Поделиться

Половое размножение растений
Жизненный цикл растения включает два поколения: диплоидного спорофита (диплобионта) и гаплоидного гаметофита (гаплобионта).
В процессе эволюционного развития спорофит становился все крупнее и самостоятельнее, а гаметофит постепенно редуцировался (уменьшился в размерах).
Цветковые растения имеют микроскопический гаметофит, входящий в состав цветка (рис. 1)
· женский гаметофит — это зародышевый мешок в семязачатке завязи пестика;
· мужской гаметофит — пыльцевое зерно в пыльнике тычинки.

Рис. 1. Микроскопические гаметофиты
Половое размножение растений происходит с участием гамет.
Тычинки являются мужскими органами цветка, и в них происходит образование и развитие мужского гаметофита. Так же, как и у голосеменных он представлен пыльцевым зерном.
Образование пыльцевых зёрен происходит в камерах пыльников из диплоидных стволовых клеток
(рис. 2, А).
Стволовая клетка (2n) делится мейозом, образуя 4 гаплоидные микроспоры (n).
Ядро каждой микроспоры делится митозом, образуя два гаплоидных ядра: вегетативное (n) и генеративное (n).
Вокруг генеративного ядра концентрируется часть цитоплазмы, и формируется генеративная клетка (внутри вегетативной клетки).
На поверхности цитоплазматической мембраны микроспоры из содержимого пыльцевого мешка образуется очень прочная оболочка, нерастворимая в кислотах и щелочах.
Таким образом, каждое пыльцевое зерно состоит из вегетативной и генеративной клеток и покрыто двумя оболочками.
В дальнейшем из генеративной клетки путем митоза образуются 2 спермия. Этот процесс может происходить позже, параллельно с прорастанием пыльцевого зерна при оплодотворении.
Множество пыльцевых зёрен составляет пыльцу растения. Пыльца созревает в пыльниках к моменту распускания цветка.
Развитие пыльцы приводит к разрастанию пыльцевых камер, их стенки разрываются и пыльца высыпается наружу.
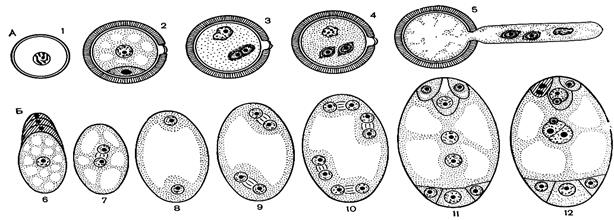
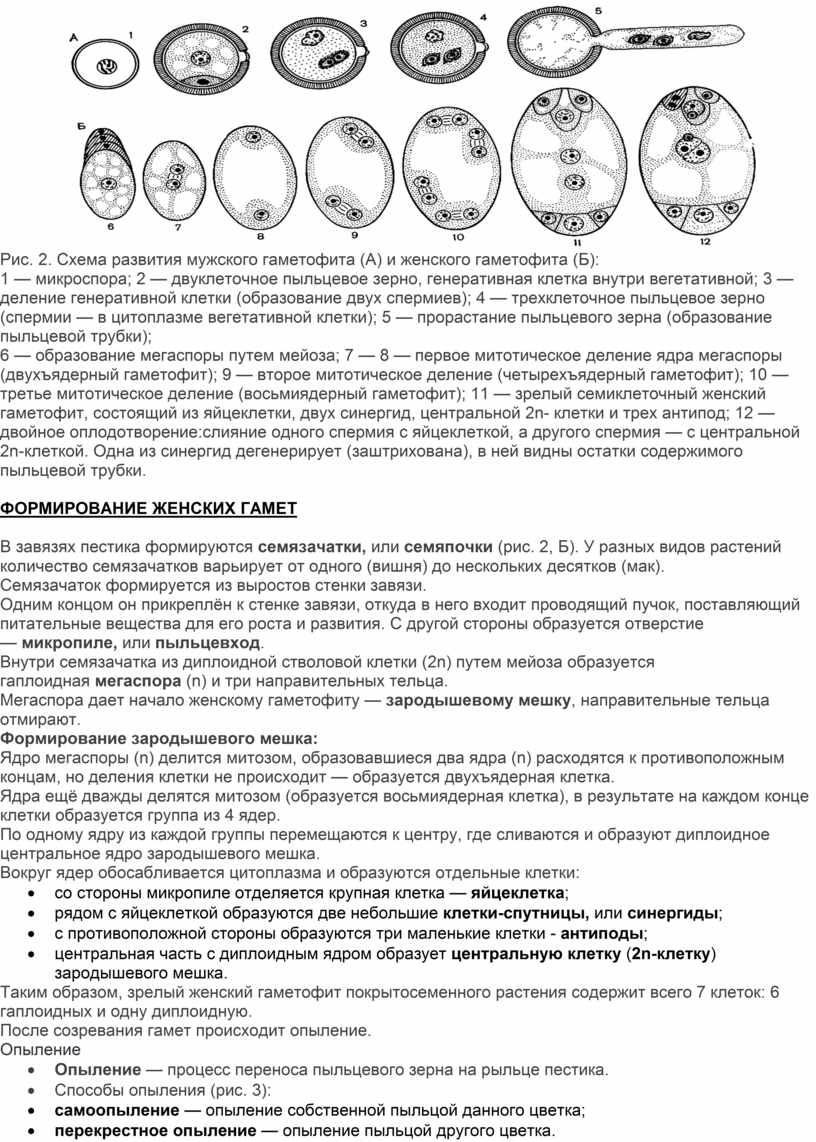
Рис. 2. Схема развития мужского гаметофита (А) и женского гаметофита (Б):
