Биосинтез белка в природе. Проблема биосинтеза белка — одна из двух наиболее важных и острых проблем современного естествознания: если в неживой природе принципиально новые пути получения энергии будут найдены благодаря успехам физики элементарных частиц, то в живой природе решение кардинального вопроса управления самой жизнью может быть получено в результате познания химии и биологии белковых тел.
Биосинтез белков в организме осуществляется весьма интенсивно. Средняя скорость сборки полипептидных цепей в клетках бактерий составляет 16-17 аминокислотных остатков в 1 с, дрожжей — 7-10, млекопитающих — 5-7. Время синтеза (в с) молекулы глобина в ретикулоцитах кролика равно 20, овальбумина в яйцеводах курицы — 80; суммарных белков в печени крысы — 80. За 1 мин в ретикулоците кролика синтезируется 5∙104 молекул глобина, в клетке яйцеводов курицы — 6•105 молекул овальбумина, в гигантской клетке заднего отдела шелкоотделительной железы тутового шелкопряда — 38-1011 молекул фиброина шелка.
Метаболизм белков.
1.
2.
3.
БИОСИНТЕЗ БЕЛКОВ В ПРИРОДЕ. ПРОТЕОМИКА.
СТРОЕНИЕ РИБОСОМ.
ГЕНЕТИЧЕСКИЙ КОД
Биосинтез белка в природе. Проблема биосинтеза белка — одна из двух наиболее
важных и острых проблем современного естествознания: если в неживой природе
принципиально новые пути получения энергии будут найдены благодаря успехам физики
элементарных частиц, то в живой природе решение кардинального вопроса управления самой
жизнью может быть получено в результате познания химии и биологии белковых тел.
Биосинтез белков в организме осуществляется весьма интенсивно. Средняя скорость
сборки полипептидных цепей в клетках бактерий составляет 1617 аминокислотных остатков
в 1 с, дрожжей — 710, млекопитающих — 57. Время синтеза (в с) молекулы глобина в
ретикулоцитах кролика равно 20, овальбумина в яйцеводах курицы — 80; суммарных белков
в печени крысы — 80. За 1 мин в ретикулоците кролика синтезируется 5∙104 молекул глобина,
в клетке яйцеводов курицы — 6•105 молекул овальбумина, в гигантской клетке заднего
отдела шелкоотделительной железы тутового шелкопряда — 381011 молекул фиброина
шелка.
История развития представлений о механизме биосинтеза белков. Первой по
времени была гипотеза обращения протеолиза. В 1886 г. А.Я. Данилевский наблюдал
образование белковоподобных веществ при действии ферментов желудочного сока на
концентрированный раствор пептонов, возникших в результате расщепления белка пепсином.
Позже был расширен круг ферментов, способных к обращению протеолиза (трипсин, пепсин,
папаин, катепсины), и белков, с неполными гидролизатами которых такое обращение
удавалось осуществить (альбумины, глобулины, фибрин, казеин и др.). Продукты,
возникающие в результате обращения реакции гидролиза белков, назвали пластеинами.
Определенный вклад в развитие современных представлений о механизме биосинтеза
белков внесли исследования биосинтеза квазипептидных связей (т. е. не истинных пептидных
связей, но близких к ним), осуществленные в 50е годы в ряде лабораторий, в том числе у нас
в лаборатории А.Е. Браунштейна. Эти исследования доказали ферментативный характер
реакций их образования, необходимость энергообеспечения за счет окислительных процессов
и, что самое важное, участие в биосинтезе квазипептидных связей АТФ.
Существенными оказались результаты разработки гипотезы транспептидирования в
качестве возможного варианта механизма биосинтеза пептидных связей. В нашей стране
исследования в этом направлении интенсивно проводились в 50е годы В.Н. Ореховичем с
сотр. При изучении реакций транспептидирования впервые было показано, что перенос
аминоациальных или пептидильных группировок на аминогруппу аминокислот может
происходить не только с амидной или пептидной связи, но и со сложноэфирной связи. В
дальнейшем оказалось, что именно этот механизм лежит
основе реакции
транспептидирования в рибосоме.
