1. Аэробный распад углеводов
2. Энергетический эффект распада углеводов.
3. Механизм синтеза углеводов. Синтез олиго и полисахаридов.
4. Регуляция обмена углеводов.
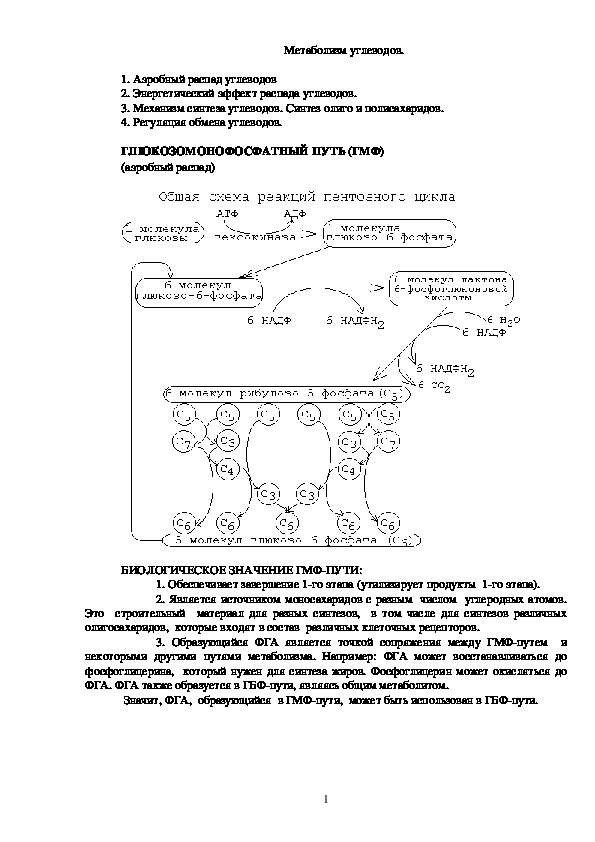
ГЛЮКОЗОМОНОФОСФАТНЫЙ ПУТЬ (ГМФ)
(аэробный распад)
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ГМФ-ПУТИ:
1. Обеспечивает завершение 1-го этапа (утилизирует продукты 1-го этапа).
2. Является источником моносахаридов с разным числом углеродных атомов. Это строительный материал для разных синтезов, в том числе для синтезов различных олигосахаридов, которые входят в состав различных клеточных рецепторов.
Метаболизм углеводов.
1. Аэробный распад углеводов
2. Энергетический эффект распада углеводов.
3. Механизм синтеза углеводов. Синтез олиго и полисахаридов.
4. Регуляция обмена углеводов.
ГЛЮКОЗОМОНОФОСФАТНЫЙ ПУТЬ (ГМФ)
(аэробный распад)
БИОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ГМФПУТИ:
1. Обеспечивает завершение 1го этапа (утилизирует продукты 1го этапа).
2. Является источником моносахаридов с разным числом углеродных атомов.
Это строительный материал для разных синтезов, в том числе для синтезов различных
олигосахаридов, которые входят в состав различных клеточных рецепторов.
3. Образующийся ФГА является точкой сопряжения между ГМФпутем и
некоторыми другими путями метаболизма. Например: ФГА может восстанавливаться до
фосфоглицерина, который нужен для синтеза жиров. Фосфоглицерин может окисляться до
ФГА. ФГА также образуется в ГБФпути, являясь общим метаболитом.
Значит, ФГА, образующийся в ГМФпути, может быть использован в ГБФпути.
1ГМФпуть заключается в том, что молекулы глюкозы превращаются в молекулы
глюкозо6фосфата, каждая из которых два раза окисляется и дает молекулу рибозо5
фосфата, 2 молекулы НАДФН2 и 1 молекулу СО2. Затем из каждых 6ти молекул
рибозо5фосфата получается 5 молекул глюкозо6фосфата. Эти 5 молекул глюкозо6
фосфата вместе с еще одной, поступающей дополнительно, опять вступают в ГМФпуть.
