Поделиться

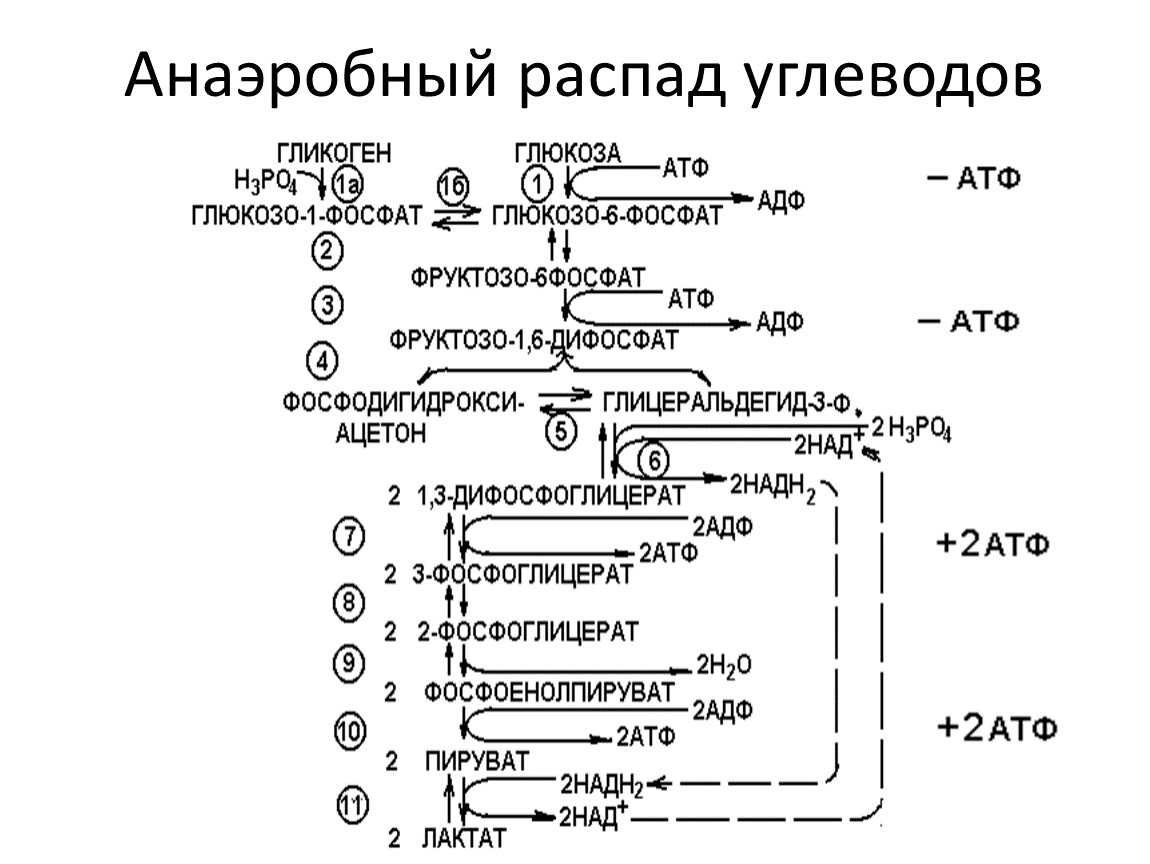
Анаэробный распад углеводов

Ферменты анаэробных гликолиза и гликогенолиза
1.Гексокиназа (глюкокиназа) (Mg2+) (2.7.1.1.)
2.Глюкозофосфатизомераза (5.3.1.9.)
3.Фосфофруктокиназа (Mg2+) (2.7.1.11.)
4.Фруктозо-1,6-дифосфатальдолаза (4.1.2.13.)
5.Триозофосфатизомераза (5.3.1.1.)
6.Глицеральдегидфосфатдегидрогеназа (1.2.1.12.)
7.Фосфоглицераткиназа (2.7.2.3.)
8.Фосфоглицеромутаза (Mg2+) (5.4.2.1.)
9.Енолаза (Mg2+; Mn2+) (4.2.1.11.)
10.Пируваткиназа (Mg2+; Mn2+; К+ или Na+) (2.7.1.40.)
11. Лактатдегидрогеназа (1.1.1.27.)
1а. Фосфорилаза (2.4.1.1.) и амило-1,6-глюкозидаза