в
Переломным моментом в развитии подходов к выяснению механизма биосинтеза
белковых тел было установление сцепленности его с биосинтезом РНК (Ж. Браше, 1941; Т.
Оно привело к созданию матричной схемы биосинтеза белков,
Касперсон,
являющейся фундаментом современных представлений.
1941).
1Общая схема матричного биосинтеза
белковых тел
Матричный механизм биосинтеза полимеров, обеспечивающий безошибочное
воспроизведение их первичной структуры, представляет одну из наиболее специфических
черт живого.
Принципиальное значение в разработке вопроса о механизме биосинтеза белков имело
выявление локализации его в рибосомальном аппарате клетки и создание бесклеточных
систем, где единственной структурой, на которой протекал биосинтез белка, были рибосомы.
Наряду с матричным механизмом в природе существует мультиэнзимный путь
биосинтеза белков и пептидов, где специфическое чередование аминокислот обеспечивается
расположением каталитически активных белков в мультиэнзимном комплексе.
Исследования по выяснению механизма синтеза белков положили начало науки
протеомики. Осуществление внеклеточного синтеза белка в биореакторах настоящее время
позволяет получать белки самого различного состава.
Биосинтез белков во всех структурных элементах клетки (ядро, митохондрии,
хлоропласты, эндоплазматический ретикулум и др.) идет на рибосомах. Поэтому именно на
пути исследования структуры и свойств рибосом, а также механизма их взаимодействия с
исходными для биосинтеза белков соединениями достигнуты наиболее впечатляющие
результаты в выявлении закономерностей новообразования белковых тел.
Строение и свойства рибосом.
Рибосомы — мельчайшие тельца
рибонуклеопротеиновой природы. Они содержатся в основном в цитоплазме растительных,
животных и бактериальных клеток в количестве нескольких десятков тысяч в каждой.
Рибосомы выделяют путем дифференциального центрифугирования клеточного
содержимого, получаемого после гомогенизации клеток. Известно, что этим путем удается
разделить клеточное содержимое на ряд фракций. Рибосомы связаны главным образом с
мембранами эндоплазматической сети. Поэтому при фракционировании содержимого клетки
они осаждаются вместе с обломками липопротеиновой мембраны — микросомами.
Форма и структура рибосом, выделенных из разных источников, сходна, но по
значению константы седиментации их делят на два класса: 70S и 80S. Первые найдены у всех
2прокариот, а также содержатся в ядрах, митохондриях и хлоропластах эукариот; вторые
локализованы в цитоплазме только эукариот.
70S и 80S рибосомы стабильны при концентрации Mg2+, близкой к 0,001 М. При
повышении содержания Mg2+ до 0,01 М рибосомы дают димеры или слипаются в более
крупные агрегаты, а при понижении до 0,0001 М диссоциируют на субчастицы: 70(80)S =
30(40)S + 50(60)S. Этот процесс диссоциации—ассоциации рибосом и их субчастиц —
контролируется также полиаминами и изменением концентрации других двухвалентных
катионов.
