Известно, что информация о последовательности аминокислот в белке закодирована в генетическом аппарате клетки последовательностью дезоксирибонуклеотидных остатков в молекуле ДНК. Молекулы мРНК в качестве матрицы поступают в рибосому. Здесь она переводится (транслируется) с полинуклеотидной последовательности в аминокислотную последовательность новообразуемого в рибосомальном аппарате белка.
Матричный синтез белков состоит их этапов
1.Рекогниция – активироание аминокислот
2. Инициация.
3. Элонгация
4. Терминация
Рекогниция-активирование аминокислот и перенос их в рибосому. Синтез пептидной связи из свободных аминокислот протекает с поглощением энергии в количестве около 12 кДж/моль. В связи с этим давно уже была высказана идея о сопряженности биосинтеза белка с окислительными процессами (А.В. Благовещенский, М.П. Юргенсон, 1937) или с распадом соединений, содержащих макроэргические связи (Ф. Липман, 1941. М. Хогленд впервые предложил общепринятую сейчас двухэтапную схему активирования аминокислот.
Метаболизм белков.
1.
2.
МАТРИЧНАЯ СИСТЕМА СИНТЕЗА БЕЛКА.
ЭТАПЫ МАТРИЧНОГО СИНТЕЗА
Общая схема матричного биосинтеза белковых тел складывается из подготовительных
процессов — переноса вещества, энергии и информации в рибосому, и главного центрального
процесса — сборки полипептидных цепей в рибосоме.
В результате транскрипции (переписывание) информации о порядке расположения
аминокислотных остатков в молекуле синтезируемого белка перенесена с ДНК на мРНК.
(рассмотрено ранее).
Известно, что информация о последовательности аминокислот в белке закодирована в
генетическом аппарате клетки последовательностью дезоксирибонуклеотидных остатков в
молекуле ДНК. Молекулы мРНК в качестве матрицы поступают в рибосому. Здесь она
переводится (транслируется) с полинуклеотидной последовательности в аминокислотную
последовательность новообразуемого в рибосомальном аппарате белка.
Матричный синтез белков состоит их этапов
1.Рекогниция – активироание аминокислот
2. Инициация.
3. Элонгация
4. Терминация
Рекогницияактивирование аминокислот и перенос их в рибосому. Синтез
пептидной связи из свободных аминокислот протекает с поглощением энергии в количестве
около 12 кДж/моль. В связи с этим давно уже была высказана идея о сопряженности
биосинтеза белка с окислительными процессами (А.В. Благовещенский, М.П. Юргенсон, 1937)
или с распадом соединений, содержащих макроэргические связи (Ф. Липман, 1941. М.
Хогленд впервые предложил общепринятую сейчас двухэтапную схему активирования
аминокислот.
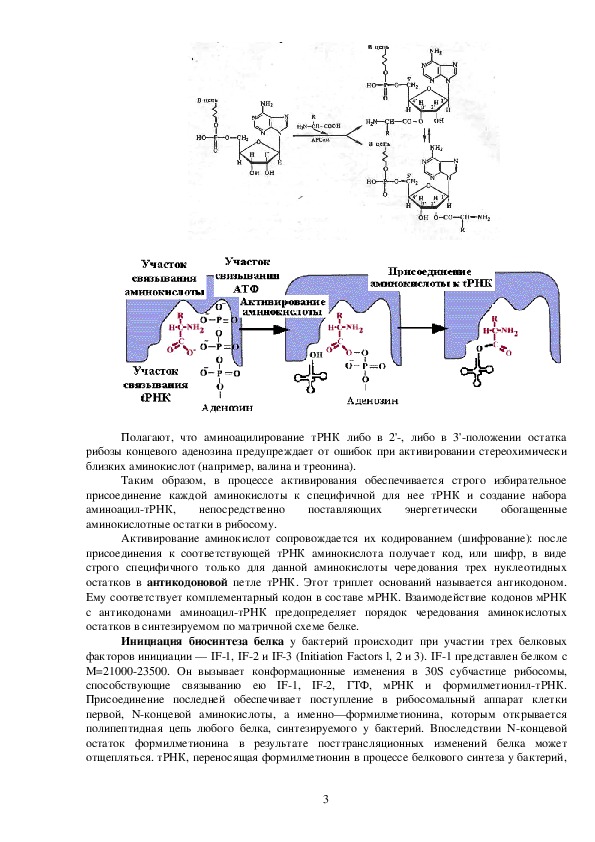
Первый ее этап состоит во взаимодействии аминокислоты с АТФ, в результате чего
возникает аминоациладенилат и выделяется пирофосфат.
