Поделиться

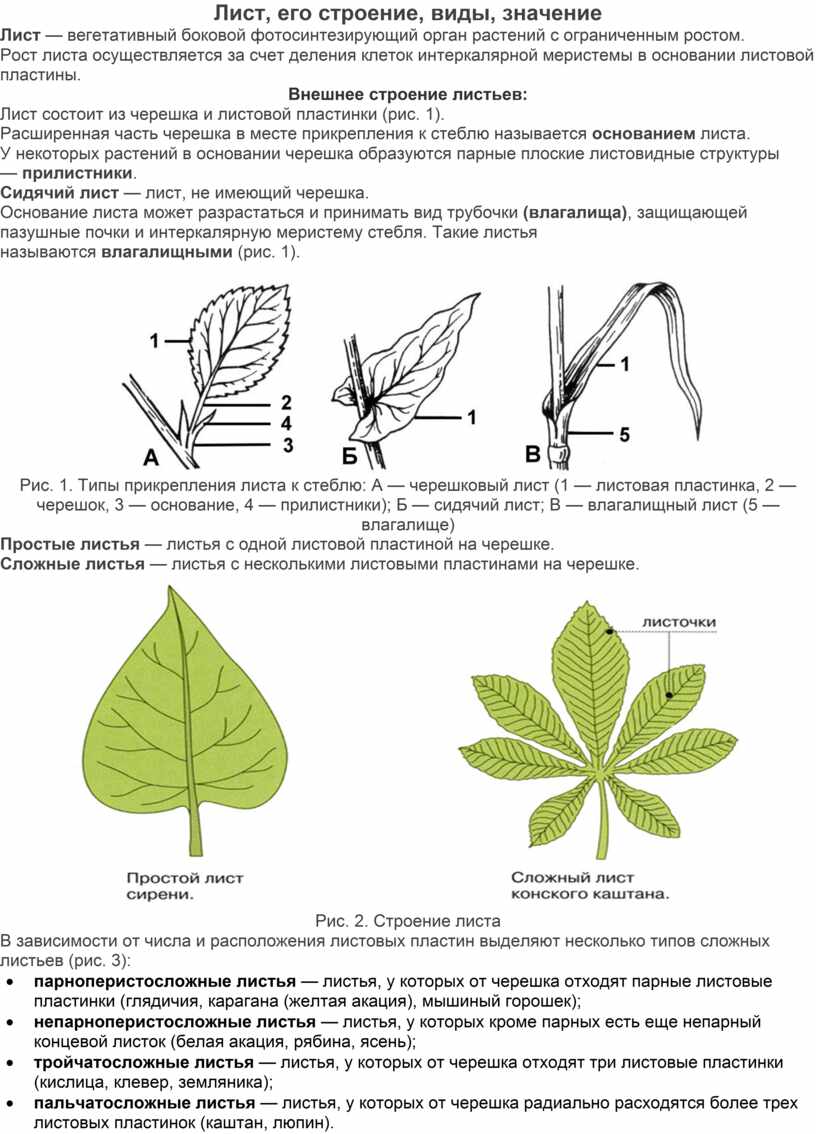
Лист, его строение, виды, значение
Лист — вегетативный боковой фотосинтезирующий орган растений с ограниченным ростом.
Рост листа осуществляется за счет деления клеток интеркалярной меристемы в основании листовой пластины.
Лист состоит из черешка и листовой пластинки (рис. 1).
Расширенная часть черешка в месте прикрепления к стеблю называется основанием листа.
У некоторых растений в основании черешка образуются парные плоские листовидные структуры — прилистники.
Сидячий лист — лист, не имеющий черешка.
Основание листа может разрастаться и принимать вид трубочки (влагалища), защищающей пазушные почки и интеркалярную меристему стебля. Такие листья называются влагалищными (рис. 1).
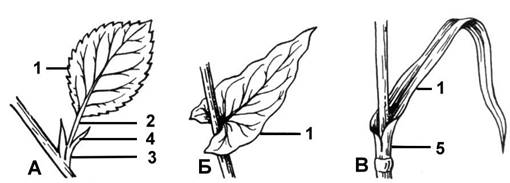
Рис. 1. Типы прикрепления листа к стеблю: A — черешковый лист (1 — листовая пластинка, 2 — черешок, 3 — основание, 4 — прилистники); Б — сидячий лист; В — влагалищный лист (5 — влагалище)
Простые листья — листья с одной листовой пластиной на черешке.
Сложные листья — листья с несколькими листовыми пластинами на черешке.
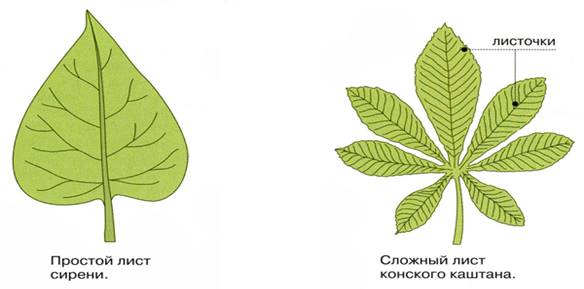
Рис. 2. Строение листа
В зависимости от числа и расположения листовых пластин выделяют несколько типов сложных листьев (рис. 3):
· парноперистосложные листья — листья, у которых от черешка отходят парные листовые пластинки (глядичия, карагана (желтая акация), мышиный горошек);
· непарноперистосложные листья — листья, у которых кроме парных есть еще непарный концевой листок (белая акация, рябина, ясень);
· тройчатосложные листья — листья, у которых от черешка отходят три листовые пластинки (кислица, клевер, земляника);
· пальчатосложные листья — листья, у которых от черешка радиально расходятся более трех листовых пластинок (каштан, люпин).