Получается цикл. На каждом его "обороте" происходит частичное окисление 6ти молекул
глюкозо6фосфата, то есть 5 молекул глюкозо6фосфата остаются нетронутыми, а одна
молекула полностью расщепляется до СО2 и Н2О.
Итоговое уравнение ГМФпути:
С6Н12О6 + 6 Н2О + 12 НАДФ > 6 CO2 + 12 НАДФН2
НАДФ поступает в дыхательную цепь В результате образуется на молекулу НАДФ 3
молекулы АТФ итого 36 молекул АТФ.
Энергетический эффект, за вычетом расхода 1 молекулы АТФ в реакции
фосфорилирования глюкозы (первая реакция ГМФ), составляет 35 молекул АТФ.
.
2Синтез углеводов.
ГЛЮКОНЕОГЕНЕЗ это образование углеводов (например, глюкозы или
гликогена из веществ, имеющих неуглеводное происхождение.
Некоторые промежуточные метаболиты ГБФпути могут образоваться из
веществ других классов (не из углеводов): например, из аминокислот, липидов.
Например:
Далее из пирувата синтезируется глюкоза (в печени) или гликоген (в печени и в
мышцах).
Для обходных реакций необратимых стадий 1го этапа ГБФпути существуют
специальные ферменты:
для 1й глюкозо6фосфатаза (только в печени!),
для 3й фруктозо1,6бисфосфатаза,
и для 10й реакции пируваткарбоксилаза.
Ключевым ферментом глюконеогенеза из пирувата является пируваткарбоксилаза. В
состав его кофермента входит витамин H биотин. Этот фермент обычно малоактивен, но он
сильно активируется даже при небольшом накоплении АцКоА в цитоплазме. Тогда обходной
обратный путь 10й стадии и весь процесс синтеза углеводов из ПВК может протекать
быстрее, чем их распад.
После образования глюкозо6фосфата начинается разветвление дальнейших
путей метаболизма глюкозы. Таких главных путей три.
Синтез и распад гликогена. Синтез гликогена протекает не во всех тканях, а только в
печени, мышцах и в лейкоцитах.
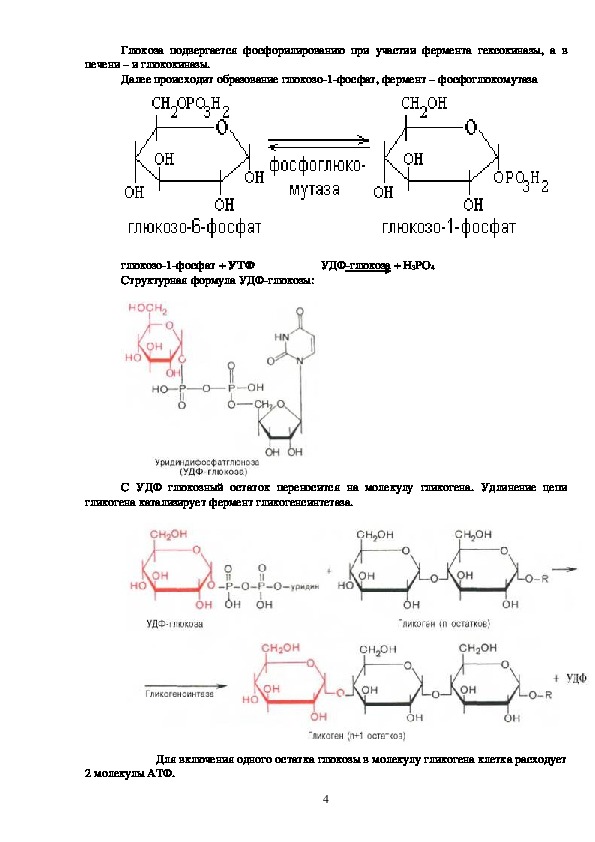
3Глюкоза подвергается фосфорилированию при участии фермента гексокиназы, а в
печени – и глюкокиназы.