Второй этап
сводится к переносу аминокислоты с образовавшегося
аминоациладенилата на концевой аденозин акцептирующего стебля тРНК:
Как видно из структурной формулы аминоациладенилата, он представляет ангидрид
аминокислоты и остатка фосфорной кислоты аденозин5'фосфата, причем донором
кислорода для ангидридной связи является ОНгруппа карбоксила аминокислоты. Свободные
аминоациладенилаты с легкостью вступают в реакцию с аминокислотами даже при очень
небольшой концентрации (103 моль), при почти нейтральной реакции среды (рН 7,4). Реакция
осуществляется без участия ферментов и сопровождается образованием пептидных связей.
В связанном с ферментом состоянии аминоациладенилаты инертны, и об их
использовании для синтеза белка прямо из состава комплекса не может быть речи.
1Реакции активирования сверхспецифичны. Активируются изомеры аминокислот
только природного Lряда; Dизомеры аминокислот в реакцию не вступают. Каталитическое
ускорение активирования каждой протеиногенной аминокислоты осуществляется
собственным, специфичным только для данной аминокислоты ферментом.
Активирующие ферменты называют аминоацилтРНКсинтетазамн (АРСазами).
Для нормальной деятельности активирующих аминокислоты ферментов необходимо
присутствие в реакционной среде Mg2+, так как связывание АТФ идет по имидазольному
радикалу гистидина активного центра фермента через ион магния.
Механизм действия. Первой к АРСазе присоединяется АТФ, аденилирующая остаток
гистидина в активном центре фермента с выделением пирофосфата:
С аденилированной АРСазой взаимодействует активируемая аминокислота, в
результате чего возникает аминоациладенилат. Последний,
обладая выдающейся
способностью аминоацилировать любые радикалы, содержащие подвижный атом водорода,
передает аминоацильную группу на радикал гистидина активного центра АРСазы, в
результате чего образуется аминоацилированный фермент:
После связывания ферментом соответствующей тРНК аминоацильная группа
передается на ОНгруппу остатка аденозина ее акцептирующего конца(ЦЦА).
Установлено также, что аминоацильная группа в аминоацилтРНК способна
перемещаться от 2' к 3'углеродному атому рибозы и наоборот, вплоть до установления
равновесия:
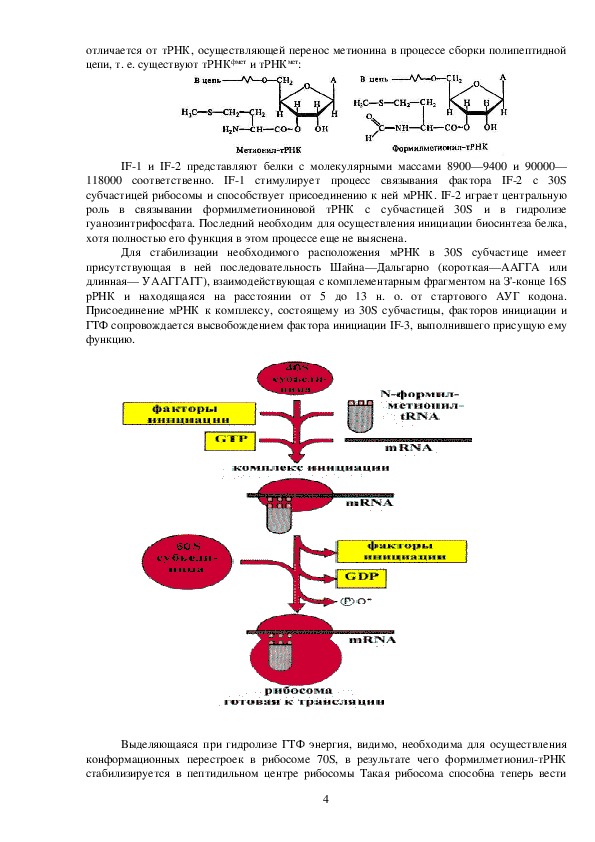
2Полагают, что аминоацилирование тРНК либо в 2', либо в 3'положении остатка
рибозы концевого аденозина предупреждает от ошибок при активировании стереохимически
близких аминокислот (например, валина и треонина).
Таким образом, в процессе активирования обеспечивается строго избирательное
присоединение каждой аминокислоты к специфичной для нее тРНК и создание набора
аминоацилтРНК,
непосредственно поставляющих энергетически обогащенные
аминокислотные остатки в рибосому.
Активирование аминокислот сопровождается их кодированием (шифрование): после
присоединения к соответствующей тРНК аминокислота получает код, или шифр, в виде
строго специфичного только для данной аминокислоты чередования трех нуклеотидных
остатков в антикодоновой петле тРНК. Этот триплет оснований называется антикодоном.
Ему соответствует комплементарный кодон в составе мРНК. Взаимодействие кодонов мРНК
с антикодонами аминоацилтРНК предопределяет порядок чередования аминокислотых
остатков в синтезируемом по матричной схеме белке.