Рис. 3. Сложные листья: 1 — непарноперистосложный; 2 — парноперистосложный; 3 — пальчатосложный; 4 — тройчатосложный; 5 — дваждытройчатосложный; 6 — дваждыперистосложный
Жилки — сосудисто-волокнистые пучки, осуществляющие транспорт веществ в листовой пластине.
Расположение жилок в листовой пластинке называется жилкованием (рис. 4).
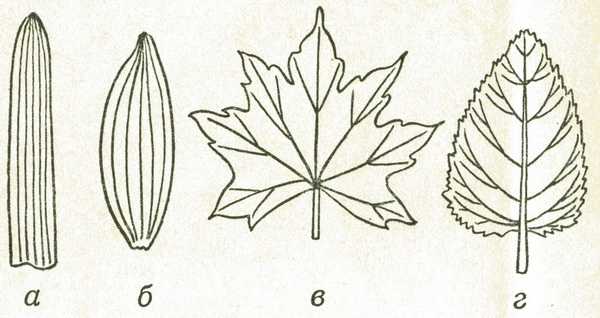
Рис. 4. Жилкование листа: а — параллельное, б — дуговидное, в — пальчатое, г — перистое
Жилкование у большинства папоротников и примитивных семенных растений (гинкго) дихотомическое, т. е. вильчатое: жилки делятся на двое, затем снова на двое и т. д. (рис. 5).

Рис. 5. Дихотомическое жилкование листа гинкго
У большинства хвойных в листе проходит одна или несколько продольных не связанных между собой жилок.
Параллельное и дуговое жилкование характерно для однодольных растений. В листовую пластинку входят сразу несколько жилок, проходящих вдоль всего листа, не пересекаясь. Если пластинка узкая, они идут параллельно друг другу (пшеница, кукуруза). Если же листовая пластинка широкая, жилки принимают дугообразную форму (ландыш, подорожник).
Пальчатое и перистое жилкование часто объединяют под названием сетчатое, оно характерно для двудольных растений.
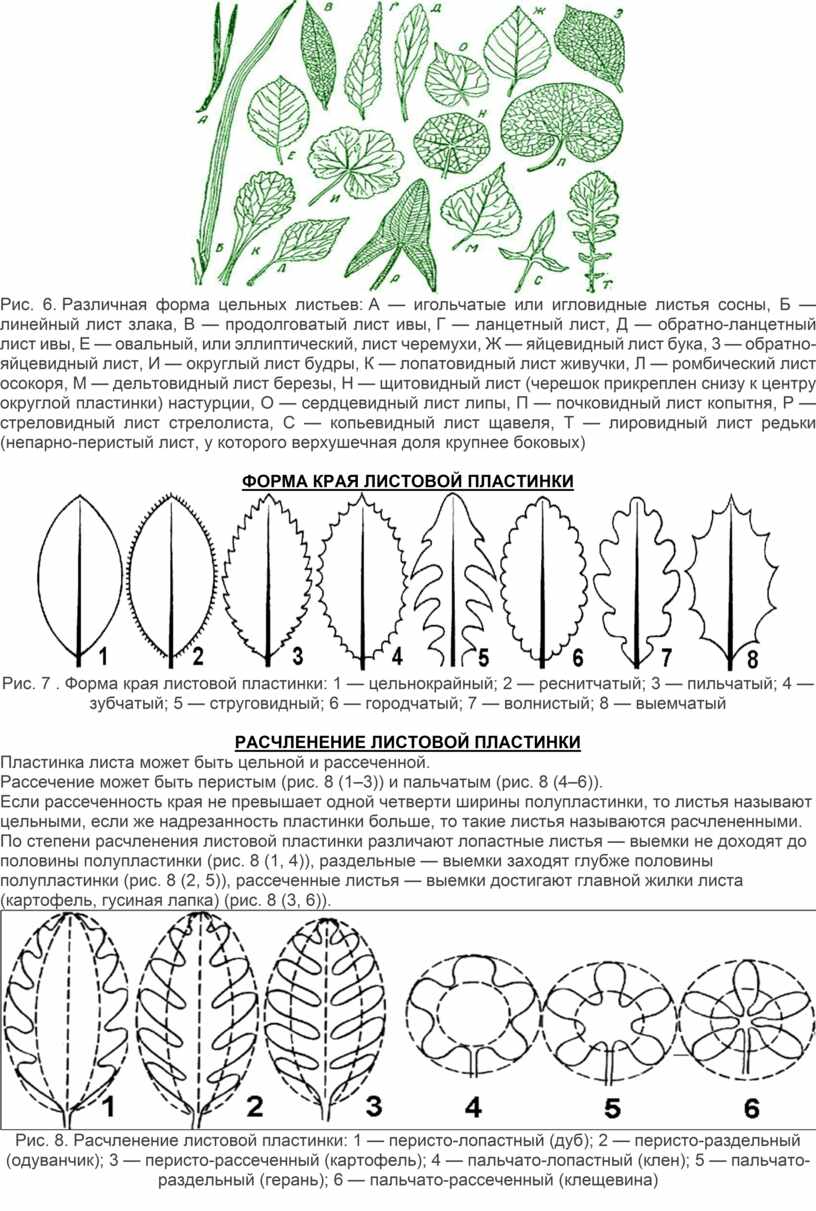
По форме листовые пластинки бывают округлые, овальные, эллиптические, яйцевидные, линейные, ланцетные, стреловидные, копьевидные и другие (рис. 6).