Далее происходит образование глюкозо1фосфат, фермент – фосфоглюкомутаза
глюкозо1фосфат + УТФ УДФглюкоза + H3PO4
Структурная формула УДФглюкозы:
С УДФ глюкозный остаток переносится на молекулу гликогена. Удлинение цепи
гликогена катализирует фермент гликогенсинтетаза.
Для включения одного остатка глюкозы в молекулу гликогена клетка расходует
2 молекулы АТФ.
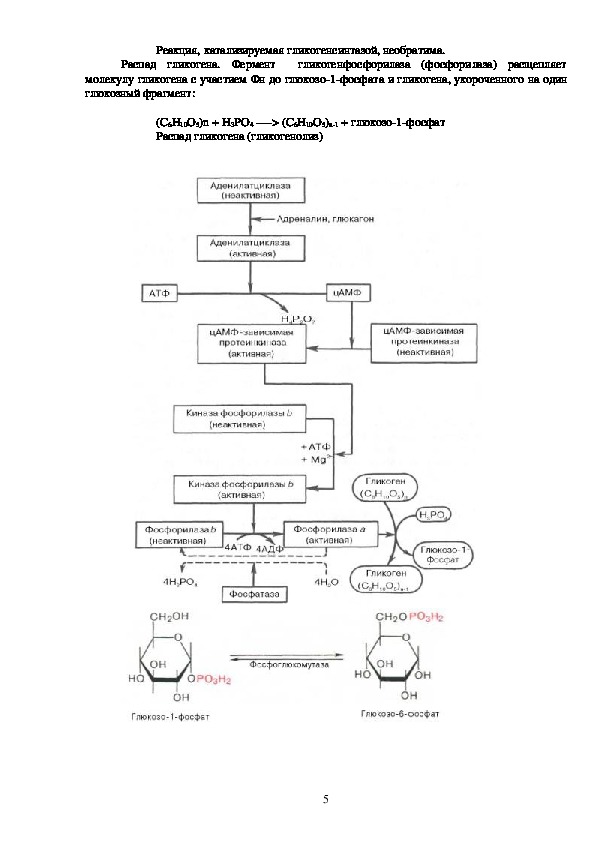
4Реакция, катализируемая гликогенсинтазой, необратима.
Распад гликогена. Фермент
гликогенфосфорилаза (фосфорилаза) расщепляет
молекулу гликогена с участием Фн до глюкозо1фосфата и гликогена, укороченного на один
глюкозный фрагмент:
(С6Н10О5)n + H3PO4 > (C6H10O5)n1 + глюкозо1фосфат
Распад гликогена (гликогенолиз)
5Сохранение постоянства концентрации глюкозы в крови является результатом
одновременного протекания двух процессов: поступления глюкозы в кровь из печени и
потребления ее из крови тканями, где она используется в первую очередь как энергетический
материал.
В тканях (в том числе в печени) распад глюкозы происходит двумя основными путями:
анаэробным (при отсутствии кислорода) и аэробным, для осуществления которого необходим
кислород.
Биосинтез олиго и полисахаридов. Синтез олигосахаридов. Долгое время
предпринимались безуспешные попытки доказать, что синтез олигосахаридов, в частности
дисахаридов, представляет собой реакцию обращения их гидролиза. Однако, несмотря на
многочисленные эксперименты такого рода, никто не сумел подобрать условия, при которых
удалось бы направить вспять реакцию гидролиза, например сахарозы, протекающую при
участии фруктофуранозидазы (сахараза).
β
Оказалось, что биосинтез олигосахаридов осуществляется путем реакций
трансгликозилирования. Перенос гликозильного остатка на один моносахарид идет с
фосфорного эфира другого моносахарида и ускоряется специфической
гликозилтрансферазой.
Исходными соединениями, с которых в процессе синтеза олигосахаридов
служат
гликозильный остаток энергично переносится на моносахарид,
нуклеозиддифосфатсахара (НДФсахара).
Они были открыты Л. Лелуаром с сотр. (1950) и очень быстро привлекли к себе
внимание как наиболее вероятные метаболиты в биосинтезе углеводов. В настоящее время
известно свыше 50 представителей НДФсахаров.
