Нуклеиновые кислоты (РНК и ДНК) распадаются в организме при посредстве особых ферментов — нуклеаз. Они ускоряют реакцию разрыва межнуклеотидных фосфодиэфирных связей в молекулах нуклеиновых кислот и принадлежат, к классу ферментов гидролаз — фосфодиэстераз.
Нуклеазы, действующие на внутренние межнуклеотидные связи в молекулах ДНК и РНК, называются эндонуклеазами.
При их участии осуществляется деполимеризация нуклеиновых кислот до олигонуклеотидов.
Нуклеазы, ускоряющие реакции последовательного отщепления нуклеотидов от РНК, ДНК или их фрагментов, начиная с конца полинуклеотидной цепи, называют экзонуклеазами. Они обеспечивают распад нуклеиновых кислот до свободных нуклеотидов.
В зависимости от специфичности действия среди нуклеаз различают рибонуклеазы и дезоксирибонуклеазы. Первые ускоряют реакции распада как внутренних, так и внешних (концевых) межнуклеотидных связей в молекулах РНК. Вторые выполняют такую же функцию по отношению к ДНК. Существует большая группа неспецифических эндо- и экзонуклеаз, действующих одновременно и на РНК, и на ДНК.
Метаболизм нуклеиновых кислот.
1. Обмен нуклеиновых кислот
2. Распад нуклеотидов
3. распад пуриновых и пиримидиновых нуклеотидов.
4. Синтез нуклеотидов.
Обмен нуклеиновых кислот. Изучение обмена нуклеиновых кислот имеет большое
значение. Это имеет важное значение для глубокого понимания процессов жизнедеятельности
организмов.
Исследование молекулярных механизмов биосинтеза пуриновых и пиримидиновых
оснований позволило открыть ряд важнейших закономерностей в регуляции новообразования
этих соединений и сформулировать общие принципы регуляции обмена веществ.
Раскрытие механизма специфического биосинтеза молекул полинуклеотидов, при
осуществлении обеспечивается порядок чередования мононуклеотидных звеньев, их
составляющих, привело к признанию ведущей роли в этом процессе взаимодействия
комплементарных пуриновых и пиримидиновых оснований. Это, в свою очередь, дало
возможность впервые понять механизм обеспечения специфического воспроизведения
первичной структуры макромолекул при их биосинтезе. Данные о регуляции новообразования
нуклеиновых кислот привели к фундаментальным открытиям, позволяющим сделать первые
шаги к объяснению закономерностей не только воспроизведения специфических
макромолекул, но также и морфогенеза.
Распад нуклеиновых кислот в организме идет достаточно интенсивно. (период
полужизни молекул ДНК в тканях мыши составляет от 1 до 5 суток; период полужизни
большинства мРНК у эукариот — несколько суток, а у прокариот — несколько секунд).
Нуклеиновые кислоты (РНК и ДНК) распадаются в организме при посредстве особых
ферментов — нуклеаз. Они ускоряют реакцию разрыва межнуклеотидных фосфодиэфирных
связей в молекулах нуклеиновых кислот и принадлежат, к классу ферментов гидролаз —
фосфодиэстераз.
Нуклеазы, действующие на внутренние межнуклеотидные связи в молекулах ДНК и
РНК, называются эндонуклеазами.
При их участии осуществляется деполимеризация нуклеиновых кислот до
олигонуклеотидов.
Нуклеазы, ускоряющие реакции последовательного отщепления нуклеотидов от РНК,
ДНК или их фрагментов, начиная с конца полинуклеотидной цепи, называют
экзонуклеазами. Они обеспечивают распад нуклеиновых кислот до свободных нуклеотидов.
В зависимости от специфичности действия среди нуклеаз различают рибонуклеазы и
дезоксирибонуклеазы. Первые ускоряют реакции распада как внутренних, так и внешних
(концевых) межнуклеотидных связей в молекулах РНК. Вторые выполняют такую же
функцию по отношению к ДНК. Существует большая группа неспецифических эндо и
экзонуклеаз, действующих одновременно и на РНК, и на ДНК.
По характеру действия на фосфодиэфирные связи в молекулах нуклеиновых кислот
нуклеазы делят на две категории. Одни из них ускоряют реакцию гидролиза сложноэфирной
связи межнуклеозидного фосфата с 3'углеродным атомом остатка рибозы или
дезоксирибозы, а другие — с 5'углеродным атомом. Поэтому в названиях указанных
ферментов всегда подчеркивается, гидролиз какой из связей ускоряет данная нуклеаза.