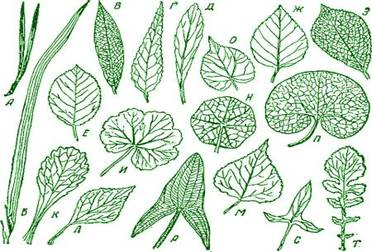
Рис. 6. Различная форма цельных листьев: А — игольчатые или игловидные листья сосны, Б — линейный лист злака, В — продолговатый лист ивы, Г — ланцетный лист, Д — обратно-ланцетный лист ивы, Е — овальный, или эллиптический, лист черемухи, Ж — яйцевидный лист бука, 3 — обратно-яйцевидный лист, И — округлый лист будры, К — лопатовидный лист живучки, Л — ромбический лист осокоря, М — дельтовидный лист березы, Н — щитовидный лист (черешок прикреплен снизу к центру округлой пластинки) настурции, О — сердцевидный лист липы, П — почковидный лист копытня, Р — стреловидный лист стрелолиста, С — копьевидный лист щавеля, Т — лировидный лист редьки (непарно-перистый лист, у которого верхушечная доля крупнее боковых)
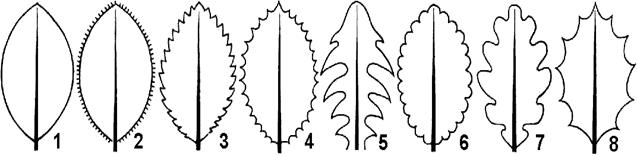
Рис. 7 . Форма края листовой пластинки: 1 — цельнокрайный; 2 — реснитчатый; 3 — пильчатый; 4 — зубчатый; 5 — струговидный; 6 — городчатый; 7 — волнистый; 8 — выемчатый
Пластинка листа может быть цельной и рассеченной.
Рассечение может быть перистым (рис. 8 (1–3)) и пальчатым (рис. 8 (4–6)).
Если рассеченность края не превышает одной четверти ширины полупластинки, то листья называют цельными, если же надрезанность пластинки больше, то такие листья называются расчлененными.
По степени расчленения листовой пластинки различают лопастные листья — выемки не доходят до половины полупластинки (рис. 8 (1, 4)), раздельные — выемки заходят глубже половины полупластинки (рис. 8 (2, 5)), рассеченные листья — выемки достигают главной жилки листа (картофель, гусиная лапка) (рис. 8 (3, 6)).
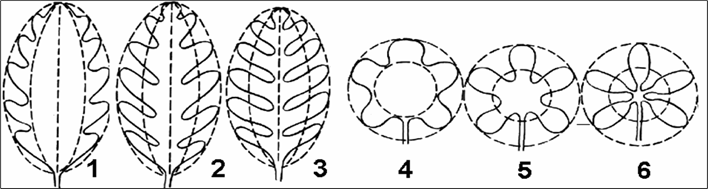
Рис. 8. Расчленение листовой пластинки: 1 — перисто-лопастный (дуб); 2 — перисто-раздельный (одуванчик); 3 — перисто-рассеченный (картофель); 4 — пальчато-лопастный (клен); 5 — пальчато-раздельный (герань); 6 — пальчато-рассеченный (клещевина)
Листорасположение — это расположение листьев на стебле (рис. 9).
Очередное листорасположение: листья располагаются будто по растянутой спирали, окружающей стебель, по очереди друг за другом (алоэ, береза, яблоня, роза).
Супротивное листорасположение: листья на стебле располагаются парами (каланхоэ, сирень, яснотка, мята).
Мутовчатое листорасположение: листья прикрепляются к стеблю мутовками — пучками (олеандр, вороний глаз, элодея, можжевельник).
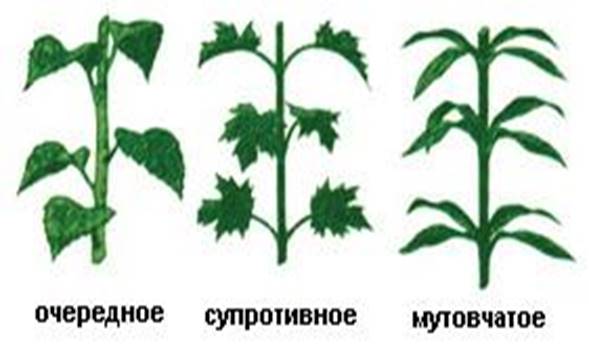
Рис. 9. Листорасположение
Пластинку листа образуют 4 группы тканей:
· покровная — кожица, или эпидермис;
· основная (фотосинтезирующая (ассимилирующая) паренхима) — мезофилл;
· проводящая — сосудисто-волокнистые пучки (жилки);
· механическая — древесные и лубяные волокна жилок, придающие листу жесткость и прочность.
Лист окружен со всех сторон покровной тканью — эпидермисом, или кожицей (рис. 10).
Кожица состоит из одного слоя плотно сомкнутых живых клеток без межклетников. Наружные стенки этих клеток сильно утолщены и выделяют воскоподобные вещества, образующие кутикулу, т. к. их основная функция — защита листа от механических повреждений и избыточной транспирации.