1б.Фосфоглюкомутаза (2.7.5.1.)

Молочнокислое брожение
Известны две группы молочнокислых бактерий. Одни из них в процессе брожения углеводов образуют только молочную кислоту, другие из каждой молекулы глюкозы “производят” по одной молекуле молочной кислоты, этанола и СО2.
Стадии молочнокислого брожения, катализируемые ферментами бактерий первого вида, аналогичны стадиям анаэробного гликолиза; второго типа - смесь реакций анаэробного гликолиза и спиртового брожения.
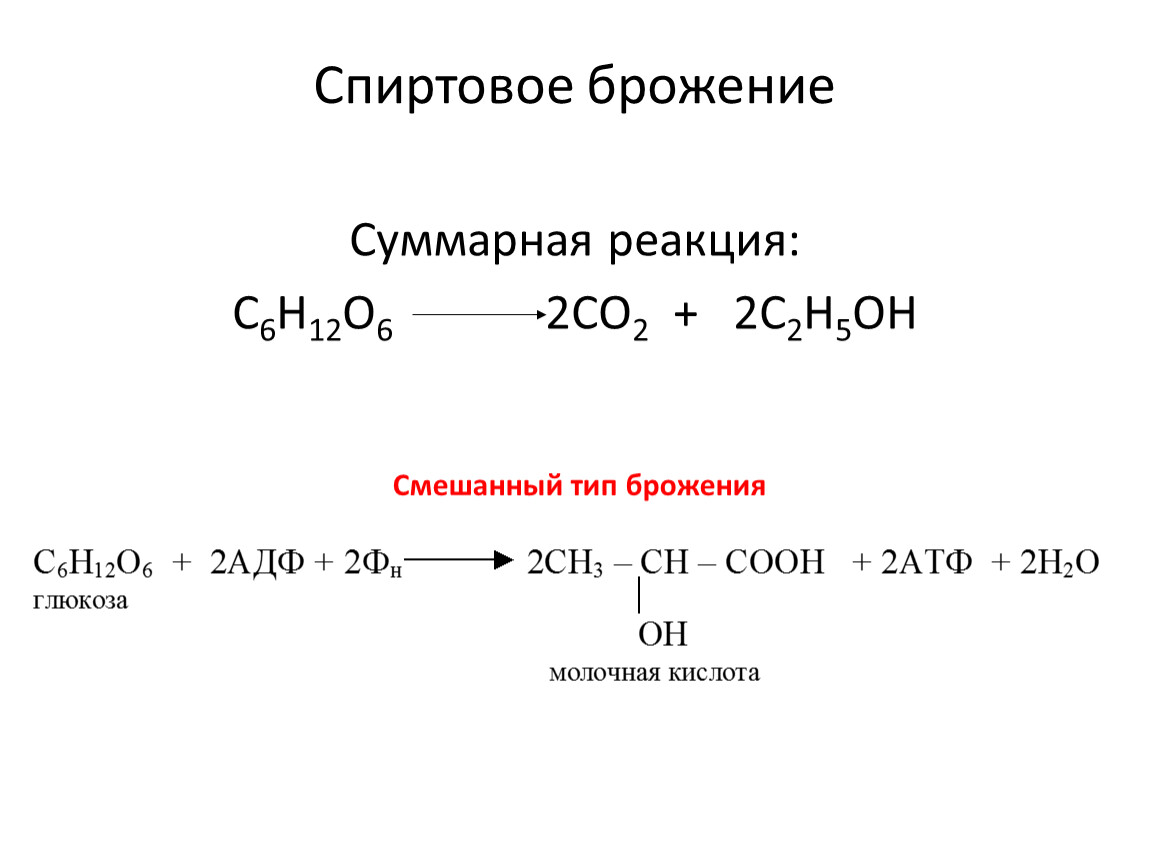
Спиртовое брожение
Суммарная реакция:
С6Н12О6 2СО2 + 2С2Н5ОН
Смешанный тип брожения