Инициация биосинтеза белка у бактерий происходит при участии трех белковых
факторов инициации — IF1, IF2 и IF3 (Initiation Factors l, 2 и 3). IF1 представлен белком с
М=2100023500. Он вызывает конформационные изменения в 30S субчастице рибосомы,
способствующие связыванию ею IF1,
ГТФ, мРНК и формилметионилтРНК.
Присоединение последней обеспечивает поступление в рибосомальный аппарат клетки
первой, Nконцевой аминокислоты, а именно—формилметионина, которым открывается
полипептидная цепь любого белка, синтезируемого у бактерий. Впоследствии Nконцевой
остаток формилметионина в результате посттрансляционных изменений белка может
отщепляться. тРНК, переносящая формилметионин в процессе белкового синтеза у бактерий,
IF2,
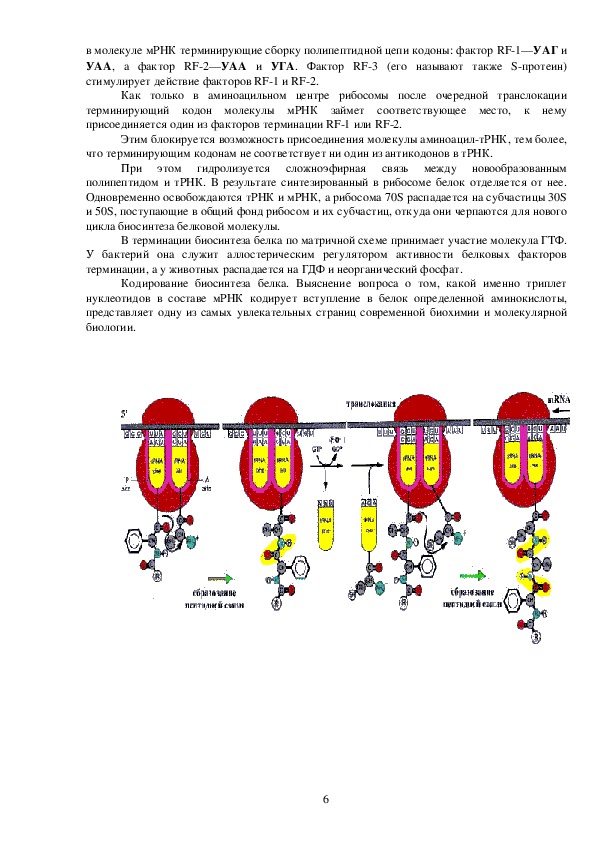
3отличается от тРНК, осуществляющей перенос метионина в процессе сборки полипептидной
цепи, т. е. существуют тРНКфмет и тРНКмет:
IF1 и IF2 представляют белки с молекулярными массами 8900—9400 и 90000—
118000 соответственно.
IF1 стимулирует процесс связывания фактора IF2 с 30S
субчастицей рибосомы и способствует присоединению к ней мРНК. IF2 играет центральную
роль в связывании формилметиониновой тРНК с субчастицей 30S и в гидролизе
гуанозинтрифосфата. Последний необходим для осуществления инициации биосинтеза белка,
хотя полностью его функция в этом процессе еще не выяснена.
Для стабилизации необходимого расположения мРНК в 30S субчастице имеет
присутствующая в ней последовательность Шайна—Дальгарно (короткая—ААГГА или
длинная— УААГГАГГ), взаимодействующая с комплементарным фрагментом на З'конце 16S
рРНК и находящаяся на расстоянии от 5 до 13 н. о. от стартового АУГ кодона.
Присоединение мРНК к комплексу, состоящему из 30S субчастицы, факторов инициации и
ГТФ сопровождается высвобождением фактора инициации IF3, выполнившего присущую ему
функцию.
Выделяющаяся при гидролизе ГТФ энергия, видимо, необходима для осуществления
конформационных перестроек в рибосоме 70S, в результате чего формилметионилтРНК
стабилизируется в пептидильном центре рибосомы Такая рибосома способна теперь вести
4сборку полипептидной цепи белка заданной структуры, вследствие чего ее называют
транслирующей (активной) рибосомой. В транслирующей рибосоме идет, следовательно,
процесс элонгации белкового синтеза.
Элонгация Элонгация биосинтеза белка в бактериальной клетке обслуживается тремя
белковыми факторами элонгации: EFTU, EFTS и EFG У млекопитающих два фактора
элонгации: TF1 и TF2
Процесс элонгации начинается со связывания аминоацилтРНК, содержащей
аминокислотный остаток, который должен быть вторым с Nконца молекулы синтезируемого
в рибосоме белка. У бактерий эта аминоацилтРНК образует комплекс с EFTU и ГТФ, в виде
которого она присоединяется к аминоацильному центру транслирующей рибосомы в
соответствии с кодом белкового синтеза т. е. благодаря взаимодействию комплементарных
триплетов антикодона тРНК и кодона мРНК, локализованного против аминоацильного
центра рибосомы. У млекопитающих это происходит при участии TF1.