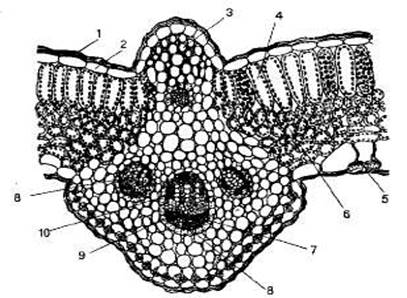
Рис. 10. Внутреннее строение листа: 1 — кутикула; 2 — эпидермис; 3 — колленхима; 4 — палисадная (столбчатая) паренхима; 5 — устьице; 6 — губчатая паренхима; 7 — флоэма; 8 — проводящий пучок; 9 — склеренхима; 10 — ксилема
Клетки кожицы не содержат хлорофилловых зерен, следовательно, не фотосинтезируют. Однако они свободно пропускают солнечные лучи вглубь, к фотосинтезирующей паренхиме мезофилла листа.
Кроме кожицы лист может быть покрыт восковым слоем и кутикулой. Наличие и толщина этих слоев зависят от условий произрастания растения, т. к. они защищают в первую очередь от излишней транспирации (испарения воды). Но, у клеток верхней кожицы она обычно более значительна, чем у клеток нижней кожицы. Волоски у опушенных листьев располагаются преимущественно на нижней стороне листа.
Эпидермис практически непроницаем для газов. Дыхание и транспирация осуществляются через устьица.
Устьице — пора в эпидермисе листа (рис. 11).
Пора состоит из пары замыкающих клеток, которые содержат хлоропласты и способны к фотосинтезу (рис. 12). Между ними располагается устьичная щель. Стенки замыкающих клеток утолщены неравномерно: окаймляющие щель стенки более толстые. Щель может расширяться и сужаться, регулируя транспирацию и газообмен. Когда воды мало, замыкающие клетки плотно прилегают друг к другу и устьичная щель закрыта. Когда воды в замыкающих клетках много, то она давит на стенки, и более тонкие стенки растягиваются сильнее, а более толстые втягиваются внутрь, между замыкающими клетками появляется щель.
Под щелью расположена воздушная полость (крупный межклетник), окруженная клетками мякоти листа, через которую непосредственно и происходит газообмен. Воздух проникает внутрь ткани листа через эти поры и далее используется в процессе фотосинтеза и дыхании. Избыточный кислород, произведенный в процессе фотосинтеза внутренними клетками листа, выходит обратно в окружающую среду через эти же поры. Также в процессе испарения через поры выделяются пары воды. Клетки эпидермиса, примыкающие к замыкающим, получили название сопровождающих (околоустьичных). Они участвуют в движении замыкающих клеток. Замыкающие и сопровождающие клетки образуют устьичный аппарат.
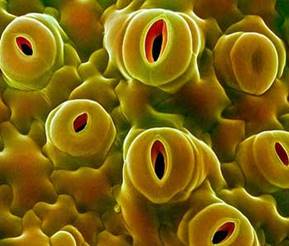
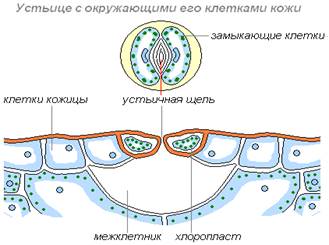
Рис. 11. Устьица на поверхности листа Рис. 12. Строение устьица
Устьица располагаются на обеих сторонах листа (у многих злаков), только на верхней стороне (у части водных растений), но чаще всего они располагаются только на нижней стороне.
Мезофилл занимает все внутреннее пространство листа, исключая проводящие пучки и участки механической ткани. Клетки мезофилла состоят из основной фотосинтезирующей паренхимы двух видов: столбчатой (полисадной) и губчатой.
Столбчатый мезофилл расположен под верхним слоем эпидермиса и состоит из вертикально вытянутых, плотно расположенных клеток, содержащих большое количество хлорофилла. Столбчатый мезофилл является основной фотосинтезирующей тканью листа.
Губчатый мезофилл состоит из рыхло расположенных фотосинтезирующих клеток с большими межклетниками, которые способствуют свободному газообмену всех слоев мезофилла с внешней средой. Таким образом, углекислый газ через устьица и межклетники губчатого мезофилла поступает в столбчатый мезофилл, а кислород, образующийся в процессе фотосинтеза, свободно выходит из листа в атмосферу.
За счет рыхлости ткани площадь внутренней поверхности мезофилла значительно больше площади поверхности листа, что увеличивает интенсивность фотосинтеза.
У некоторых растений поверхность мезофилла увеличивается за счет складок клеток, вдающихся внутрь, например складчатая паренхима хвойных.
У растений умеренной зоны, не испытывающих недостатка влаги, столбчатый мезофилл располагается на верхней стороне листа, а губчатый — на нижней. У сухолюбивых растений столбчатый мезофилл располагается и с верхней и с нижней стороны листовой пластины.