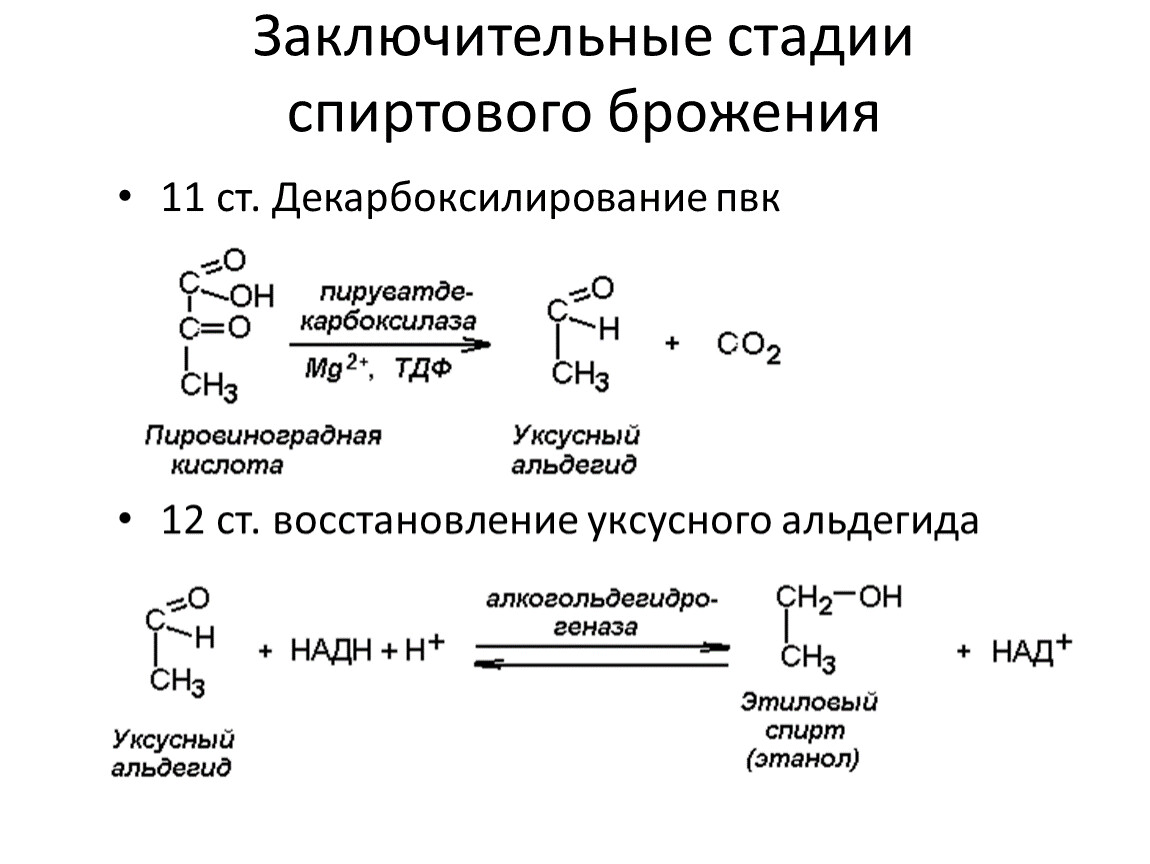
Заключительные стадии спиртового брожения
11 ст. Декарбоксилирование пвк
12 ст. восстановление уксусного альдегида
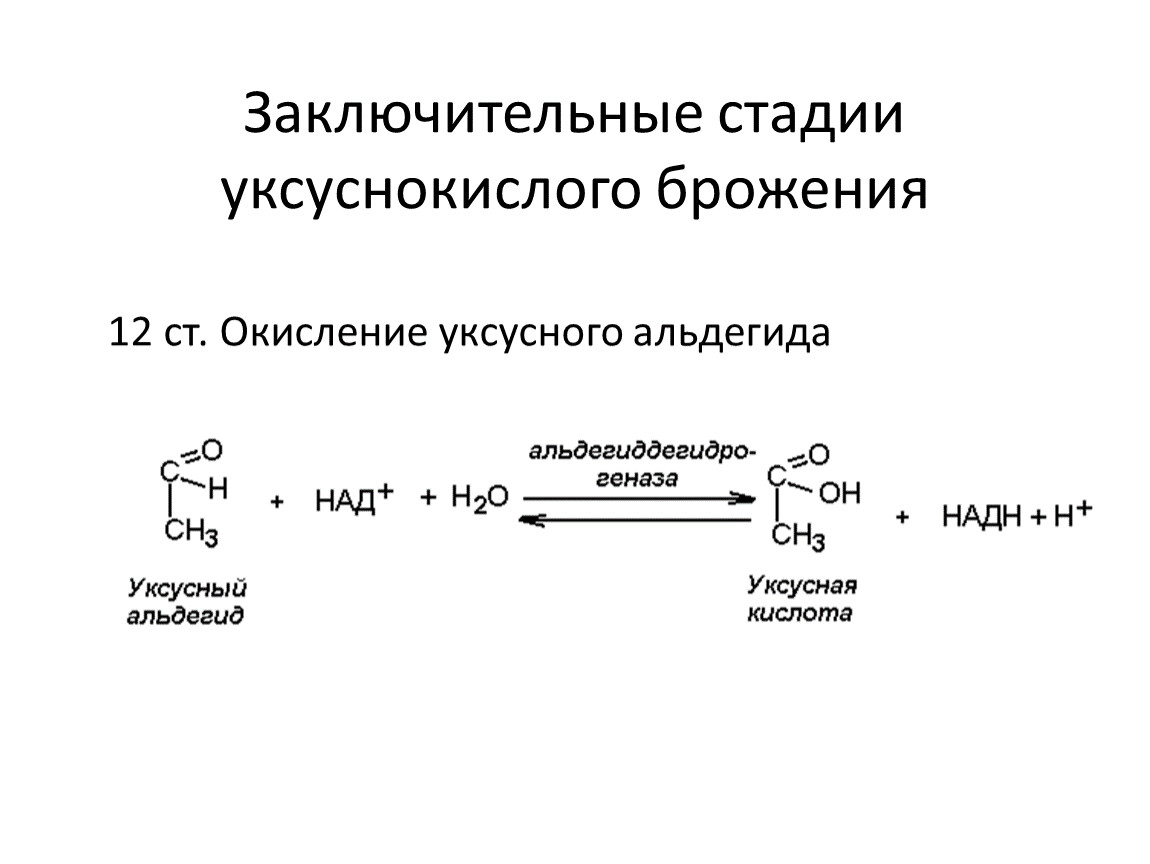
Заключительные стадии уксуснокислого брожения
12 ст. Окисление уксусного альдегида

В аэробных условиях (при наличии кислорода) распад глюкозы будет проходить в 3 этапа:
I этап: распад глюкозы не до молочной кислоты, а до пирувата.
II этап: окислительное декарбоксилирование пировиноградной кислоты.
III этап: цитратный цикл (общий путь катаболизма).
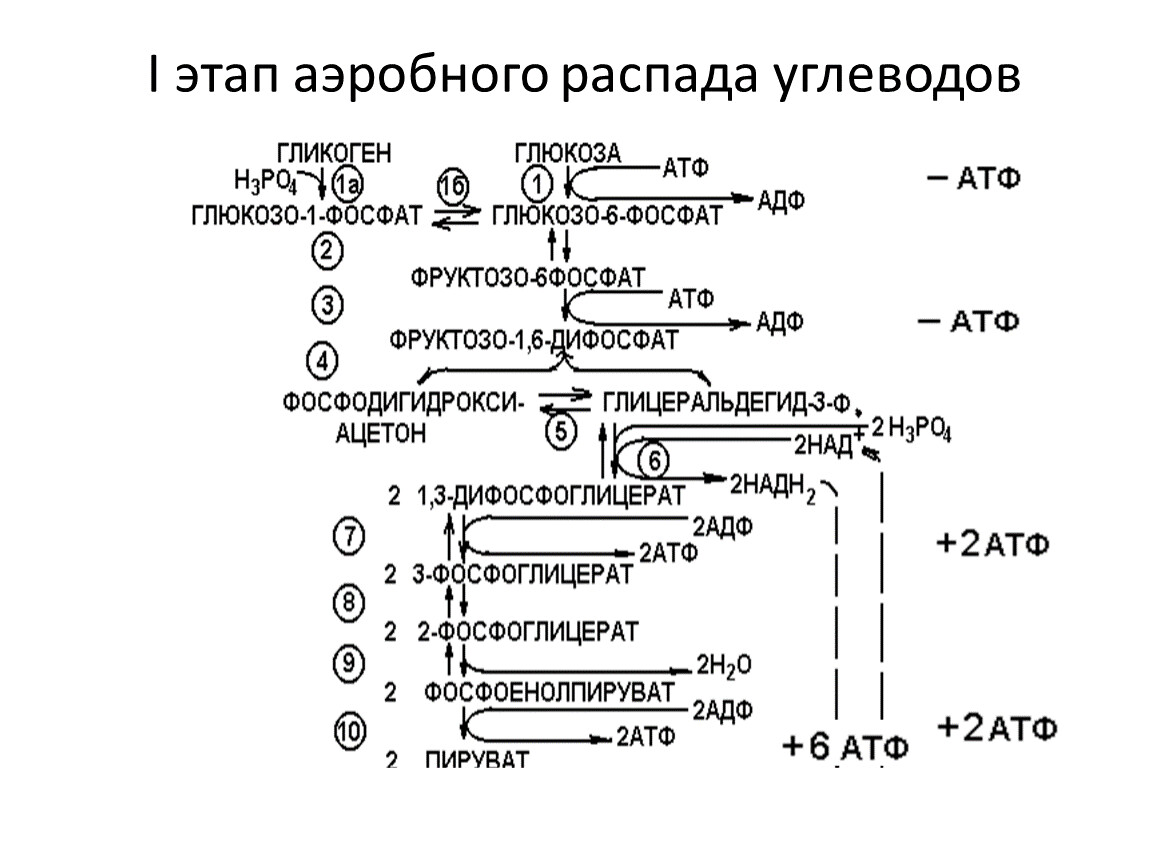
I этап аэробного распада углеводов