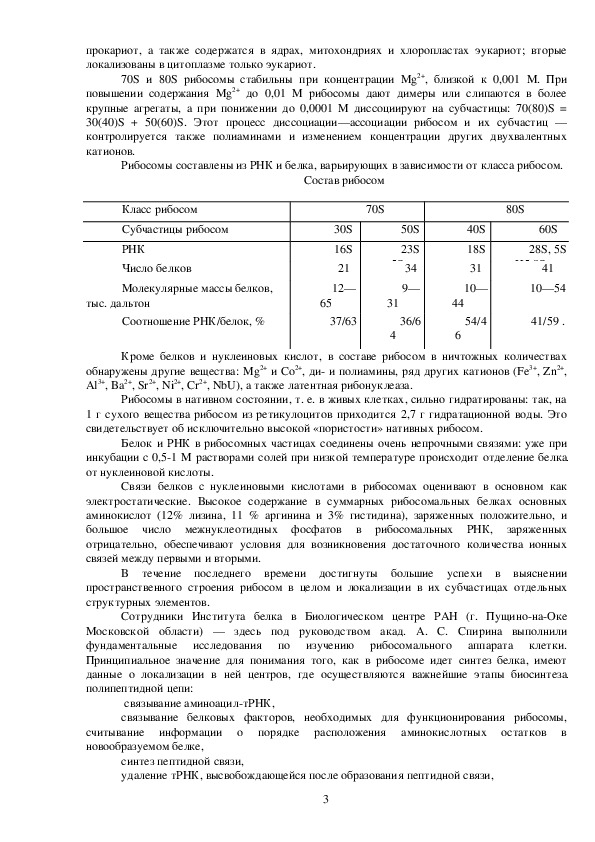
Рибосомы составлены из РНК и белка, варьирующих в зависимости от класса рибосом.
Состав рибосом
Класс рибосом
Субчастицы рибосом
РНК
Число белков
Молекулярные массы белков,
тыс. дальтон
Соотношение РНК/белок, %
30S
16S
21
12—
65
37/63
70S
50S
23S
34
9—
и 5S
31
4
36/6
80S
60S
28S, 5S
H5.8S
41
10—54
40S
18S
31
10—
44
6
54/4
41/59 .
Кроме белков и нуклеиновых кислот, в составе рибосом в ничтожных количествах
обнаружены другие вещества: Mg2+ и Со2+, ди и полиамины, ряд других катионов (Fe3+, Zn2+,
Al3+, Ba2+, Sr2+, Ni2+, Cr2+, NbU), а также латентная рибонуклеаза.
Рибосомы в нативном состоянии, т. е. в живых клетках, сильно гидратированы: так, на
1 г сухого вещества рибосом из ретикулоцитов приходится 2,7 г гидратационной воды. Это
свидетельствует об исключительно высокой «пористости» нативных рибосом.
Белок и РНК в рибосомных частицах соединены очень непрочными связями: уже при
инкубации с 0,51 М растворами солей при низкой температуре происходит отделение белка
от нуклеиновой кислоты.
Связи белков с нуклеиновыми кислотами в рибосомах оценивают в основном как
электростатические. Высокое содержание в суммарных рибосомальных белках основных
аминокислот (12% лизина, 11 % аргинина и 3% гистидина), заряженных положительно, и
большое число межнуклеотидных фосфатов в рибосомальных РНК, заряженных
отрицательно, обеспечивают условия для возникновения достаточного количества ионных
связей между первыми и вторыми.
В течение последнего времени достигнуты большие успехи в выяснении
пространственного строения рибосом в целом и локализации в их субчастицах отдельных
структурных элементов.
Сотрудники Института белка в Биологическом центре РАН (г. ПущинонаОке
Московской области) — здесь под руководством акад. А. С. Спирина выполнили
фундаментальные исследования по изучению рибосомального аппарата клетки.
Принципиальное значение для понимания того, как в рибосоме идет синтез белка, имеют
данные о локализации в ней центров, где осуществляются важнейшие этапы биосинтеза
полипептидной цепи:
связывание аминоацилтРНК,
связывание белковых факторов, необходимых для функционирования рибосомы,
считывание информации о порядке расположения аминокислотных остатков в
новообразуемом белке,
синтез пептидной связи,
удаление тРНК, высвобождающейся после образования пептидной связи,
3перенос пептидилтРНК из аминоацильного центра в пептидильный центр с
одновременным продвижением мРНК на один триплет нуклеотидных остатков,
гидролиз сложноэфирной связи между полипептидом и тРНК в момент завершения
синтеза белка в рибосоме и др.
Пространственная структура 30S субчастицы рибосомы кишечной палочки:
Как перевести четырехбуквенный (по числу оснований, входящих в мРНК, т. е. А
аденин, Ггуанин, Ццитозин и Уурацил) код в двадцатибуквенный (по числу аминокислот,
составляющих белковую молекулу)?