1 — микроспора; 2 — двуклеточное пыльцевое зерно, генеративная клетка внутри вегетативной; 3 — деление генеративной клетки (образование двух спермиев); 4 — трехклеточное пыльцевое зерно (спермии — в цитоплазме вегетативной клетки); 5 — прорастание пыльцевого зерна (образование пыльцевой трубки);
6 — образование мегаспоры путем мейоза; 7 — 8 — первое митотическое деление ядра мегаспоры (двухъядерный гаметофит); 9 — второе митотическое деление (четырехъядерный гаметофит); 10 — третье митотическое деление (восьмиядерный гаметофит); 11 — зрелый семиклеточный женский гаметофит, состоящий из яйцеклетки, двух синергид, центральной 2n- клетки и трех антипод; 12 — двойное оплодотворение:слияние одного спермия с яйцеклеткой, а другого спермия — с центральной 2n-клеткой. Одна из синергид дегенерирует (заштрихована), в ней видны остатки содержимого пыльцевой трубки.
В завязях пестика формируются семязачатки, или семяпочки (рис. 2, Б). У разных видов растений количество семязачатков варьирует от одного (вишня) до нескольких десятков (мак).
Семязачаток формируется из выростов стенки завязи.
Одним концом он прикреплён к стенке завязи, откуда в него входит проводящий пучок, поставляющий питательные вещества для его роста и развития. С другой стороны образуется отверстие — микропиле, или пыльцевход.
Внутри семязачатка из диплоидной стволовой клетки (2n) путем мейоза образуется гаплоидная мегаспора (n) и три направительных тельца.
Мегаспора дает начало женскому гаметофиту — зародышевому мешку, направительные тельца отмирают.
Формирование зародышевого мешка:
Ядро мегаспоры (n) делится митозом, образовавшиеся два ядра (n) расходятся к противоположным концам, но деления клетки не происходит — образуется двухъядерная клетка.
Ядра ещё дважды делятся митозом (образуется восьмиядерная клетка), в результате на каждом конце клетки образуется группа из 4 ядер.
По одному ядру из каждой группы перемещаются к центру, где сливаются и образуют диплоидное центральное ядро зародышевого мешка.
Вокруг ядер обосабливается цитоплазма и образуются отдельные клетки:
Таким образом, зрелый женский гаметофит покрытосеменного растения содержит всего 7 клеток: 6 гаплоидных и одну диплоидную.