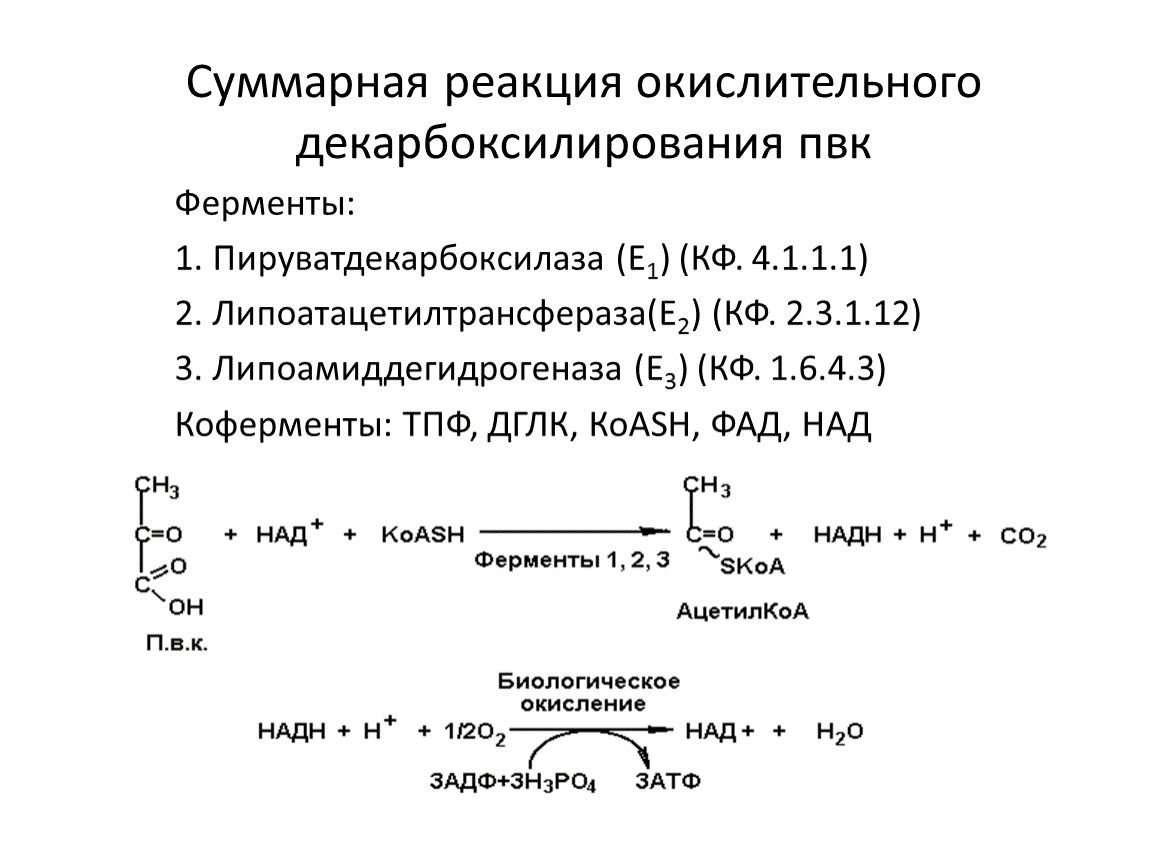
Суммарная реакция окислительного декарбоксилирования пвк
Ферменты:
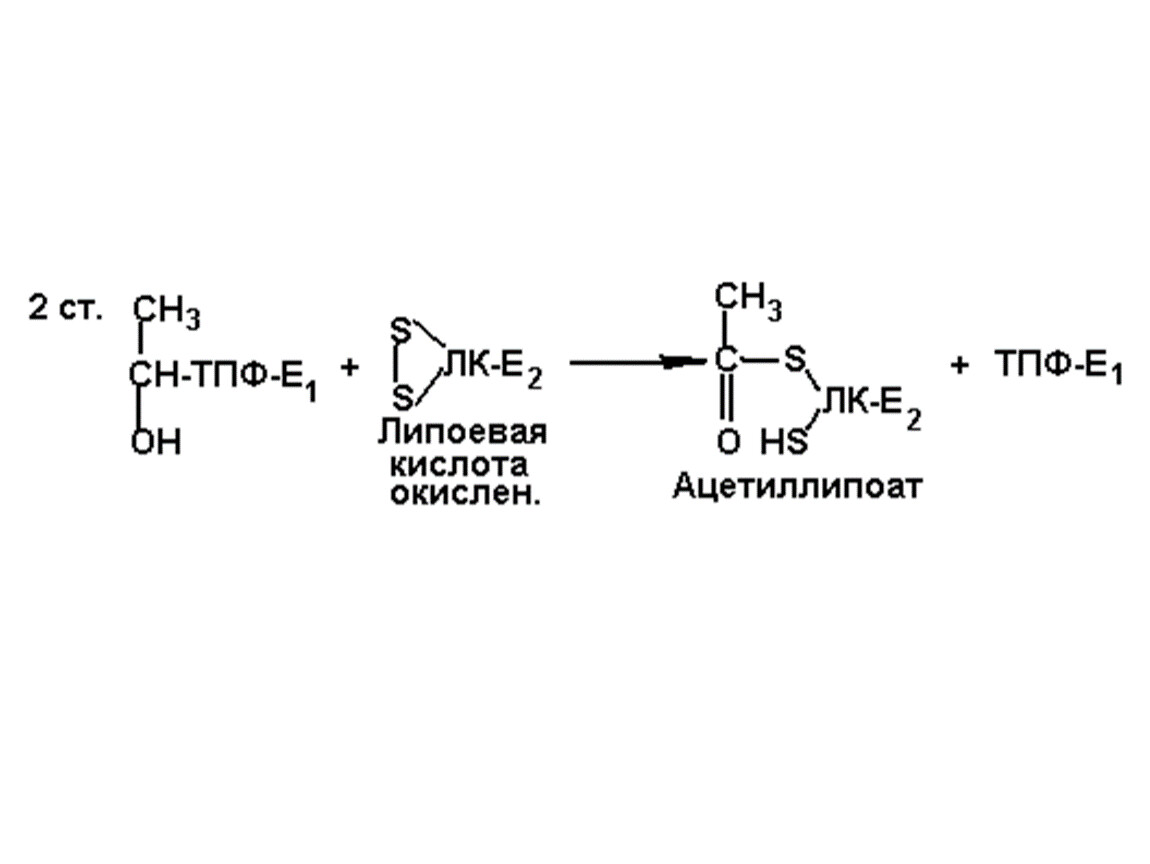
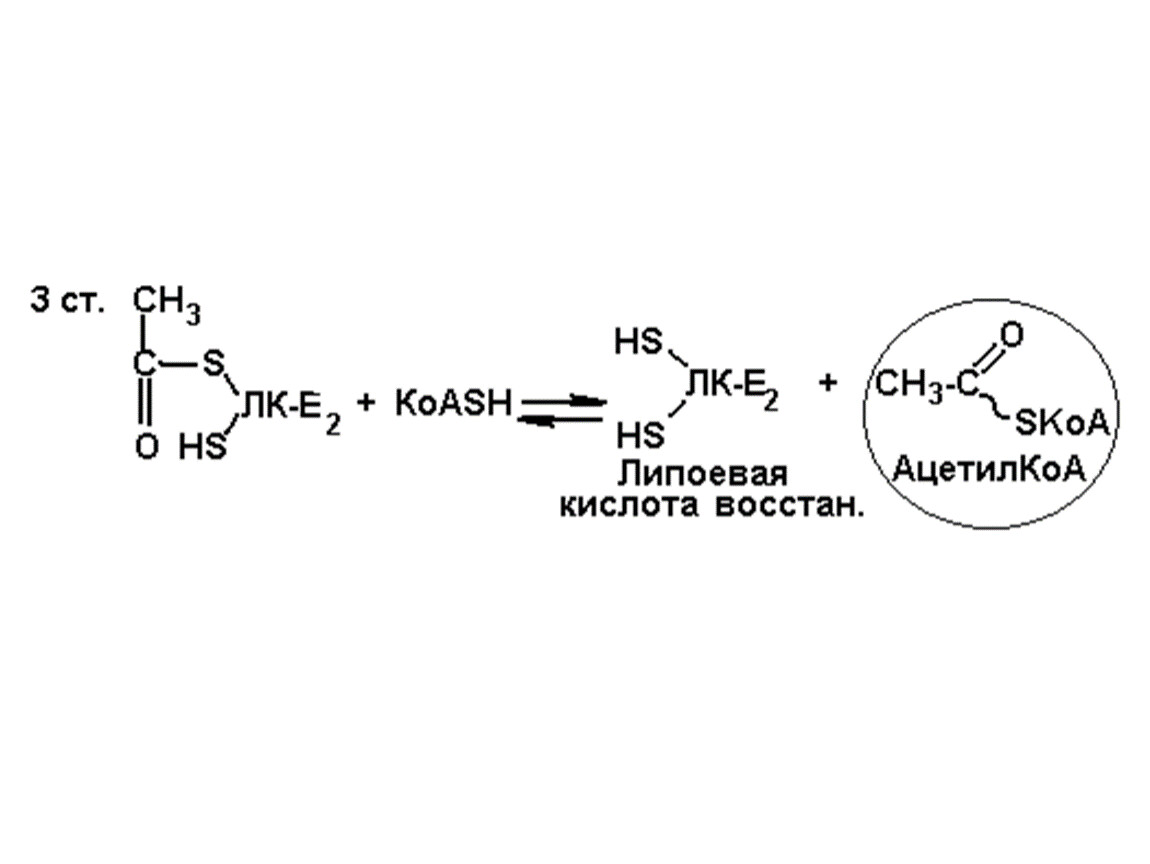
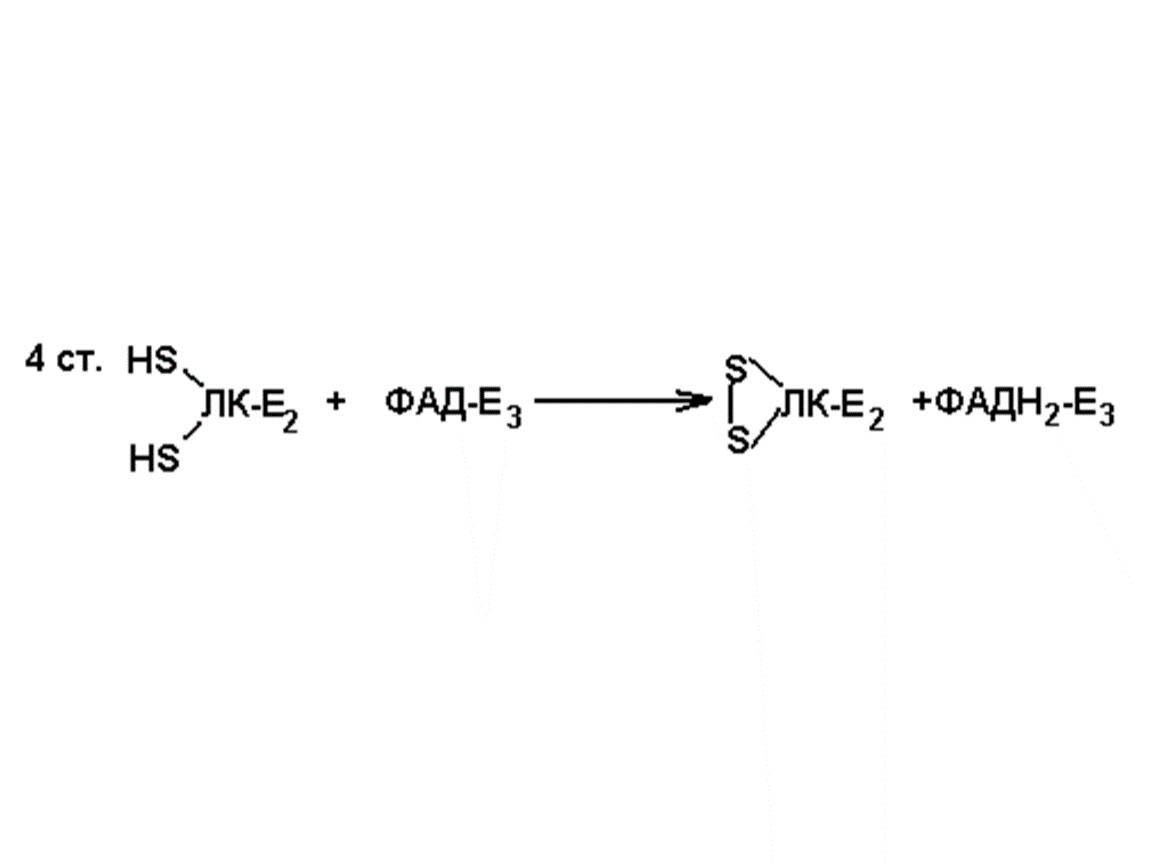
1. Пируватдекарбоксилаза (Е1) (КФ. 4.1.1.1)
2. Липоатацетилтрансфераза(Е2) (КФ. 2.3.1.12)
3. Липоамиддегидрогеназа (Е3) (КФ. 1.6.4.3)
Коферменты: ТПФ, ДГЛК, КоАSН, ФАД, НАД

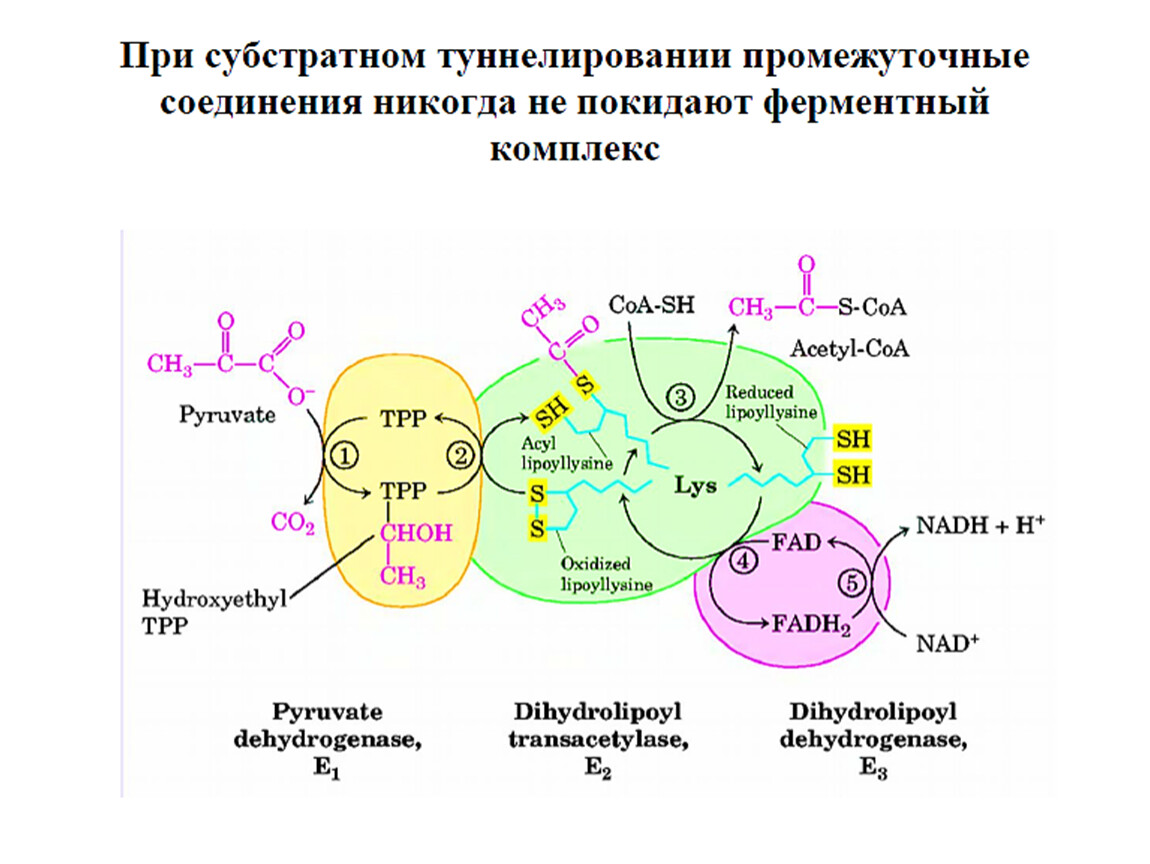


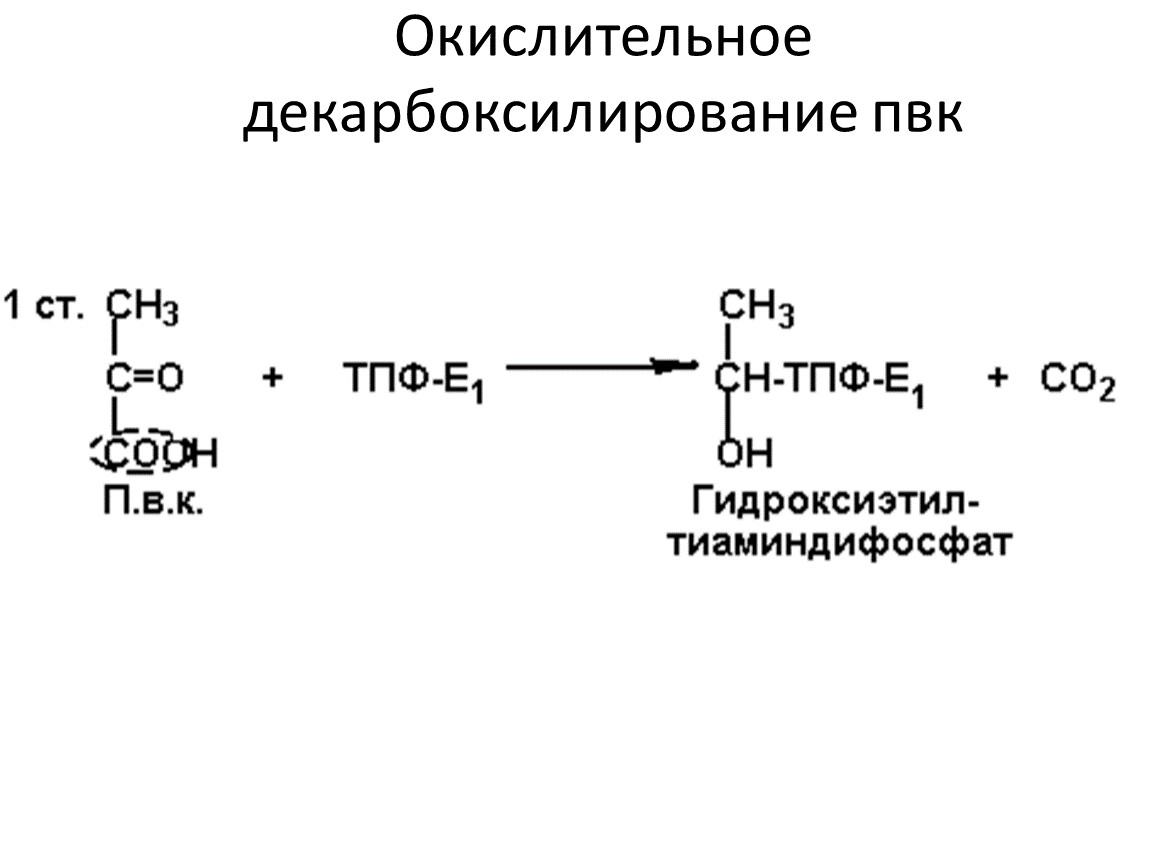
Окислительное декарбоксилирование пвк
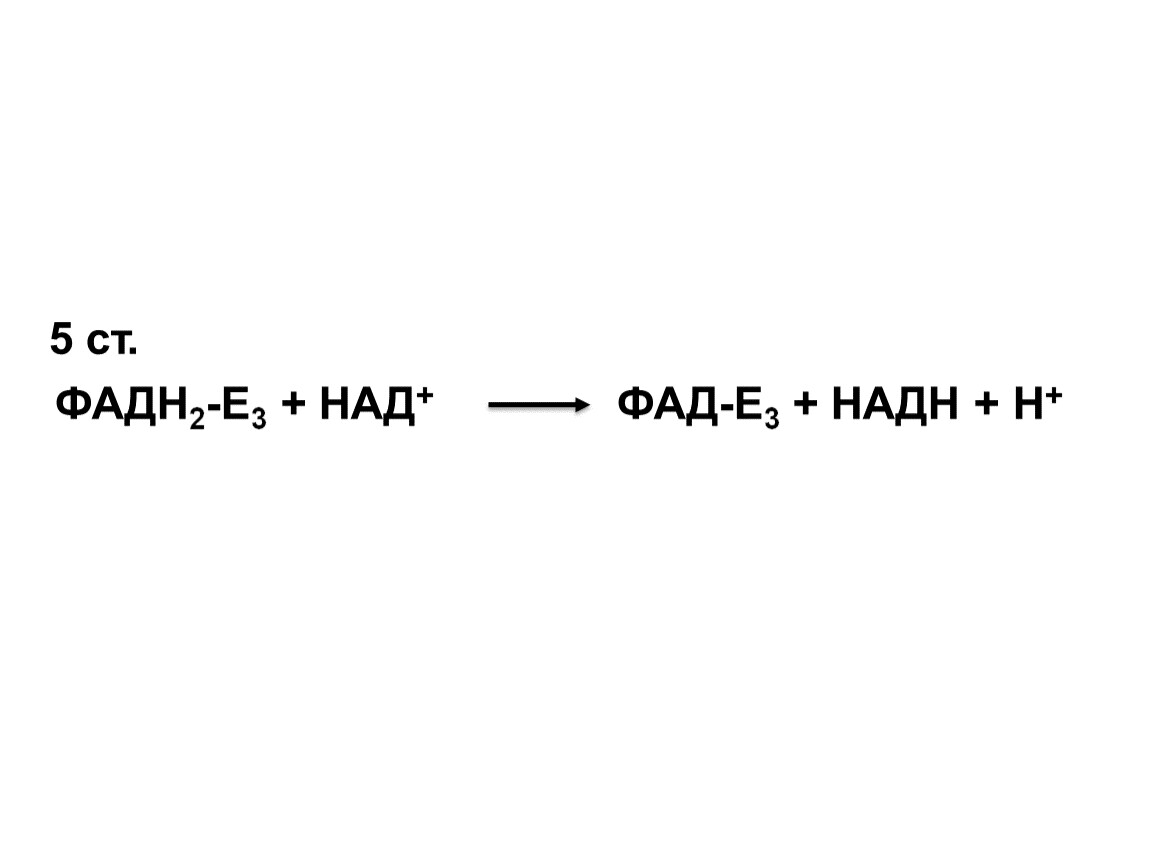
5 ст.
ФАДН2-Е3 + НАД+ ФАД-Е3 + НАДН + Н+
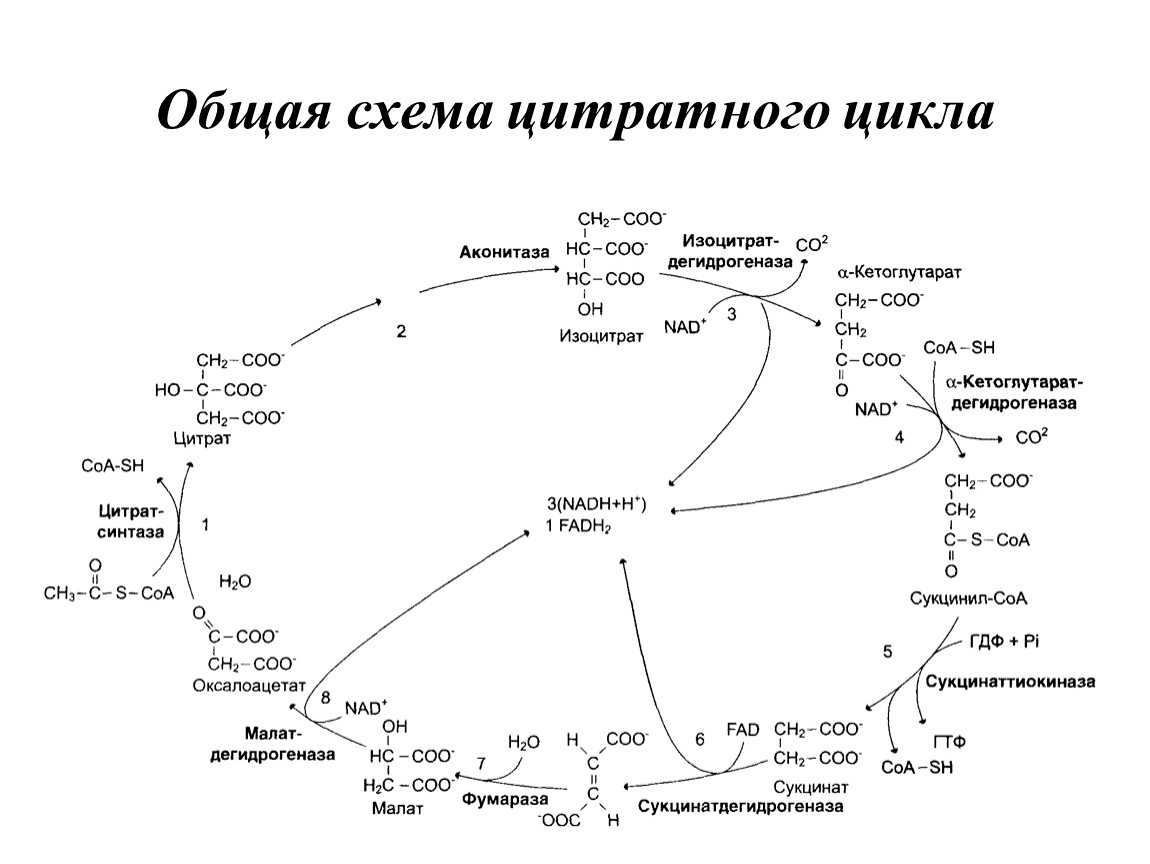