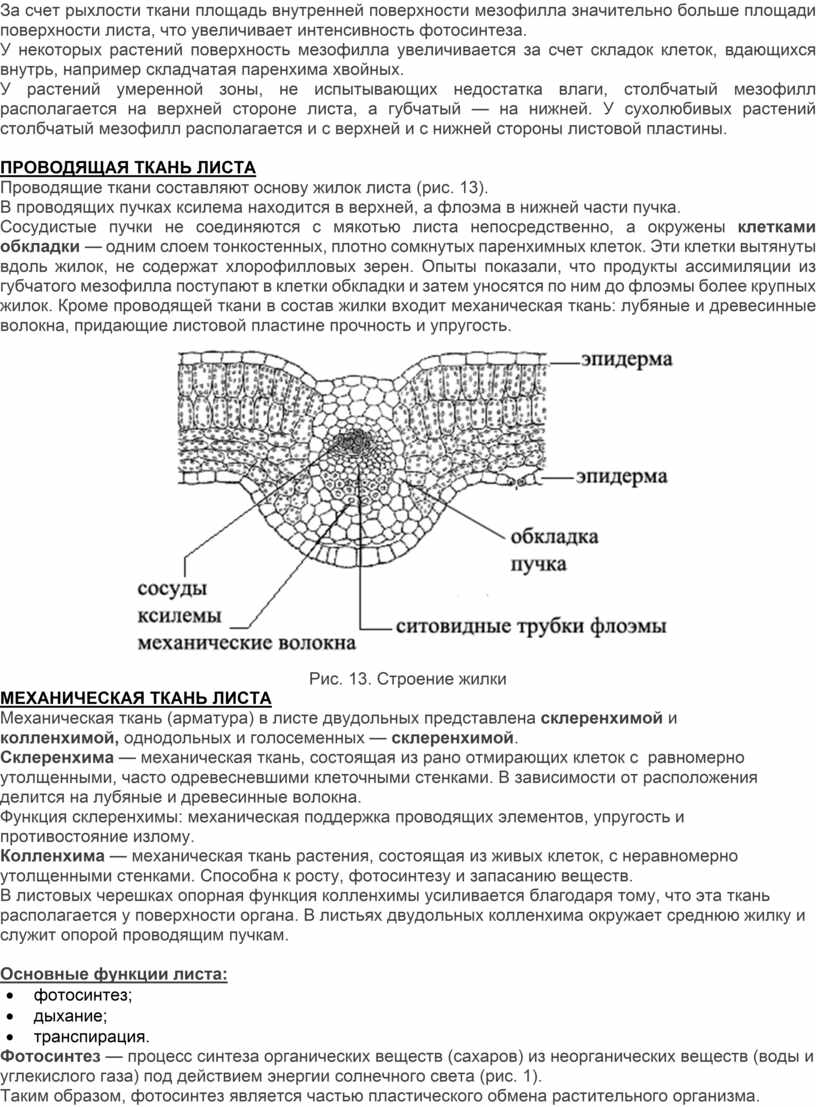
Проводящие ткани составляют основу жилок листа (рис. 13).
В проводящих пучках ксилема находится в верхней, а флоэма в нижней части пучка.
Сосудистые пучки не соединяются с мякотью листа непосредственно, а окружены клетками обкладки — одним слоем тонкостенных, плотно сомкнутых паренхимных клеток. Эти клетки вытянуты вдоль жилок, не содержат хлорофилловых зерен. Опыты показали, что продукты ассимиляции из губчатого мезофилла поступают в клетки обкладки и затем уносятся по ним до флоэмы более крупных жилок. Кроме проводящей ткани в состав жилки входит механическая ткань: лубяные и древесинные волокна, придающие листовой пластине прочность и упругость.
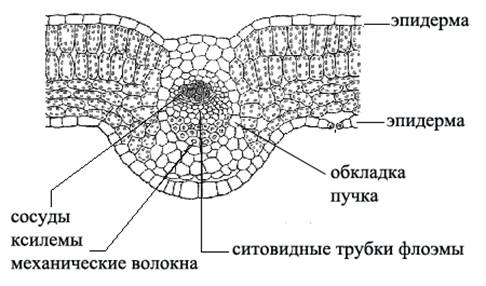
Рис. 13. Строение жилки
Механическая ткань (арматура) в листе двудольных представлена склеренхимой и колленхимой, однодольных и голосеменных — склеренхимой.
Склеренхима — механическая ткань, состоящая из рано отмирающих клеток с равномерно утолщенными, часто одревесневшими клеточными стенками. В зависимости от расположения делится на лубяные и древесинные волокна.
Функция склеренхимы: механическая поддержка проводящих элементов, упругость и противостояние излому.
Колленхима — механическая ткань растения, состоящая из живых клеток, с неравномерно утолщенными стенками. Способна к росту, фотосинтезу и запасанию веществ.
В листовых черешках опорная функция колленхимы усиливается благодаря тому, что эта ткань располагается у поверхности органа. В листьях двудольных колленхима окружает среднюю жилку и служит опорой проводящим пучкам.
Основные функции листа:
· фотосинтез;
· дыхание;
· транспирация.
Фотосинтез — процесс синтеза органических веществ (сахаров) из неорганических веществ (воды и углекислого газа) под действием энергии солнечного света (рис. 1).
Таким образом, фотосинтез является частью пластического обмена растительного организма.

Рис. 1. Схема фотосинтеза
В процессе фотосинтеза участвуют особые вещества — фотосинтетические пигменты — улавливающие солнечный свет и трансформирующие энергию солнечного света в энергию химических связей. У зеленых растений основной фотосинтетический пигмент — хлорофилл. Хлорофилл содержится в хлоропластах, в мембранных пузырьках (тилакоидах), собранных в стопки (граны) (рис. 2).