Нуклеазы, расщепляющие связь Ф — 5'С, называются З'нуклеазами, а расщепляющие
связь Р — З'С именуют 5'нуклеазамн.
Под воздействием рибонуклеазы происходит разрыв:
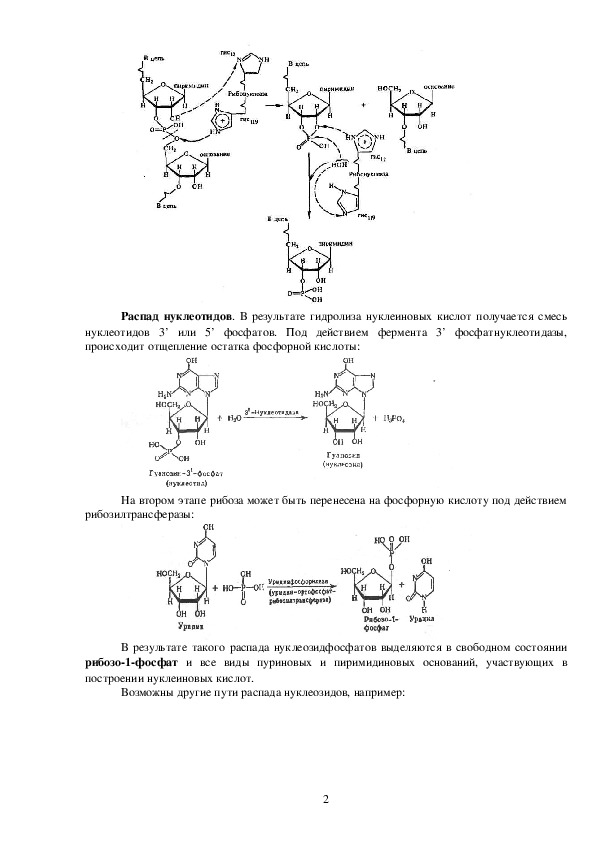
1Распад нуклеотидов. В результате гидролиза нуклеиновых кислот получается смесь
нуклеотидов 3’ или 5’ фосфатов. Под действием фермента 3’ фосфатнуклеотидазы,
происходит отщепление остатка фосфорной кислоты:
На втором этапе рибоза может быть перенесена на фосфорную кислоту под действием
рибозилтрансферазы:
В результате такого распада нуклеозидфосфатов выделяются в свободном состоянии
рибозо1фосфат и все виды пуриновых и пиримидиновых оснований, участвующих в
построении нуклеиновых кислот.
Возможны другие пути распада нуклеозидов, например:
2Далее углевод и азотистые основания видоизменяются.
Рибоза и рибозо1фосфат включаются в реакции обмена, характерные для углеводов.
Пуриновые и пиримидиновые основания претерпевают дальнейший распад и
превращаются в те или иные простейшие азотсодержащие продукты, которые далее либо
выводятся из организма, либо откладываются в нем.
Синтез нуклеиновых кислот в клетке осуществляется в процессах репликации и
транскрипции (см. лекцию 7,8)
Распад пуриновых и пиримидиновых оснований. Первая фаза распада пуриновых и
пиримидиновых оснований заключается в дезаминировании тех из них, которые обладают
аминогруппами. Этот процесс осуществляется при посредстве специфических аминогидролаз.
В результате аденин превращается в гипоксантин:
Дальнейшая судьба дезаминированных пуриновых оснований различна. Гипоксантин и
ксантин окисляются в мочевую кислоту:
Реакция окисления гипоксантина в ксантин, а последнего в мочевую кислоту
ускоряется ксантиноксидазой оксидоредуктазой с широким спектром действия,
представляющей собой молибденсодержащий флавопротеин.
У ряда животных (человекообразные обезьяны, птицы, рептилии, тутовый шелкопряд)
и человека конечным продуктом распада пуриновых оснований является мочевая кислота,
которая и выводится из организма.
У большинства животных и растений есть ферменты и ферментные системы,
способные ускорять реакции дальнейшего распада мочевой кислоты. От названия мочевой
кислоты acidum uricum и по характеру действия, выражающемуся в расщеплении (лизисе) ее,
эти ферменты получили наименование ферментов уриколиза.