Общая схема цитратного цикла
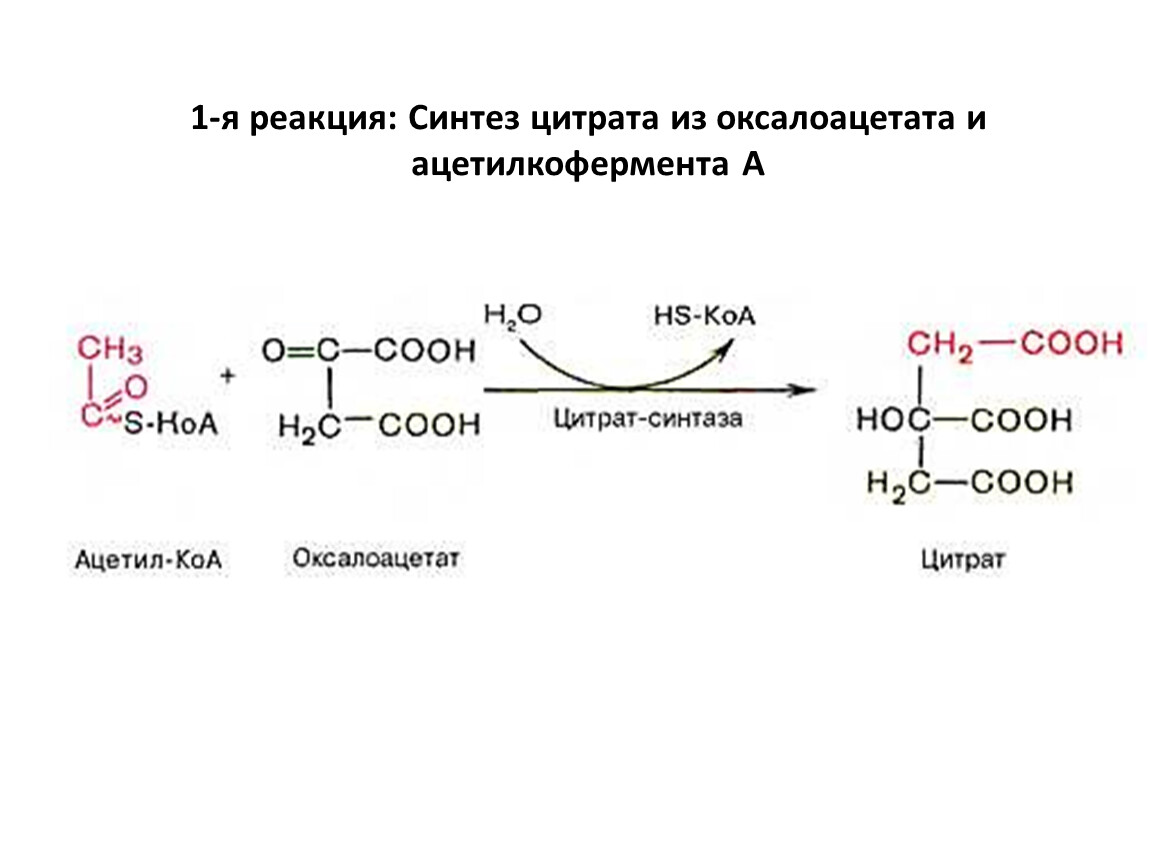
1-я реакция: Синтез цитрата из оксалоацетата и
ацетилкофермента А
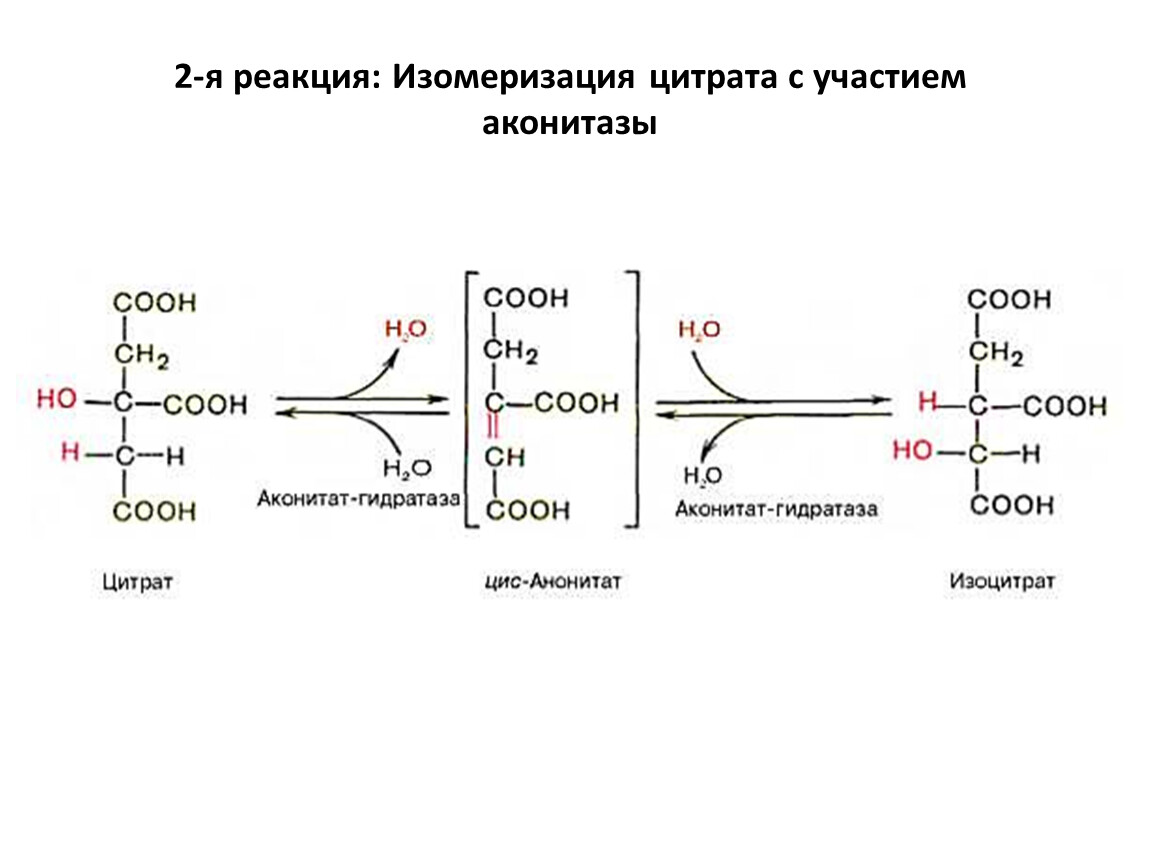
2-я реакция: Изомеризация цитрата с участием аконитазы
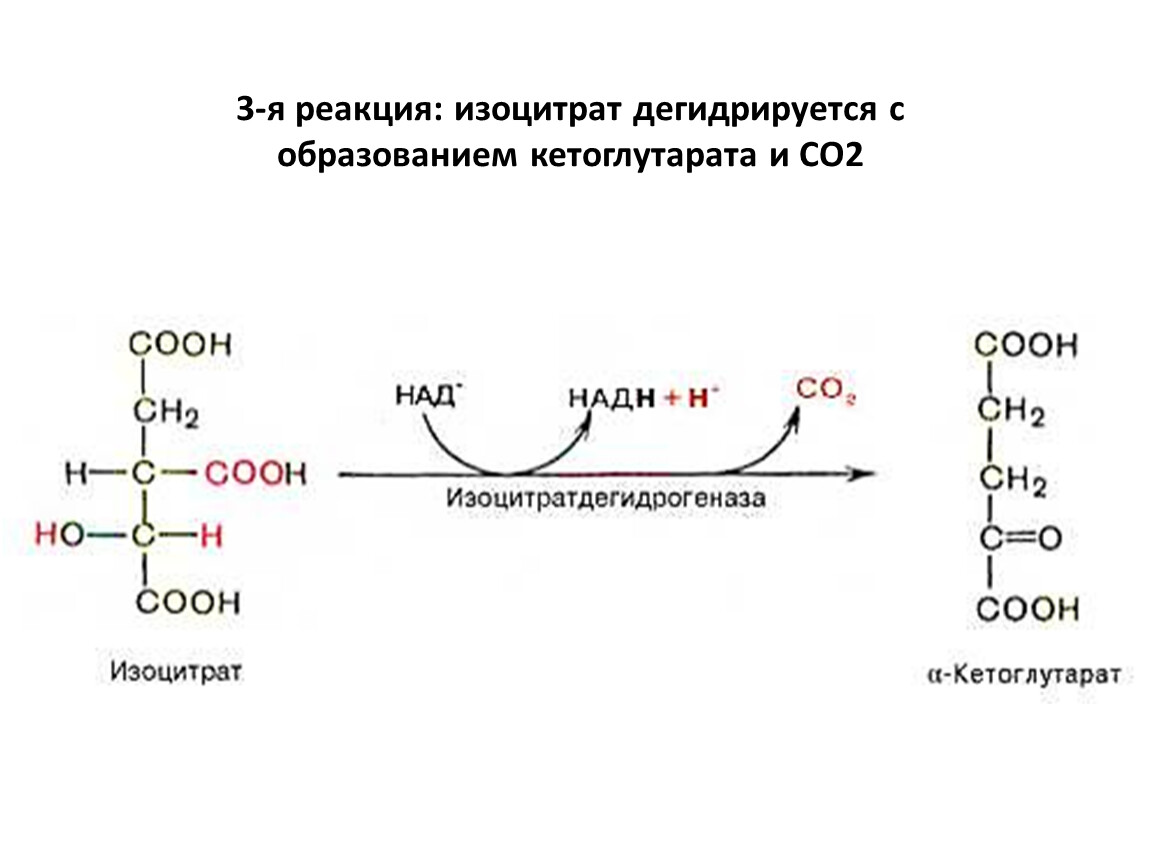
3-я реакция: изоцитрат дегидрируется с образованием кетоглутарата и CO2
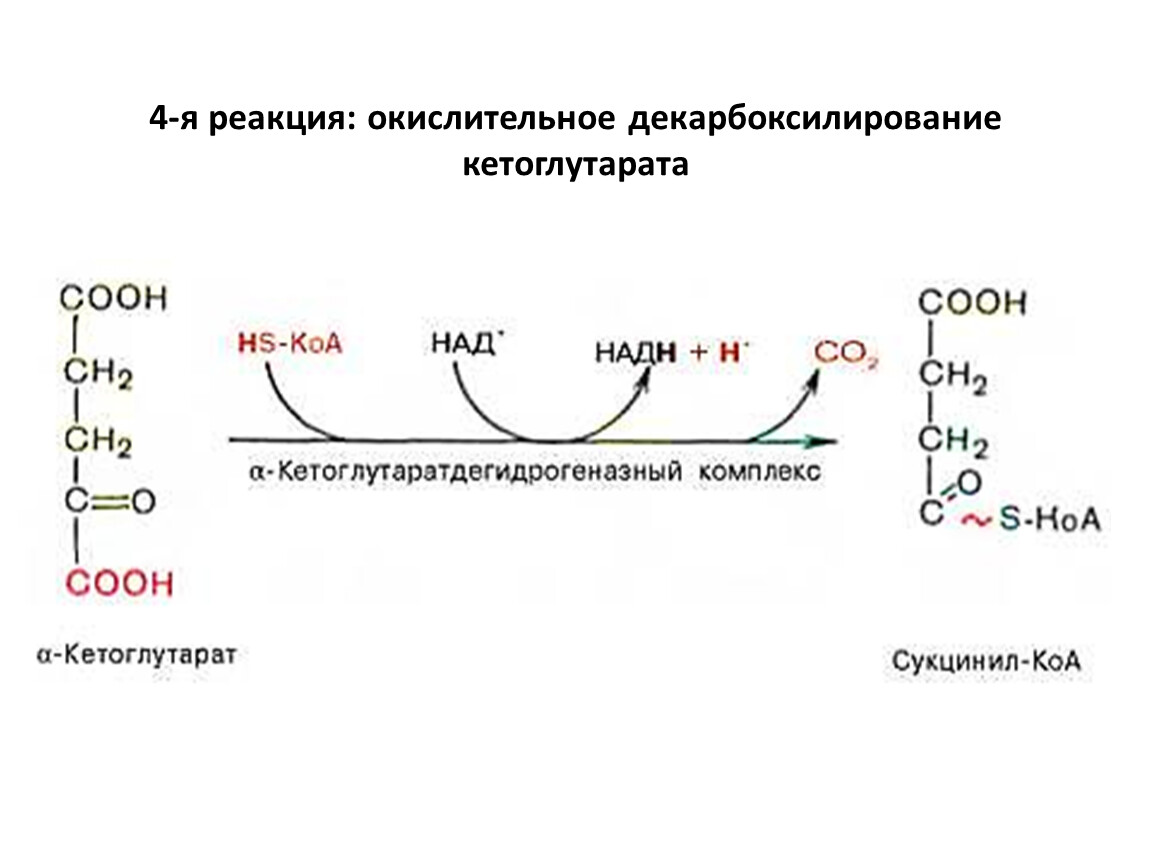
4-я реакция: окислительное декарбоксилирование кетоглутарата
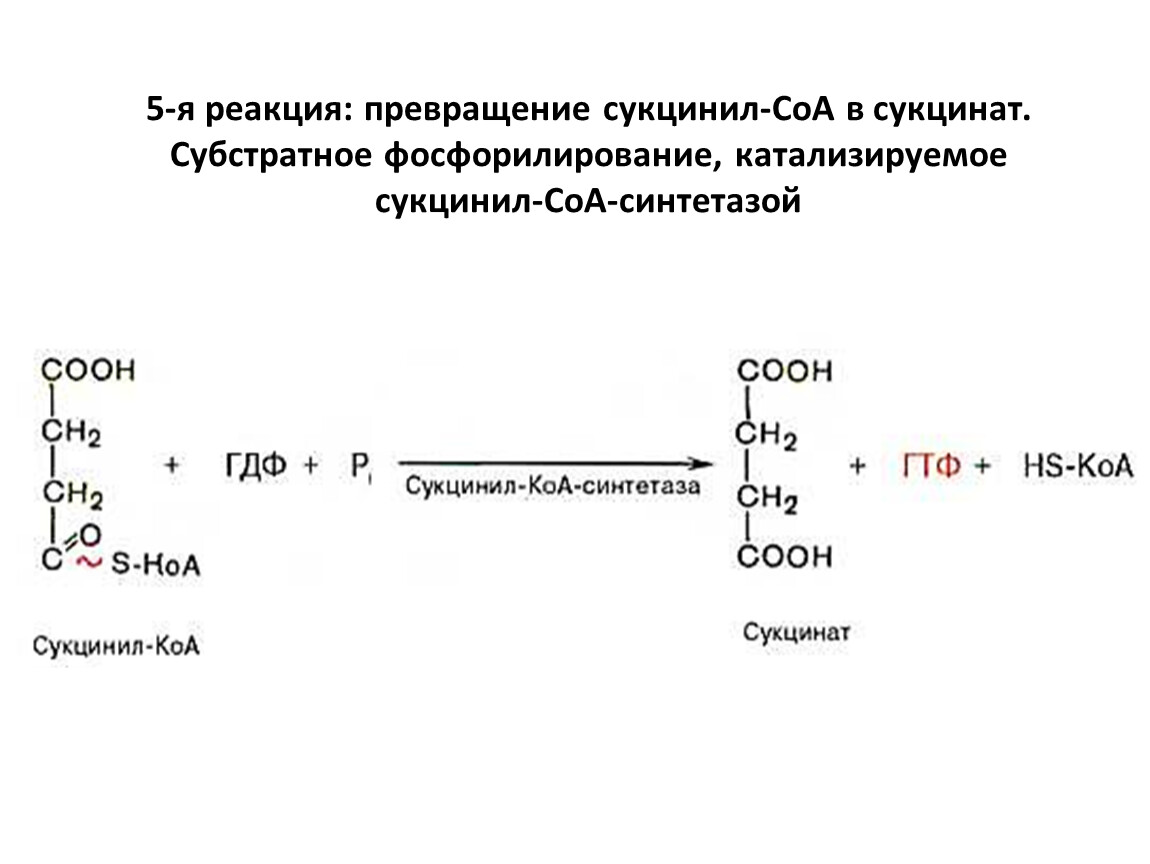
5-я реакция: превращение сукцинил-СоА в сукцинат. Субстратное фосфорилирование, катализируемое сукцинил-СоА-синтетазой