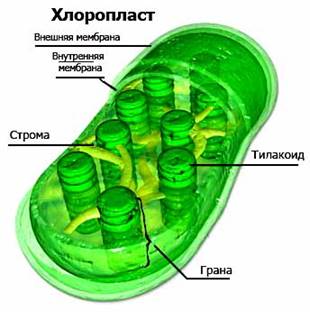
Рис. 2. Строение хлоропласта
Пигменты:
· хлорофилл a (для всех растений и многих цианобактерий);
· хлорофилл b (у сосудистых растений, мхов, зеленых и эвгленовых водорослей);
· хлорофилл с (в основном у бурых и диатомовых водорослей);
· хлорофилл d (у многих красных водорослей);
· бактериохлорофилл (у бактерий);
· каротиноиды — это красные, оранжевые и желтые жирорастворимые пигменты (в хлоропластах и у цианобактерий);
· фикобилины (у цианобактерий и красных водорослей).
Фотосинтез включает 2 фазы (рис. 3)
Проходит на мембранах тилакоидов хлоропластов, необходим солнечный свет, под действием энергии солнечного света происходит распад молекул воды (фотолиз) с последующим образованием свободного кислорода, как побочного продукта и богатых энергией молекулы АТФ.
Проходит в строме хлоропластов, может происходить без солнечного света; необходима энергия АТФ, запасенная в световую фазу, синтез органических веществ (глюкозы) из неорганических веществ (углекислого газа и воды), поступающих из окружающей среды, с использованием энергии АТФ (из световой фазы).
· Синтез органических веществ для организма растения и всех гетеротрофных организмов планеты.
· Выделение кислорода как побочного продукта световой фазы фотосинтеза.
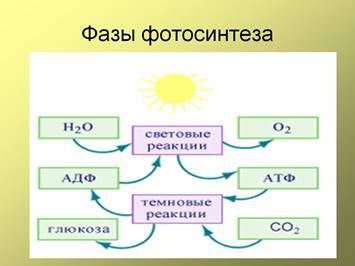
Рис. 3. Фазы фотосинтеза
Дыхание(окисление) — процесс поглощения кислорода и выделения углекислого газа.
Процесс
дыхания связан с непрерывным потреблением кислорода днем и ночью.
Полученный в процессе дыхания кислород расходуется организмом на окисление
органических соединений с получением энергии для жизнедеятельности.
Таким образом, процесс дыхания — это часть энергетического
обмена растительного организма. Интенсивность дыхания обусловлена
потребностями роста и развития растений. Особенно интенсивно идет процесс
дыхания в молодых тканях и органах растения. Много кислорода требуется для
деления и роста клеток, образования цветков и плодов. По окончании роста, с
пожелтением листьев и особенно в зимнее время интенсивность дыхания заметно
снижается, но не прекращается.
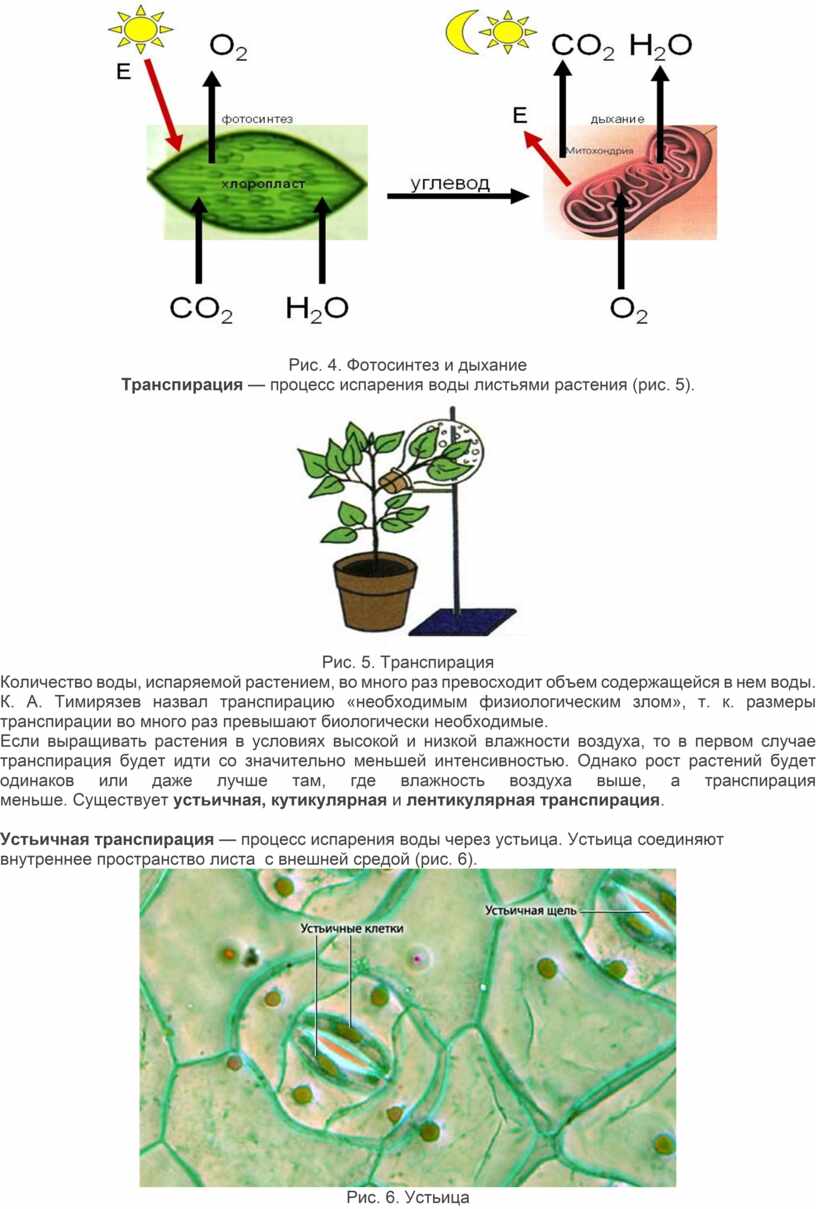
Процесс дыхания противоположен процессу фотосинтеза (рис. 4).