Значение: при гидролизе НДФсахаров изменение уровня свободной энергии
значительно выше, чем при гидролизе других доноров, а нуклеотидная часть их молекул
способна обеспечить избирательность гликозилтрансферазной реакции.
В случае синтеза сахарозы, например, специфический фермент—сахарозосинтаза,
ускоряет реакцию переноса остатка глюкозы с уридиндифосфатглюкозы на фруктозу.
Долгое время считали, что именно так синтезируется сахароза в растениях (животные
не синтезируют сахарозу, а лишь используют ее). Однако оказалось, что эта реакция ввиду ее
обратимости служит для поддержания равновесия между сахарозой и УДФглюкозой и даже
используется для наработки УДФглюкозы во время ее энергичного использования для
биосинтеза крахмала и гликогена.
Этот процесс идет в больших масштабах в клубнях растений, запасающих крахмал.
Сахароза же в действительности синтезируется при помощи гликозилтрансферазной реакции
из УДФглюкозы и фруктозо6фосфата:
6Сахарозофосфатсинтаза (молекулярная масса фермента из проростков пшеницы —
380000) действует в паре с фосфатазой, которая обеспечивает молниеносное отщепление
остатка фосфорной кислоты от сахарозо6фосфата, чем полностью сдвигает реакцию вправо.
Этот процесс идет в листьях растений, и образовавшаяся сахароза оттекает из них в клубни и
корни. Аналогично синтезируется главный дисахарид насекомых, высших и низших грибов и
микобактерий — трегалоза.
НДФсахара синтезируются из фосфорных эфиров моносахаридов и соответствующих
нуклеозидтрифосфатов путем нуклеотидилтрансферазных реакций, например:
Известно более двух десятков нуклеотидилтрансфераз, ускоряющих реакцию переноса
тех или иных нуклеотидных остатков на соответствующие фосфорные эфиры моносахаридов
с освобождением пирофосфата. Наряду с фосфорными эфирами моносахаридов и
нуклеозиддифосфатсахарами роль доноров гликозильных остатков в реакциях биосинтеза
олигосахаридов могут выполнять сами олигосахариды, а также декстрины.
Синтез полисахаридов. Подобно синтезу олигосахаридов
Таким образом биосинтез олигосахаридов идет путем переноса гликозильных
остатков на моносахариды с разнообразных субстратов при участии в каждом конкретном
случае соответствующих гликозилтрансфераз. При этом новообразование химических связей
сопряжено с распадом их в тех соединениях, с которых идет перенос гликозильных остатков.
новообразование
полисахаридов также идет путем трансгликозилирования. Полная аналогия существует и в
характере субстратов, с которых переносятся гликозильные остатки на конец растущей цепи
полисахарида. Ими могут быть фосфорные эфиры моноз, НДФсахара и олигосахариды.
Реакции переноса остатков моносахаридов в процессе биосинтеза полисахаридов ускоряются
соответствующими гликозилтрансферазами.
Например Синтез амилозы, целлюлозы и подобных им 1,4глюканов может
происходить путем переноса гликозильных остатков с глюкозо1фосфата или аналогичных
фосфорных эфиров моносахаридов. Эта реакция представляет собой обращение реакции
фосфоролиза указанных соединений. Однако более существенное значение имеет другой путь:
биосинтез из НДФсахаров при участии соответствующих трансгликозидаз.
Перенос гликозильного остатка идет на невосстанавливающий конец молекулы
синтезируемого полисахарида. Эта реакция может повторяться многократно, что
обеспечивает ступенчатый синтез молекул полисахаридов, содержащих огромное число
остатков моносахаридов.
Характерная особенность реакций такого типа состоит в необходимости затравки, т. е.
наличия в реакционной среде небольшого количества молекул полисахарида. Он как бы
предопределяет тот тип связи, который возникает в процессе трансгликозилирования, и,
следовательно, синтезируется полисахарид, одноименный с «затравочным».