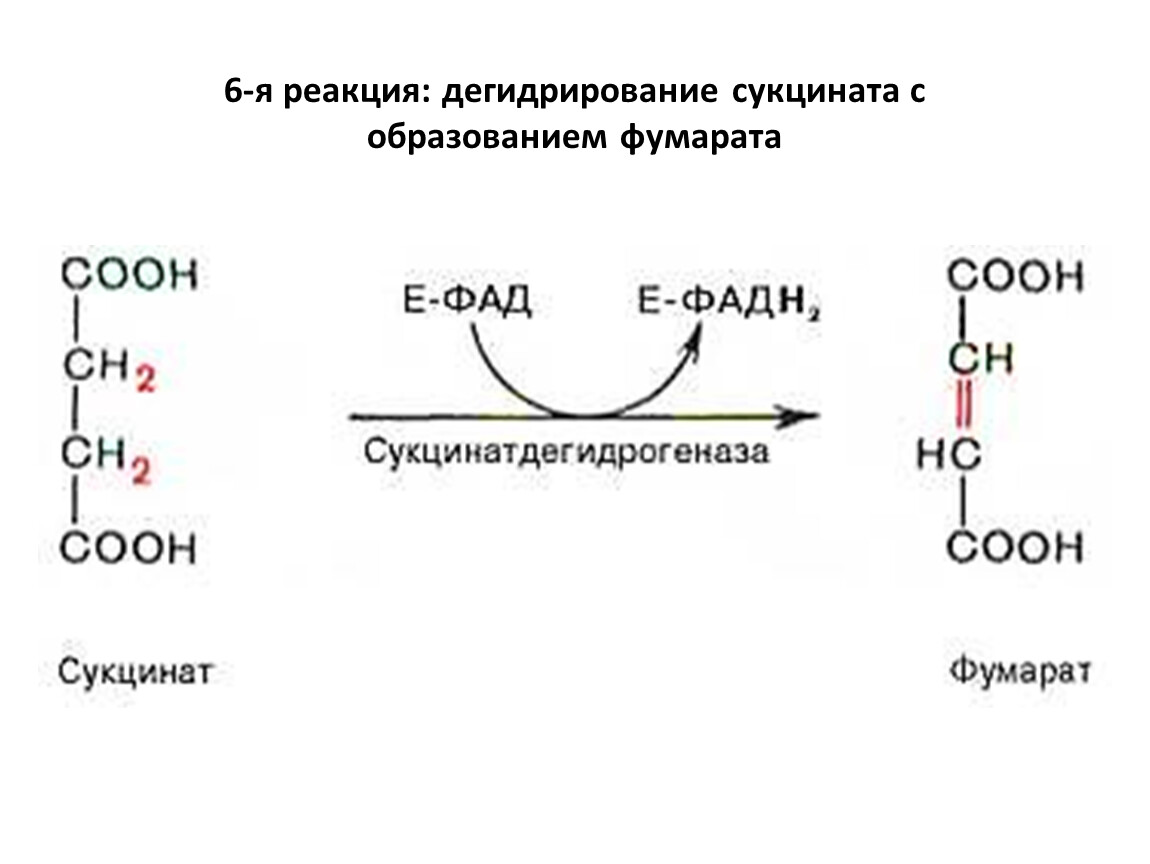
6-я реакция: дегидрирование сукцината с образованием фумарата

7-я реакция: гидратирование фумарата с образованием малата
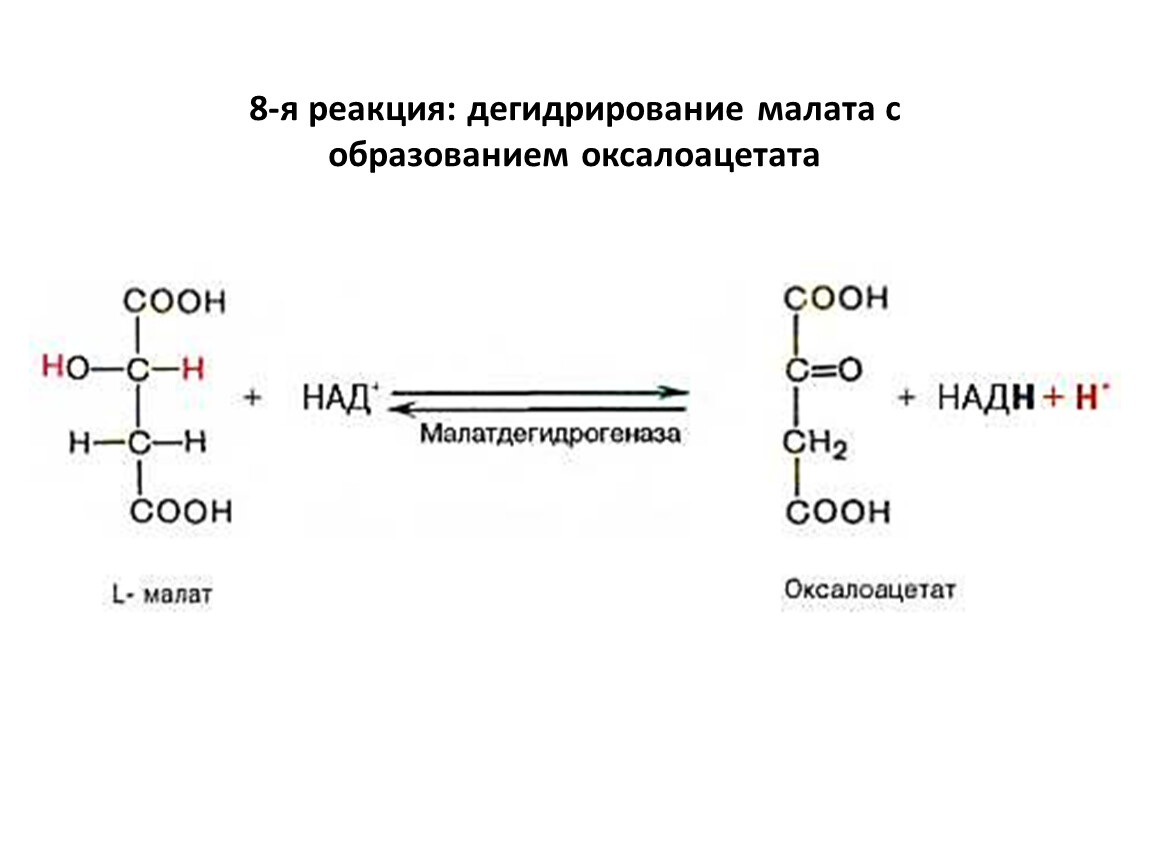
8-я реакция: дегидрирование малата с образованием оксалоацетата

Энергетический баланс аэробного гликолиза
I этап дает 8 молекул АТФ
II этап дает 6 молекул АТФ: 1 молекула ПВК образует при окислительном декарбоксилировании 3 молекулы АТФ; из 1 молекулы глюкозы образуется 2 молекулы ПВК, следовательно 2х3= 6;
III этап дает 24 молекулы АТФ, т.к. в цикл Кребса вступает 2 молекулы ацетилКоА.
Итого: 38 молекул АТФ.