Если каждой комбинации нуклеотидов в мРНК приписать способность кодировать
положение одной аминокислоты в белке, то дуплетный код вряд ли возможен (число пар
нуклеотидов менее числа постоянно встречающихся в белке аминокислот), квадруплетный
нереален (число сочетаний слишком сильно превышает число аминокислот), в то время как
триплетный код наиболее удовлетворяет численному соотношению возможных кодонов и
белковых аминокислот. Указанные расчеты основаны на том, что при соединении 4
нуклеотидов попарно можно получить 16 комбинаций, по три—64, по четыре—256
комбинаций и т. д.:
Посредством ряда опытов триплетная природа кода белкового синтеза доказана
экспериментально. Сначала был установлен качественный состав нуклеотидных остатков,
которые входят в состав одного или нескольких кодонов, обеспечивающих вступление в
состав белка той или иной аминокислоты. Решающую роль здесь сыграло наблюдение М.
Ниренберга (1961), который, используя в качестве мРНК полиуридиловую кислоту, впервые
показал, что вступление в полипептидную цепь фенилаланина кодируется УУУтриплетом.
Полиуридиловая кислота, поли (У), вводилась им в белоксинтезирующую бесклеточную
систему, составленную из промытых (лишенных мРНК) рибосом, полного набора тРНК,
аминоацилтРНКсинтетаз и аминокислот (некоторые из последних мечены 14С или 15N), АТФ
и генерирующих ее соединений (фосфоенолпируват, ацетилфосфат и подобные им вещества),
ГТФ, Mg2+ и Мп . В этой системе на поли (У) в качестве матрицы шел синтез пептидов,
составленных только из остатков фенилаланина. Благодаря применению в этой же системе
почти полного набора синтетических гомои гетерополирибонуклеотидных матриц,
4полученных с помощью полинуклеотидфосфорилазы, в лаборатории С. Очоа в течение года
была завершена работа по выявлению качественного состава кодонов для всех аминокислот.
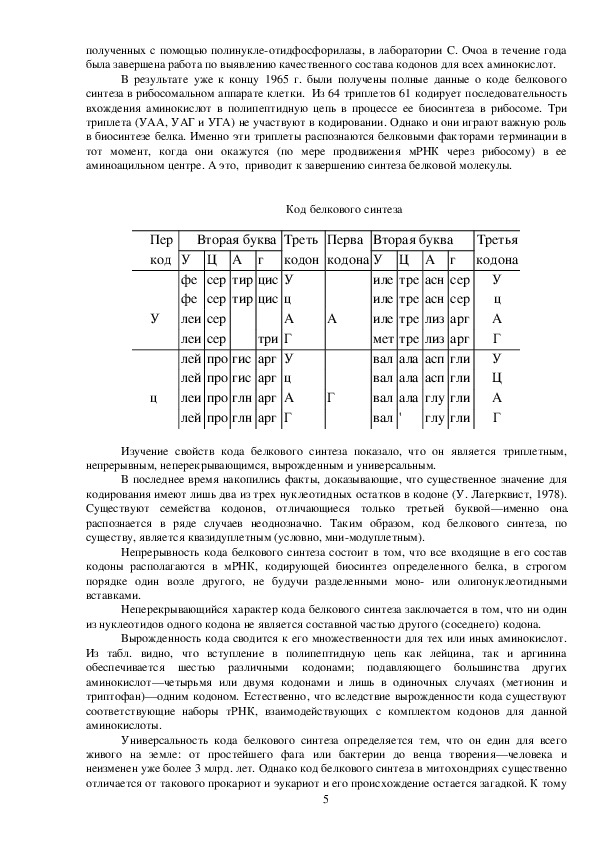
В результате уже к концу 1965 г. были получены полные данные о коде белкового
синтеза в рибосомальном аппарате клетки. Из 64 триплетов 61 кодирует последовательность
вхождения аминокислот в полипептидную цепь в процессе ее биосинтеза в рибосоме. Три
триплета (УАА, УАГ и УГА) не участвуют в кодировании. Однако и они играют важную роль
в биосинтезе белка. Именно эти триплеты распознаются белковыми факторами терминации в
тот момент, когда они окажутся (по мере продвижения мРНК через рибосому) в ее
аминоацильном центре. А это, приводит к завершению синтеза белковой молекулы.