По современным данным, не только кодонантикодоновое взаимодействие
предопределяет отбор соответствующих аминоацилтРНК для сборки полипептидной цепи. В
этом участвует вся молекула тРНК, так как посттрансляционная модификация сильно
изменяет ее способность акцептироваться рибосомой. В процесс декодирования вовлекаются
и белки рибосомы: S4, S9, S13, L2 и L7. Два последних, будучи локализованы в пептидильном
центре рибосомы, образуют с двумя молекулами тРНК тетрамерный комплекс.
Благодаря расщеплению ГТФ на ГДФ и неорганический фосфат аминоацилтРНК
сближается с формилметионилтРНК, локализованной в пептидильном центре рибосомы, a
EFTU в комплексе с ГДФ и неорганический фосфат выносятся из рибосомы. EFTU • ГДФ
комплекс при взаимодействии с EFTS и ГТФ преобразуется в EFT„ • ГТФкомплекс,
способный соединяться со следующей молекулой аминоацилтРНК.
В пептидильном центре между формилметионилтРНК и аминоацилтРНК происходит
реакция, благодаря которой остаток формилметионина переносится на свободную NH2
группy аминокислотного остатка, являющегося составной частью аминоацилтРНК. В
результате возникает дипептидилтРНК, т. е. замыкается первая пептидная связь в будущей
молекуле белка, а также образуется деацилированная тРНКфмет .
Этот процесс получил название реакции транспептидирования. Он ускоряется
соответствующим ферментом, причем транспептидазная активность присуща рибосомным
белкам L16, L11 и L6 и, вероятно, фрагменту 23S рРНК.
ПептидилтРНК на следующей фазе элонгации переносится на место тРНКфмет в
пептидильном центре рибосомы, а последняя удаляется из него, вытесняется очередной
аминоацилтРНК и выносится из рибосомы.
Эта ступень элонгации называется транслокацией и происходит при участии EFG у
бактерий и TF2 у эукариот, а также сопровождается непременным гидролизом еще одной
молекулы ГТФ.
В результате транслокации дипептидилтРНК занимает место в пептидильном центре
рибосомы, тогда как ее аминоацильный центр полностью освобождается и готов принять
новую аминоацилтРНК с TF1 и ГТФ.
Важно, что при транслокации пептидилтРНК перемещается в пептидильный центр
рибосомы вместе с молекулой мРНК, с которой она связана благодаря антикодон
кодоновому взаимодействию. Это перемещение идет точно на один триплет нуклеотидных
остатков, т. е. напротив аминоацильного центра оказывается следующий по порядку кодон
молекулы мРНК, предопределяющий, какая аминоацилтРНК вступит в аминоацильный центр
рибосомы и явится источником очередного аминокислотного остатка в новообразуемом
белке.
Терминация белкового синтеза в рибосоме осуществляется тоже при участии трех
белковых факторов: RF1, RF2 и RF3 у бактерий и единственного белкового фактораR — у
высших организмов (от англ. recognize—узнавать). Белковые факторы способны распознавать
5в молекуле мРНК терминирующие сборку полипептидной цепи кодоны: фактор RF1—УАГ и
УАА, а фактор RF2—УАА и УГА. Фактор RF3 (его называют также Sпротеин)
стимулирует действие факторов RF1 и RF2.
Как только в аминоацильном центре рибосомы после очередной транслокации
терминирующий кодон молекулы мРНК займет соответствующее место, к нему
присоединяется один из факторов терминации RF1 или RF2.
Этим блокируется возможность присоединения молекулы аминоацилтРНК, тем более,
что терминирующим кодонам не соответствует ни один из антикодонов в тРНК.
При этом гидролизуется сложноэфирная связь между новообразованным
полипептидом и тРНК. В результате синтезированный в рибосоме белок отделяется от нее.
Одновременно освобождаются тРНК и мРНК, а рибосома 70S распадается на субчастицы 30S
и 50S, поступающие в общий фонд рибосом и их субчастиц, откуда они черпаются для нового
цикла биосинтеза белковой молекулы.
В терминации биосинтеза белка по матричной схеме принимает участие молекула ГТФ.
У бактерий она служит аллостерическим регулятором активности белковых факторов
терминации, а у животных распадается на ГДФ и неорганический фосфат.
Кодирование биосинтеза белка. Выяснение вопроса о том, какой именно триплет
нуклеотидов в составе мРНК кодирует вступление в белок определенной аминокислоты,
представляет одну из самых увлекательных страниц современной биохимии и молекулярной
биологии.
67