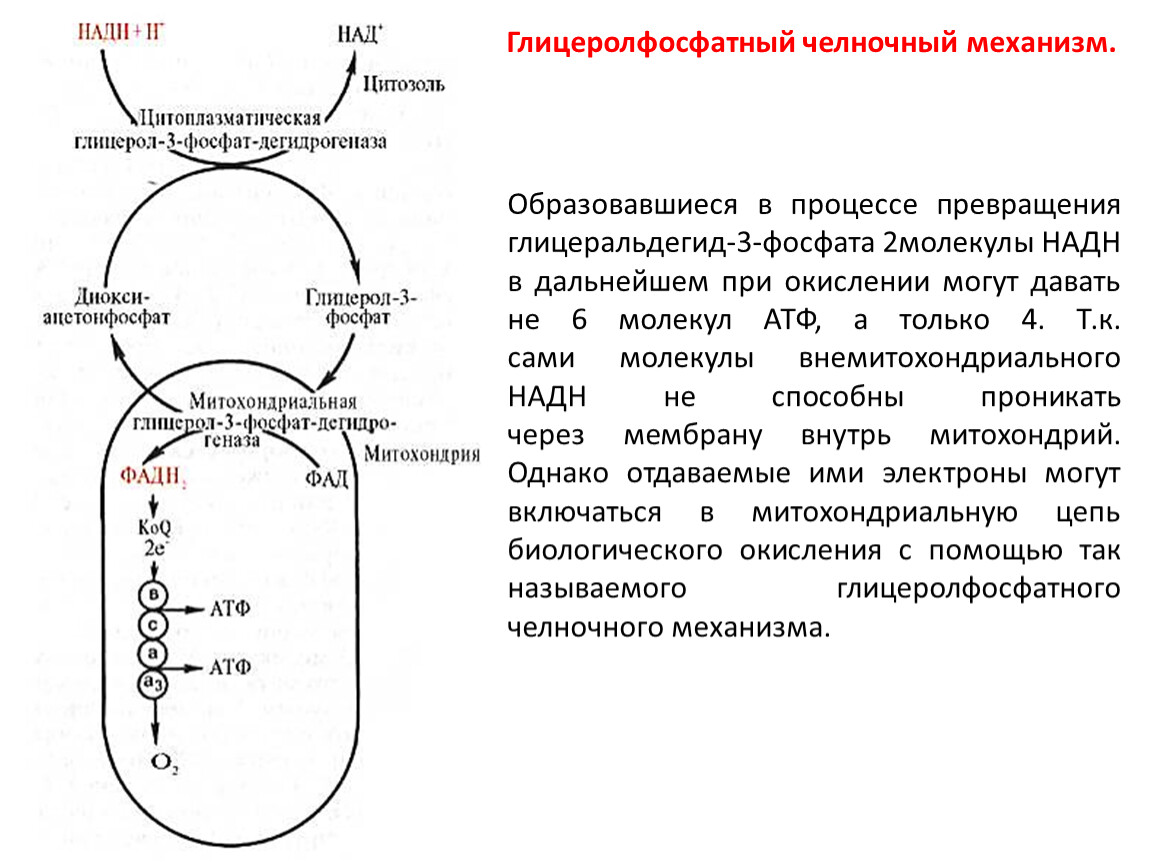
Глицеролфосфатный челночный механизм.
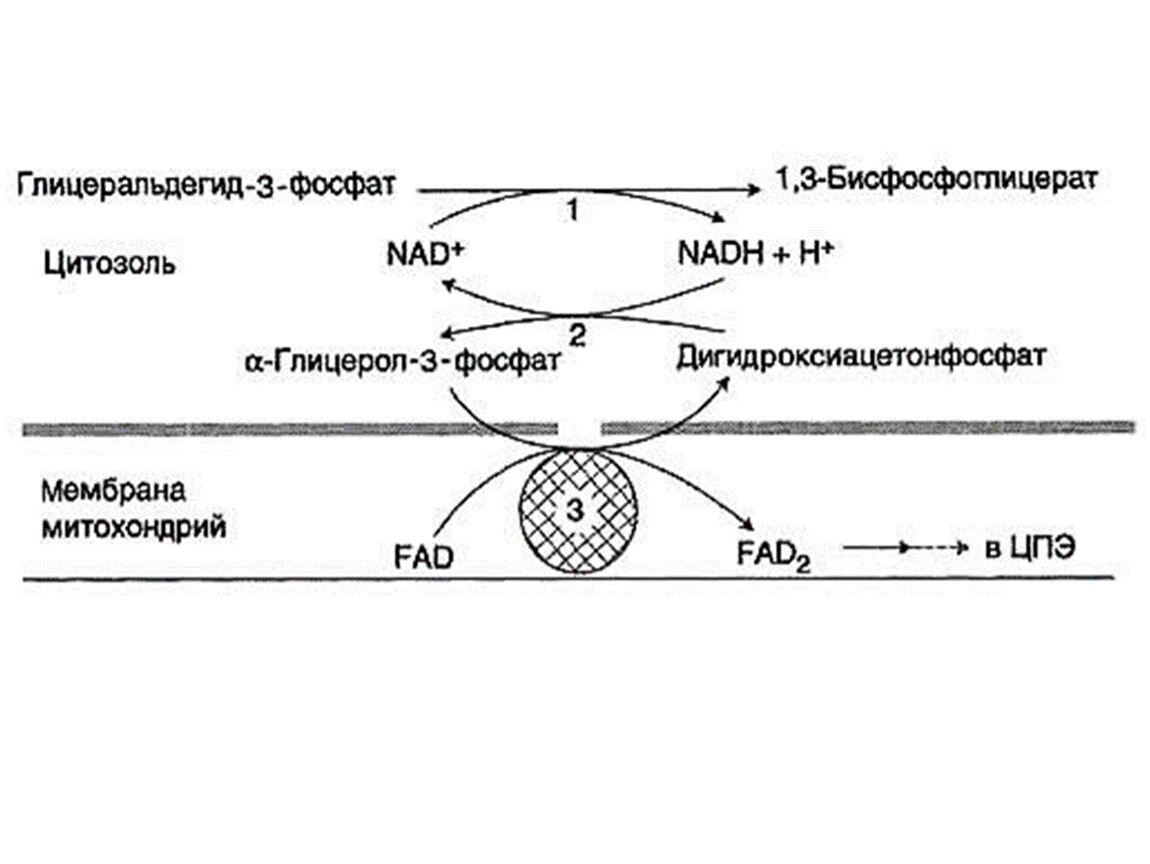
Образовавшиеся в процессе превращения глицеральдегид-3-фосфата 2молекулы НАДН в дальнейшем при окислении могут давать не 6 молекул АТФ, а только 4. Т.к. сами молекулы внемитохондриального НАДН не способны проникать через мембрану внутрь митохондрий. Однако отдаваемые ими электроны могут включаться в митохондриальную цепь биологического окисления с помощью так называемого глицеролфосфатного челночного механизма.
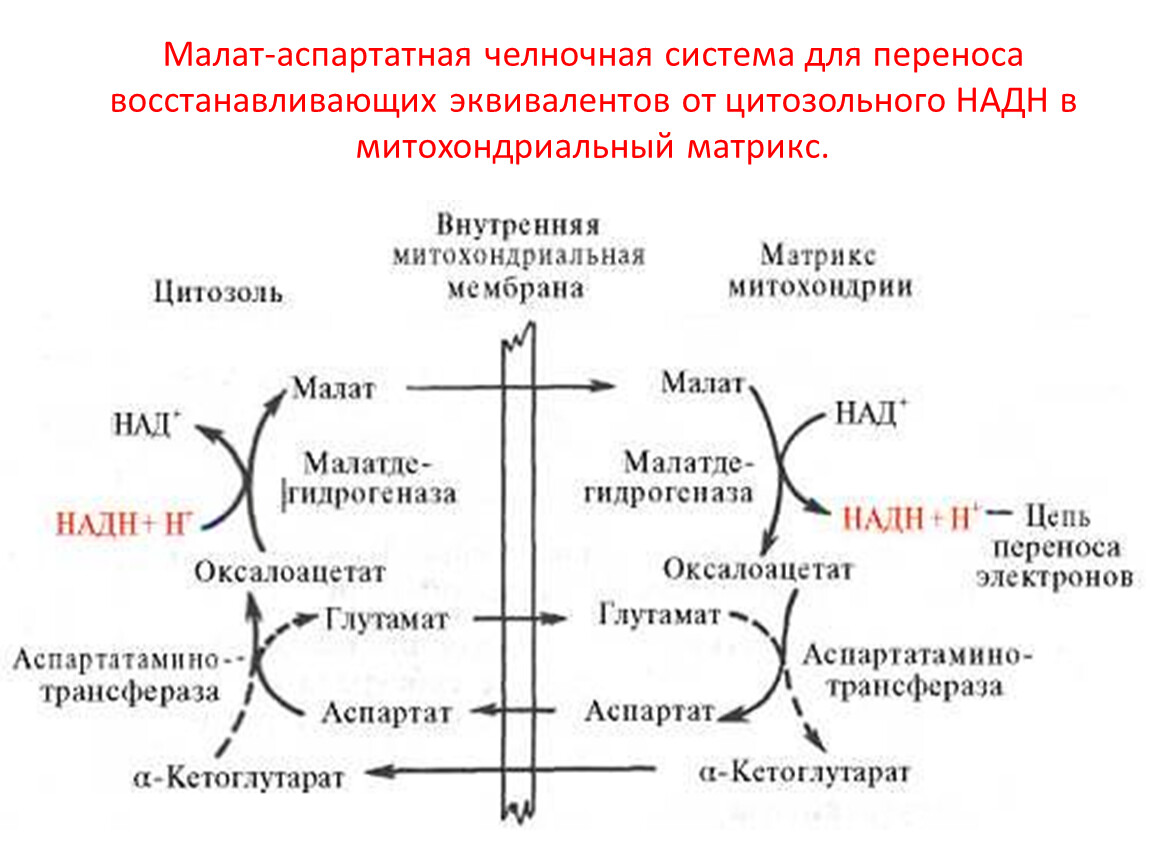
Малат-аспартатная челночная система для переноса восстанавливающих эквивалентов от цитозольного НАДН в митохондриальный матрикс.