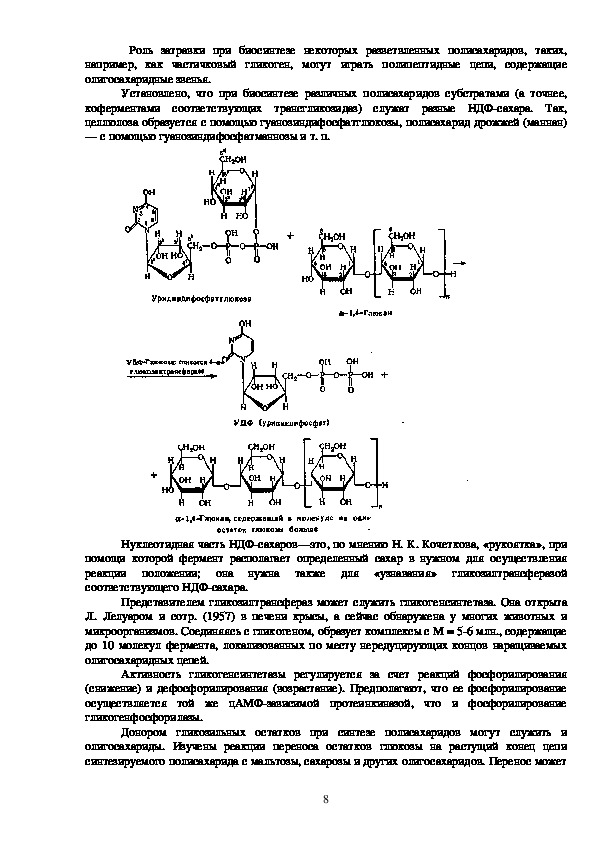
7Роль затравки при биосинтезе некоторых разветвленных полисахаридов, таких,
например, как частичковый гликоген, могут играть полипептидные цепи, содержащие
олигосахаридные звенья.
Установлено, что при биосинтезе различных полисахаридов субстратами (а точнее,
коферментами соответствующих трансгликозидаз) служат разные НДФсахара.
Так,
целлюлоза образуется с помощью гуанозиндифосфатглюкозы, полисахарид дрожжей (маннан)
— с помощью гуанозиндифосфатманнозы и т. п.
Нуклеотидная часть НДФсахаров—это, по мнению Н. К. Кочеткова, «рукоятка», при
помощи которой фермент располагает определенный сахар в нужном для осуществления
реакции положении; она нужна также для «узнавания» гликозилтрансферазой
соответствующего НДФсахара.
Представителем гликозилтрансфераз может служить гликогенсинтетаза. Она открыта
Л. Лелуаром и сотр. (1957) в печени крысы, а сейчас обнаружена у многих животных и
микроорганизмов. Соединяясь с гликогеном, образует комплексы с М = 56 млн., содержащие
до 10 молекул фермента, локализованных по месту нередуцирующих концов наращиваемых
олигосахаридных цепей.
Активность гликогенсинтетазы регулируется за счет реакций фосфорилирования
(снижение) и дефосфорилирования (возрастание). Предполагают, что ее фосфорилирование
осуществляется той же цАМФзависимой протеинкиназой, что и фосфорилирование
гликогенфосфорилазы.
Донором гликозильных остатков при синтезе полисахаридов могут служить и
олигосахариды. Изучены реакции переноса остатков глюкозы на растущий конец цепи
синтезируемого полисахарида с мальтозы, сахарозы и других олигосахаридов. Перенос может
8идти не только на 4й, но и на 6й углеродный атом остатка моносахарида, что
обеспечивает синтез 1,6глюканов, а возможно, и полисахаридов разветвленного строения.
Для реакций синтеза полисахаридов характерно также, что осуществляется перенос не
только остатков моносахаридов, но и полигликозидных фрагментов с одного полисахарида на
другой или же в пределах одной и той же молекулы. Реакции этого типа изучены у нас А. Н.
Петровой и ею же впервые получен из скелетных мышц кролика гомогенный препарат
соответствующего фермента (1970).