|
Фотосинтез |
Дыхание |
|
Поглощение углекислого газа |
Поглощение кислорода |
|
Выделение кислорода |
Выделение углекислого газа |
|
Образование сложных органических веществ (преимущественно сахаров) из простых неорганических |
Разложение сложных органических веществ (преимущественно сахаров) на простые неорганические |
|
Поглощение из окружающей среды и расходование воды |
Образование и выделение в окружающую среду воды |
|
Поглощение с помощью хлорофилла солнечной энергии и накопление ее в органических веществах |
Высвобождение энергии |
|
Происходит только на свету |
Происходит непрерывно, на свету и в темноте |
|
Протекает в хлоропластах |
Протекает в цитоплазме и митохондриях |
|
Происходит только в зеленых частях растения, преимущественно в листе |
Происходит в клетках всех органов растения |
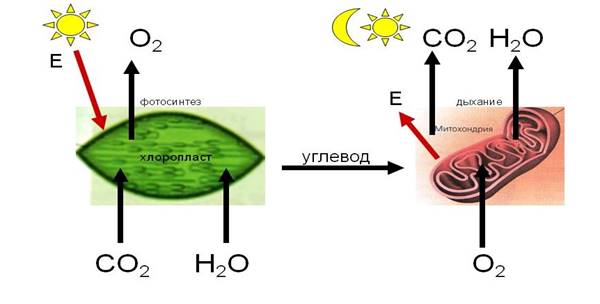
Рис. 4. Фотосинтез и дыхание
Транспирация — процесс испарения воды листьями растения (рис. 5).

Рис. 5. Транспирация
Количество воды, испаряемой растением, во много раз превосходит объем содержащейся в нем воды. К. А. Тимирязев назвал транспирацию «необходимым физиологическим злом», т. к. размеры транспирации во много раз превышают биологически необходимые.
Если выращивать растения в условиях высокой и низкой влажности воздуха, то в первом случае транспирация будет идти со значительно меньшей интенсивностью. Однако рост растений будет одинаков или даже лучше там, где влажность воздуха выше, а транспирация меньше. Существует устьичная, кутикулярная и лентикулярная транспирация.
Устьичная транспирация — процесс испарения воды через устьица. Устьица соединяют внутреннее пространство листа с внешней средой (рис. 6).
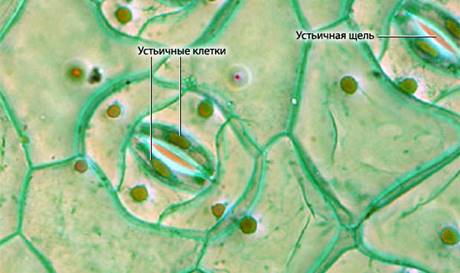
Рис. 6. Устьица
Факторы, влияющие на устьичную транспирацию:
· температура воздуха;
· движение воздуха в окружающей среде;
·
активность процесса фотосинтеза.
В отличие от других клеток эпидермиса, замыкающие клетки устьиц содержат
хлоропласты. Синтез углеводов в процессе фотосинтеза в замыкающих клетках
увеличивает их сосущую силу и вызывает поглощение воды, способствуя этим
открыванию устьиц.
·
концентрация углекислого газа.
Ночью в межклетниках листа в результате отсутствия фотосинтеза и
продолжающегося дыхания уровень углекислого газа в тканях повышается. Повышение
концентрации CO2CO2 вызывает закрытие устьиц. Это объясняет,
почему ночью устьица закрыты и открываются с восходом солнца.
· Биологический ритм транспирации бывает двух вариантов:
· У деревьев, теневыносливых растений, многих злаков и других видов с совершенной регуляцией устьичной транспирации (гидростабильных видов) испарение воды имеет два максимума — в утренние и в предвечерние часы (при снижении температуры воздуха).
· У видов растений, способных переносить резкие изменения содержания воды в клетках в течение дня (гидролабильных видов) наблюдается один максимум в полуденные часы.
· В обоих случаях ночью транспирация минимальна или полностью прекращается.
Кутикулярная транспирация — процесс испарения воды через кутикулу листа (10–30 % от всей транспирации).
· Факторы, влияющие на кутикулярную транспирацию:
· Имеются данные, что кутикулярная транспирация меньше зависит от условий внешней среды по сравнению с устьичной.
Лентикулярная транспирация — транспирация через чечевички на поверхности стебля (к листовой транспирации не относится).
В процессе транспорта минеральных веществ от корней к листьям притекает большой избыток воды, которая должна из растения удаляться. Поэтому клетки губчатого мезофилла интенсивно испаряют воду в межклетники (рис. 7). Образовавшийся пар выходит через устьица (до 90 %) и кутикулу (до 30 %). Таким образом обеспечивается работа насоса, подающего минеральные вещества от корней к листьям.
Первый этап — переход воды из клеточных оболочек в межклетники, т. е. из капельножидкого состояния в парообразное (испарения с поверхности клеточных стенок).