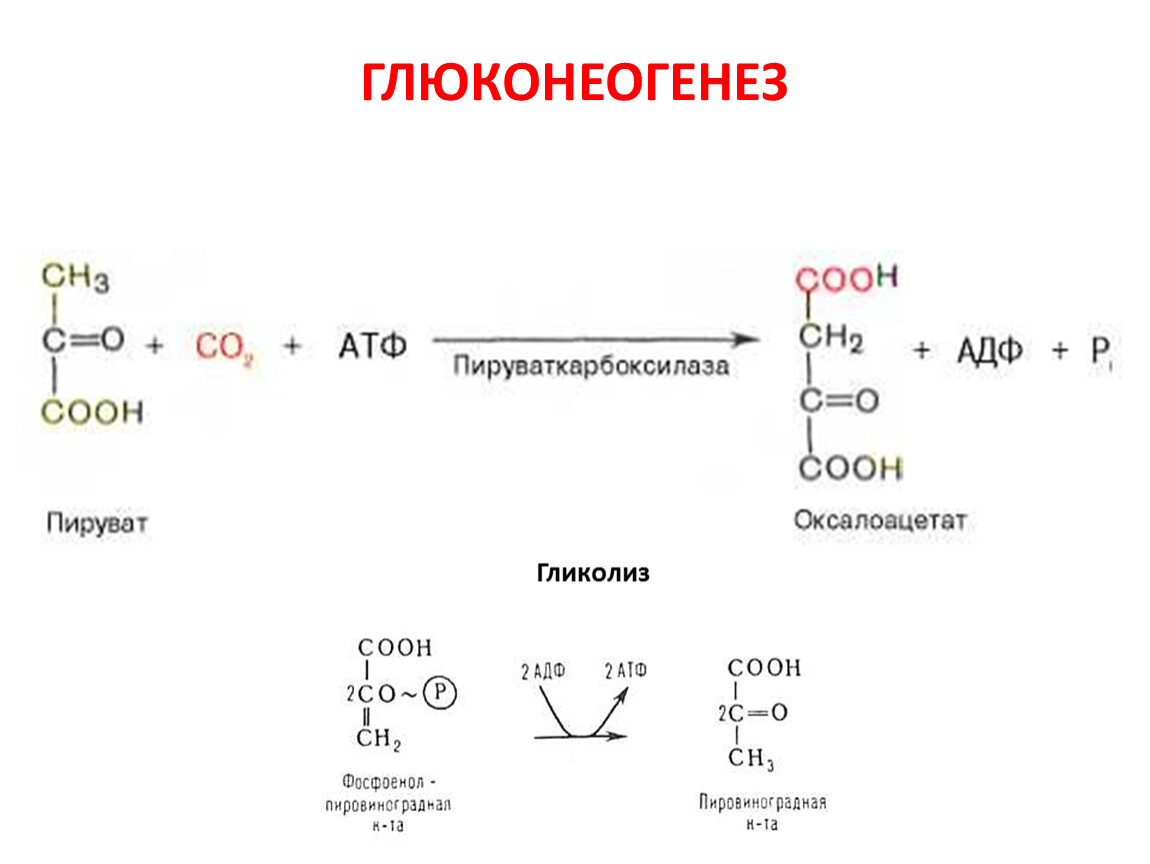
ГЛЮКОНЕОГЕНЕЗ
Гликолиз
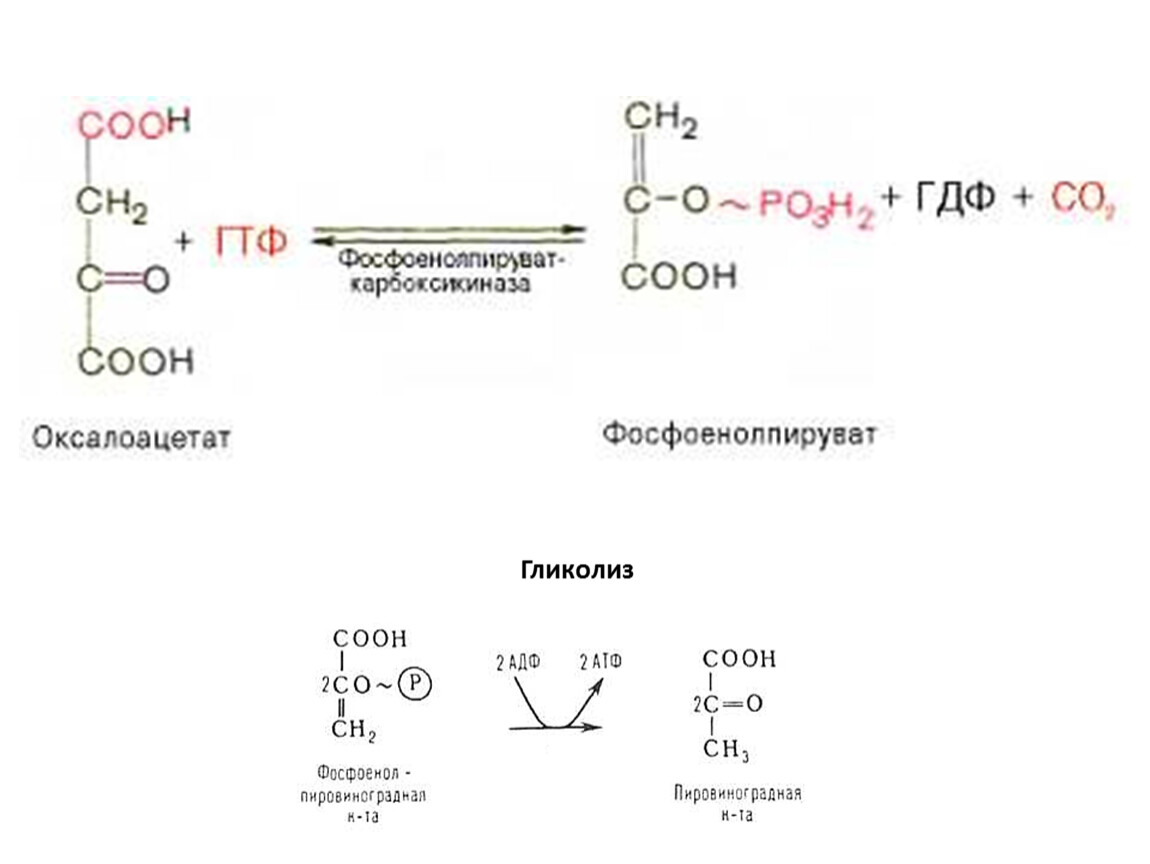
Гликолиз
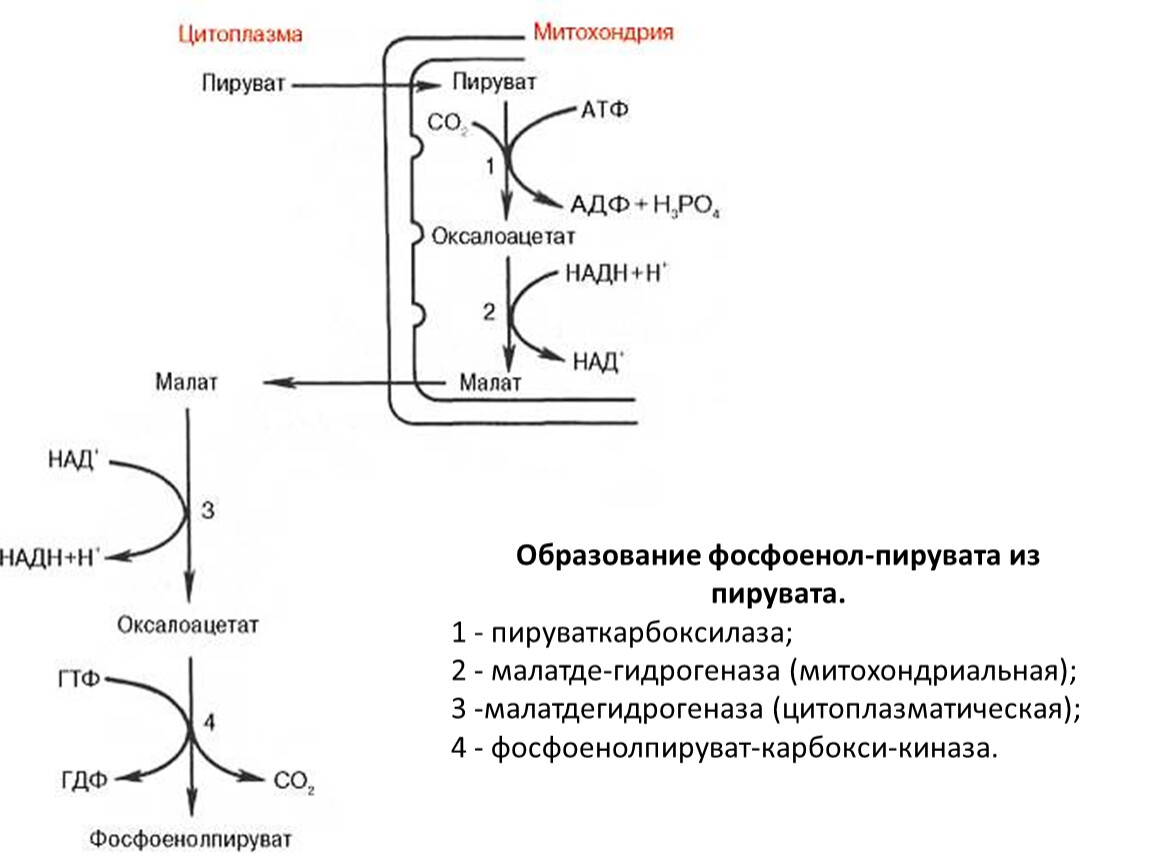
Образование фосфоенол-пирувата из пирувата.

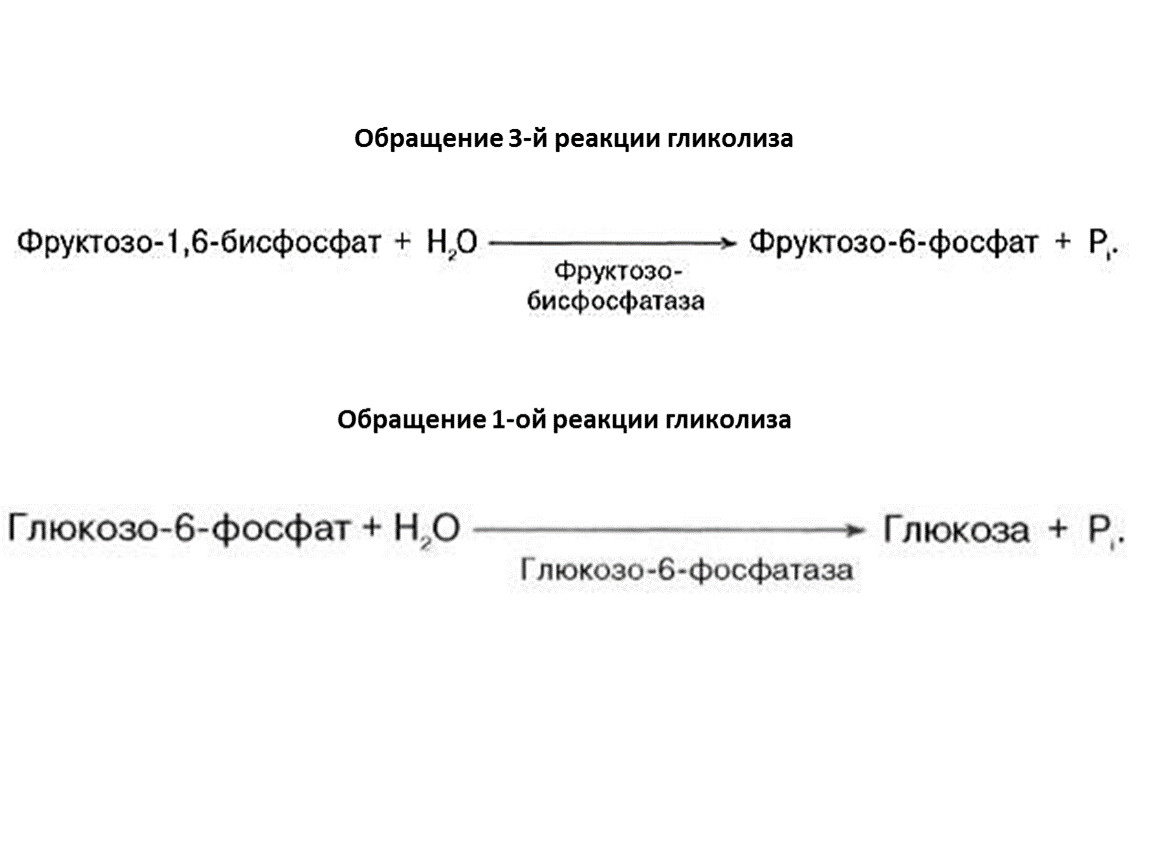
Обращение 3-й реакции гликолиза
Обращение 1-ой реакции гликолиза

Материалы на данной страницы взяты из открытых источников либо размещены пользователем в соответствии с договором-офертой сайта. Вы можете сообщить о нарушении.