Использование промежуточных продуктов распада углеводов для синтеза других
органических соединений. Выше было отмечено, что одна из функций углеводов в обмене
веществ состоит в образовании продуктов распада, которые служат исходными веществами
для синтеза многих других молекул. Из числа продуктов распада углеводов в этом смысле
важны фосфоглицериновая кислота, фосфоенолпировиноградная кислота, пировиноградная
кислота, ацетилкоэнзим А, эритрозо4фосфат, рибулозо5фосфат, а также партнеры цикла
трикарбоновых и дикарбоновых кислот: щавелевоуксусная и оскетоглутаровая кислоты. Они
служат исходными соединениям для синтеза аминокислот, высших жирных кислот, глицерина,
нуклеотидов и ряда других мономеров, используемых для построения белков, липидов,
нуклеиновых кислот и других биополимеров.
Регуляция обмена углеводов. Эффект Пастера
В состоянии покоя, наступающего после интенсивной мышечной работы, в клетку
начинает поступать кислород. Это приводит к запуску митохондриальных дыхательных цепей.
Запускаются окислительное декарбоксилирование пирувата, ЦТК и челночный механизм
переноса водорода в митохондрии, а, значит, и ГБФпуть распада глюкозы (гликогена). При
этом процесс гликолиза тормозится автоматически. Торможение гликолиза поступившим в
клетку кислородом называется ЭФФЕКТОМ ПАСТЕРА по имени ученого, открывшего это
явление.
В раковых клетках такого эффекта не наблюдается. В них одновременно могут
протекать сразу два процесса: и ГБФпуть, и гликолиз. Отсутствие эффекта Пастера в тканях,
пораженных злокачественными опухолями, называется ЭФФЕКТОМ КРЭБТРИ.
Пути регуляции метаболизма углеводов разнообразны.
На любых уровнях организации живого организма обмен углеводов регулируется
факторами, влияющими на активность ферментов, участвующих в реакциях углеводного
обмена.
К этим факторам относятся
концентрация субстратов,
содержание продуктов (метаболитов) отдельных реакций,
кислородный режим,
температура,
проницаемость биологических мембран,
концентрация коферментов, необходимых для отдельных реакций.
Гликолиз – это совокупность реакций превращения глюкозы в пируват. Десять реакций
гликолиза протекают в цитозоле.
Гликолитический путь играет двоякую роль: приводит к генерированию АТФ в
результате распада глюкозы, и он же поставляет строительные блоки для синтеза клеточных
компонентов. Реакции гликолитического пути в физиологических условиях легкообратимы,
кроме реакций, катализируемых гексокиназой, фосфофруктокиназой и пируваткиназой.
Фосфофруктокиназа – наиболее важный регуляторный элемент (фермент) в процессе
гликолиза, ингибируется высокими концентрациями АТФ и цитрата и активируется АМФ.
Цикл трикарбоновых кислот (цикл Кребса) представляет собой конечный общий путь
для окисления «топливных» молекул. Большинство «топливных» молекул вступает в цикл в
виде ацетилКоА. Окислительное декарбоксилирование пирувата, приводящее к образованию
9ацетилКоА, является связующим звеном между гликолизом и циклом трикарбоновых кислот.
Все реакции цикла протекают в митохондриях.
Скорость цикла трикарбоновых кислот зависит от потребности в АТФ. Высокий
энергетический заряд клетки понижает активность цитратсинтазы, изоцитратдегидрогеназы и
α
кетоглутаратдегидрогеназы.
У человека и животных на всех стадиях синтеза и распада углеводов регуляция
углеводного обмена осуществляется при участии ЦНС и гормонов.
Например, установлено, что концентрация глюкозы в крови ниже 3,3–3,4 ммоль/л (60–
70 мг/100 мл) приводит к рефлекторному возбуждению высших метаболических центров,
расположенных в гипоталамусе. В регуляции углеводного обмена особая роль принадлежит
высшему отделу ЦНС – коре большого мозга. Наряду с ЦНС важное влияние на содержание
глюкозы оказывают гормональные факторы, т.е. регуляции уровня глюкозы в крови
осуществляется ЦНС через ряд эндокринных желез.
10