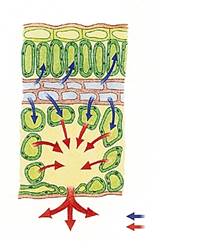
Рис. 7. Схема транспирации: синяя стрелка — жидкая вода, красная стрелка — пары воды
Важно подчеркнуть, что уже на этом этапе растение обладает способностью регулировать процесс транспирации. Между всеми частями клетки существует водное равновесие. Чем меньше воды в клетке, тем выше становится концентрация клеточного сока. Уменьшается содержание свободной воды в протопласте и клеточной оболочке. Водоудерживающая сила растет, интенсивность испарения уменьшается.
Второй этап — это выход паров воды из межклетников через устьица или через кутикулу.
Поверхность всех клеточных стенок, соприкасающихся с межклетными пространствами, превышает поверхность листа примерно в 10–30 раз. Если устьица закрыты, то все это пространство быстро насыщается парами воды, и переход воды из жидкого в парообразное состояние прекращается. Иная картина наблюдается при открытых устьицах. Как только часть паров воды выйдет из межклетников через устьичные щели, так сейчас же этот недостаток восполняется за счет испарения воды с поверхности клеток.
Поскольку устьичная транспирация составляет 80–90 % от всего испарения листа, то степень открытости устьиц является основным механизмом, регулирующим интенсивность транспирации. Именно на этом этапе вступает в действие устьичная регулировка транспирации. При недостатке воды в листе устьица автоматически закрываются. Полное закрывание устьиц сокращает транспирацию на 90 %.
Третий этап — это диффузия паров воды от поверхности листа в более далекие слои атмосферы. Этот этап регулируется лишь условиями внешней среды.
1. Терморегуляция. Транспирация спасает растение от перегрева. Температура живого (транспирирующего) листа на несколько градусов ниже температуры завядшего (нетранспирирующего) листа. Перегрев, разрушая хлоропласты, резко снижает процесс фотосинтеза (оптимальная температура для процесса фотосинтеза — 20—25 °С). Именно благодаря высокой транспирирующей способности многие растения хорошо переносят повышенную температуру.
2. Минеральное (корневое питание) питание растения. Транспирация участвует в создании непрерывного тока воды с растворенными минеральными и органическими соединениями из корневой системы к надземным органам растения.
3. Транспорт веществ в растении (саморегуляция и обмен веществ). Постоянный ток воды по сосудам связывает все органы растения в единое целое, позволяя растению функционировать как саморегулирующаяся биосистема.
В случае невозможности испарения воды (100 % влажность воздуха во влажных тропических лесах) в листьях формируются специальные структуры — гидатоды (рис. 8), выделяющие через устьица жидкую воду. Это явление называется гуттация (рис. 9).
Гуттация (лат. gutta — капля) — процесс выведения воды в виде капель жидкости на поверхности растения гидатодами.
Факторы, способствующие гуттации: слабая освещенность и высокая влажность воздуха. Это явление наблюдается у многих растений влажных тропических лесов и часто наблюдается на кончиках листьев молодых проростков.
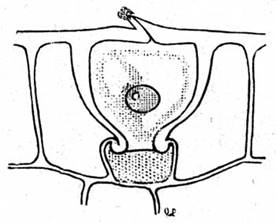

Рис. 8. Гидатода Рис. 9. Гуттация листа манжетки
В ряде случаев листья помимо основных начинают выполнять дополнительные функции. В этом случае часто они изменяют свое строение, превращаясь в различные видоизменения.
У многих лазающих растений часть листьев превращается в усики (рис. 10). Они закручиваются вокруг различных предметов, закрепляя стебель в поднятом состоянии (горох).
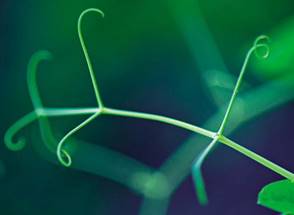
Рис. 10. Усики гороха
У других растений лист превращается в колючку (кактусы), выполняя защитную функцию (рис. 11). Такие видоизмененные листья не могут выполнять функции фотосинтеза.

Рис. 11. Колючки кактуса
В ряде случаев листья берут на себя запасающую функцию (рис. 12). В этом случае происходит разрастание мезофилла, внутренние его слои не получают света и не фотосинтезируют, но в них могут накапливаться питательные вещества (капуста) или вода (толстянковые).

Рис. 12. Утолщенные листья толстянковых
Интересную группу представляют собой ловчие листья насекомоядных растений. Как правило, такие растения произрастают в условиях дефицита азотного питания (болотные и эпифитные растения). Нехватку азота они компенсируют, переваривая насекомых, для ловли которых приспособлены их листья. У росянок (рис. 13) поверхность листьев образует многочисленные выросты, на концах которых выделяются капли вязкой жидкости, содержащие пищеварительные ферменты. Насекомое, севшее на такой лист, прилипает, лист свертывается, смачивая насекомое пищеварительным соком. В основном в нем содержатся ферменты, расщепляющие белки, образующиеся в результате аминокислоты всасываются листьями и служат источником азота.
У других насекомоядных растений, например у венериной мухоловки (рис. 14), листья способны при прикосновении к их верхней поверхности достаточно быстро сворачиваться по средней жилке, при этом насекомое оказывается зажатым в «клетке» из выростов листа. Со временем оно умирает и переваривается растением. Такие ловчие листья обычно содержат хлорофилл и участвуют в фотосинтезе.


Рис. 13. Лист росянки Рис. 14. Лист венериной мухоловки









Материалы на данной страницы взяты из открытых источников либо размещены пользователем в соответствии с договором-офертой сайта. Вы можете сообщить о нарушении.
