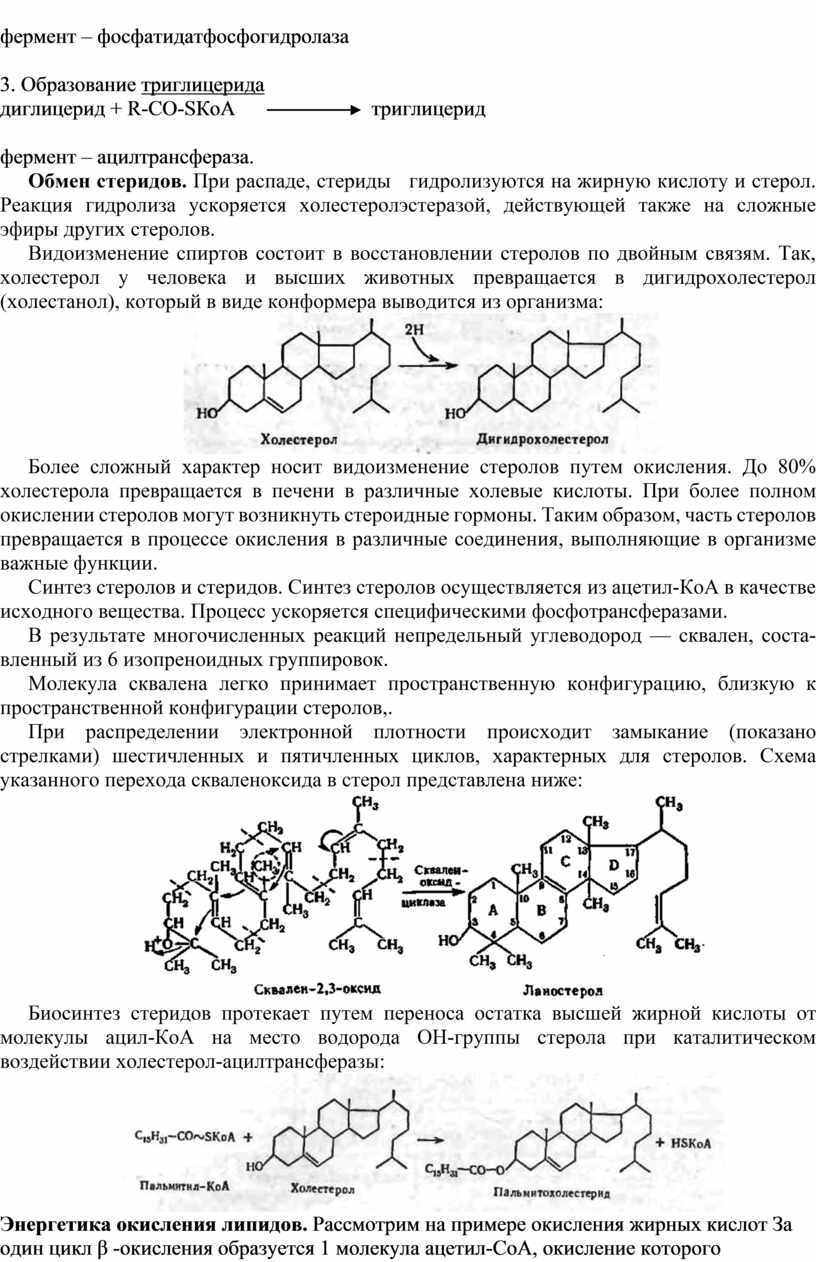
Видоизменение спиртов состоит в восстановлении стеролов по двойным связям. Так, холестерол у человека и высших животных превращается в дигидрохолестерол (холестанол), который в виде конформера выводится из организма:

Более сложный характер носит видоизменение стеролов путем окисления. До 80% холестерола превращается в печени в различные холевые кислоты. При более полном окислении стеролов могут возникнуть стероидные гормоны. Таким образом, часть стеролов превращается в процессе окисления в различные соединения, выполняющие в организме важные функции.
Синтез стеролов и стеридов. Синтез стеролов осуществляется из ацетил-КоА в качестве исходного вещества. Процесс ускоряется специфическими фосфотрансферазами.
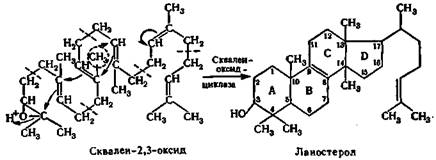
В результате многочисленных реакций непредельный углеводород — сквален, составленный из 6 изопреноидных группировок.
Молекула сквалена легко принимает пространственную конфигурацию, близкую к пространственной конфигурации стеролов,.
При распределении электронной плотности происходит замыкание (показано стрелками) шестичленных и пятичленных циклов, характерных для стеролов. Схема указанного перехода скваленоксида в стерол представлена ниже:
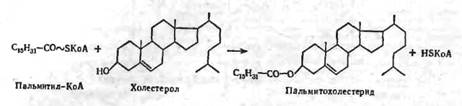
Биосинтез стеридов протекает путем переноса остатка высшей жирной кислоты от молекулы ацил-КоА на место водорода ОН-группы стерола при каталитическом воздействии холестерол-ацилтрансферазы:
Энергетика окисления липидов. Рассмотрим на примере окисления жирных кислот За один цикл β -окисления образуется 1 молекула ацетил-СоА, окисление которого в цитратном цикле обеспечивает синтез 12 моль АТФ. Кроме того, образуется 1 моль ФАДH2 и 1 моль НАДH, при окислении которых в дыхательной цепи синтезируется соответственно 2 и 3 моль АТФ (в сумме 5).
Таким образом, при окислении, например, пальмитиновой кислоты (С16) происходит 7 циклов β -окисления, в результате которых образуется 8 молей ацетил-СоА, 7 молей ФАДH2 и 7 молей НАДH+Н. Следовательно, выход АТФ составляет 35 АТФ в результате β -окисления и 96 АТФ в результате цитратного цикла, что соответствует в сумме 131 моль АТФ.
Скачано с www.znanio.ru







Материалы на данной страницы взяты из открытых источников либо размещены пользователем в соответствии с договором-офертой сайта. Вы можете сообщить о нарушении.