Код белкового синтеза
Пер
код
вая
она
У
ц
Вторая буква
Треть
я
кодона
кодон
У Ц А г
а
тир цис У
фе
сер
н
.
сер тир цис ц
фе
н
леи сер
А
леи сер
три Г
лей про гис арг У
лей про гис арг ц
леи про
глн арг А
■
лей про глн арг Г
А
Перва
Вторая буква
кодона
я
кодона У Ц А г
иле тре асн сер
иле тре асн сер
иле тре лиз арг
мет тре лиз арг
вал ала асп гли
вал ала асп гли
вал ала глу гли
вал '
глу гли
Г
Третья
кодона
У
ц
А
Г
У
Ц
А
Г
Изучение свойств кода белкового синтеза показало, что он является триплетным,
непрерывным, неперекрывающимся, вырожденным и универсальным.
ала
В последнее время накопились факты, доказывающие, что существенное значение для
кодирования имеют лишь два из трех нуклеотидных остатков в кодоне (У. Лагерквист, 1978).
Существуют семейства кодонов, отличающиеся только третьей буквой—именно она
распознается в ряде случаев неоднозначно. Таким образом, код белкового синтеза, по
существу, является квазидуплетным (условно, мнимодуплетным).
Непрерывность кода белкового синтеза состоит в том, что все входящие в его состав
кодоны располагаются в мРНК, кодирующей биосинтез определенного белка, в строгом
порядке один возле другого, не будучи разделенными моно или олигонуклеотидными
вставками.
Неперекрывающийся характер кода белкового синтеза заключается в том, что ни один
из нуклеотидов одного кодона не является составной частью другого (соседнего) кодона.
Вырожденность кода сводится к его множественности для тех или иных аминокислот.
Из табл. видно, что вступление в полипептидную цепь как лейцина, так и аргинина
обеспечивается шестью различными кодонами; подавляющего большинства других
аминокислот—четырьмя или двумя кодонами и лишь в одиночных случаях (метионин и
триптофан)—одним кодоном. Естественно, что вследствие вырожденности кода существуют
соответствующие наборы тРНК, взаимодействующих с комплектом кодонов для данной
аминокислоты.
Универсальность кода белкового синтеза определяется тем, что он един для всего
живого на земле: от простейшего фага или бактерии до венца творения—человека и
неизменен уже более 3 млрд. лет. Однако код белкового синтеза в митохондриях существенно
отличается от такового прокариот и эукариот и его происхождение остается загадкой. К тому
5же у реснитчатых и микоплазм он тоже отличается от канонического: терминирующие кодоны
у них выполняют кодирующую функцию.
Всегда ли кодирование осуществляется с абсолютной точностью или возможно ложное
кодирование и включение аминокислоты в полипептидную цепь не в соответствии со
структурой кодона? Считают, что определенный уровень ложного кодирования
запрограммирован эволюционно и в тех случаях, когда вследствие мутации изменен один из
кодонов в мРНК, он может транслироваться как неизменённый («ложь во спасение»). Это
помогает клеткам, в том числе и бактериальным, выжить при неблагоприятных мутациях.
Вопросы ложного кодирования в процессе белкового синтеза успешно изучаются в
Институте белка в Пущино под руководством акад. А. С. Спирина.
Проблема кодирования биосинтеза белков в последние годы обсуждается еще в одном
принципиальном аспекте. Исходя из работ, в которых удалось доказать способность
некоторых белков без участия нуклеиновых кислот обеспечивать собственное
мультиплицирование,
накопление и реализацию присущего им патологического
(инфекционного) процесса, В. А. Кордюмом выдвинута концепция о существовании новой
формы биологической информации—пространственной структуры белка, способной к
самовоспроизведению.
6