После созревания гамет происходит опыление.
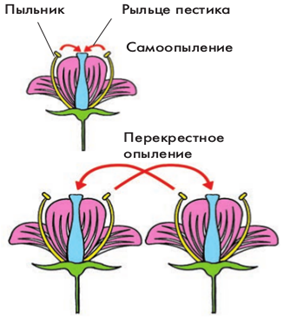
Рис. 3. Способы опыления
Существует несколько механизмов переноса пыльцы с одного растения на другое:
· с помощью ветра опыляются анемофильные растения (рис. 4);
· с помощью насекомых опыляются энтомофильные растения (рис. 6);
· с помощью животных (например, колибри и летучих мышей) опыляются многочисленные тропические зоофильные растения (рис. 7, 8);
· с помощью воды опыляются некоторые водные растения (рис. 5).



Рис. 4. Рожь Рис. 5. Роголистник Рис. 6. Короставник


Рис. 7. Опыление летучими мышами Рис. 8. Опыление колибри
Пыльца большинства растений в воде лопается из-за высокого осмотического давления, поэтому даже если растение растёт под водой, его цветы в большинстве случаев поднимаются над водой и опыляются насекомыми.
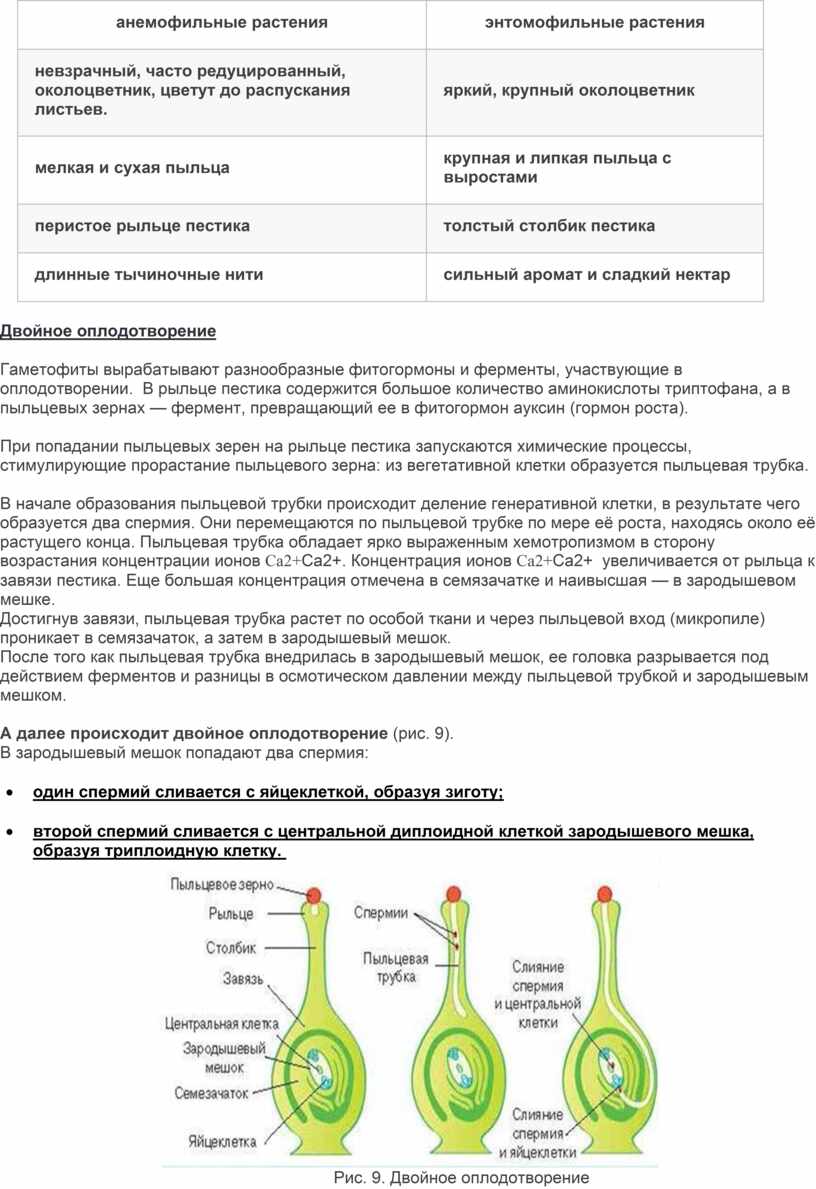
|
анемофильные растения |
энтомофильные растения |
|
невзрачный, часто редуцированный, околоцветник, цветут до распускания листьев. |
яркий, крупный околоцветник |
|
мелкая и сухая пыльца |
крупная и липкая пыльца с выростами |
|
перистое рыльце пестика |
толстый столбик пестика |
|
длинные тычиночные нити |
сильный аромат и сладкий нектар |
Гаметофиты вырабатывают разнообразные фитогормоны и ферменты, участвующие в оплодотворении. В рыльце пестика содержится большое количество аминокислоты триптофана, а в пыльцевых зернах — фермент, превращающий ее в фитогормон ауксин (гормон роста).
При попадании пыльцевых зерен на рыльце пестика запускаются химические процессы, стимулирующие прорастание пыльцевого зерна: из вегетативной клетки образуется пыльцевая трубка.
В начале образования пыльцевой трубки происходит деление генеративной клетки, в результате чего образуется два спермия. Они перемещаются по пыльцевой трубке по мере её роста, находясь около её растущего конца. Пыльцевая трубка обладает ярко выраженным хемотропизмом в сторону возрастания концентрации ионов Ca2+Ca2+. Концентрация ионов Ca2+Ca2+ увеличивается от рыльца к завязи пестика. Еще большая концентрация отмечена в семязачатке и наивысшая — в зародышевом мешке.
Достигнув завязи, пыльцевая трубка растет по особой ткани и через пыльцевой вход (микропиле) проникает в семязачаток, а затем в зародышевый мешок.
После того как пыльцевая трубка внедрилась в зародышевый мешок, ее головка разрывается под действием ферментов и разницы в осмотическом давлении между пыльцевой трубкой и зародышевым мешком.
А далее происходит двойное оплодотворение (рис. 9).
В зародышевый мешок попадают два спермия:
· один спермий сливается с яйцеклеткой, образуя зиготу;
· второй спермий сливается с центральной диплоидной клеткой зародышевого мешка, образуя триплоидную клетку.
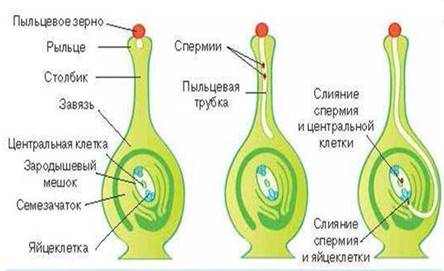
Рис. 9. Двойное оплодотворение
Клетки-спутницы и антиподы отмирают, а зигота, центральная клетка и клетки стенки семязачатка участвуют в образовании семени. Зигота митотически делится и превращается в многоклеточный зародыш, а триплоидная клетка, митотически делясь, превращается в триплоидное многоклеточное образование — эндосперм, питательную ткань для зародыша. В паренхиме эндосперма накапливаются крахмал, белки и жиры, для гетеротрофного питания зародыша.
Между опылением и оплодотворением проходит определенный промежуток времени: у некоторых растений 15 мин, у других — часы, недели и даже месяцы.
· Возникновение в процессе эволюции механизма двойного оплодотворения имело большое значение:
Процесс двойного оплодотворения у покрытосеменных растений
открыл в 1898 году
русский ботаник академик Сергей Гаврилович Навашин (рис. 10)

Рис. 10.
Академик Сергей Гаврилович Навашин



Материалы на данной страницы взяты из открытых источников либо размещены пользователем в соответствии с договором-офертой сайта. Вы можете сообщить о нарушении.