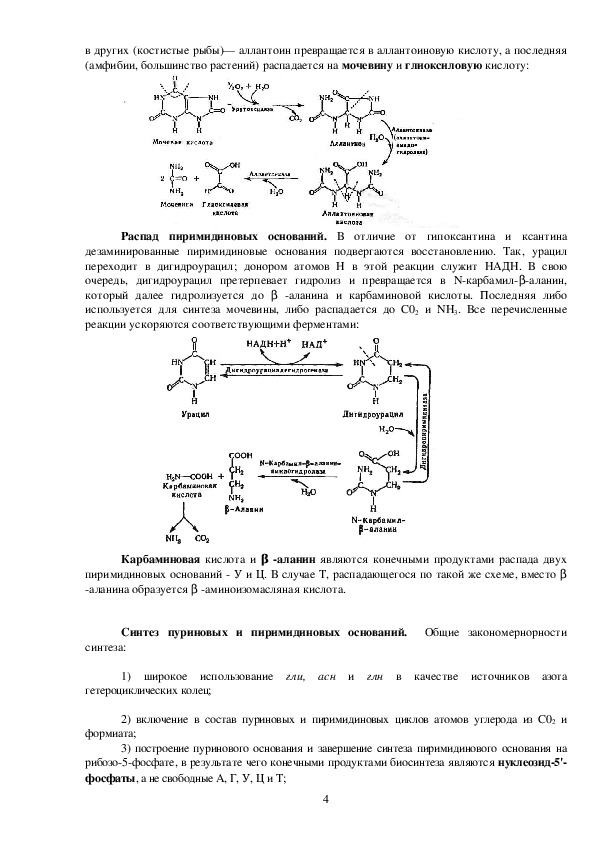
В одних случаях
(млекопитающие, насекомые) уриколиз сводится к окислению мочевой кислоты в аллантоин;
3в других (костистые рыбы)— аллантоин превращается в аллантоиновую кислоту, а последняя
(амфибии, большинство растений) распадается на мочевину и глиоксиловую кислоту:
Распад пиримидиновых оснований. В отличие от гипоксантина и ксантина
дезаминированные пиримидиновые основания подвергаются восстановлению. Так, урацил
переходит в дигидроурацил; донором атомов Н в этой реакции служит НАДН. В свою
очередь, дигидроурацил претерпевает гидролиз и превращается в Nкарбамил аланин,
который далее гидролизуется до
аланина и карбаминовой кислоты. Последняя либо
используется для синтеза мочевины, либо распадается до С02 и NH3. Все перечисленные
реакции ускоряются соответствующими ферментами:
β
β
Карбаминовая кислота и
являются конечными продуктами распада двух
пиримидиновых оснований У и Ц. В случае Т, распадающегося по такой же схеме, вместо β
аланина образуется
β
аминоизомасляная кислота.
β
аланин
Синтез пуриновых и пиримидиновых оснований. Общие закономернорности
синтеза:
1) широкое использование гли, асн и глн в качестве источников азота
гетероциклических колец;
2) включение в состав пуриновых и пиримидиновых циклов атомов углерода из С02 и
формиата;
3) построение пуринового основания и завершение синтеза пиримидинового основания на
рибозо5фосфате, в результате чего конечными продуктами биосинтеза являются нуклеозид5'
фосфаты, а не свободные А, Г, У, Ц и Т;
44) ферментативный характер всех реакций, осуществляющихся в процессе новообразования
нуклеотидов;
5) возникновение на определенном этапе биосинтеза предшественников, из которых потом
формируются уже индивидуальные нуклеозид5'фосфаты.
Механизм биосинтеза пиримидиновых основании. Подготовительной реакцией,
открывающей этот синтез, является реакция образования карбамилфосфата из NH3 и СО2 при
участии АТФ:
При участии специфического фермента остаток карбаминовой кислоты (карбамил)
переносится на аминогруппу аспарагиновой кислоты с образованием карбамиласпарагиновой
кислоты. Эта реакцию является первой специфической реакцией в синтезе пиримидиновых
нуклеотидов:
При сближении NH2 и СООНгрупп в молекуле карбамиласпарагиновой кислоты
между ними осуществляется взаимодействие с выделением молекул воды.
Эта реакция катализируется ферментом из класса гидролаз дигидрооротазой,
названной так от дигидрооротовой кислоты, гидролиз которой она ускоряет вследствие
обратимости данной реакции:
Дигидрооротовая кислота ферментативно окисляется. Снятие двух атомов Н
осуществляется первичной дегидрогеназой либо с НАД+ или НАДФ+, либо с ФАД в качестве
кофермента:
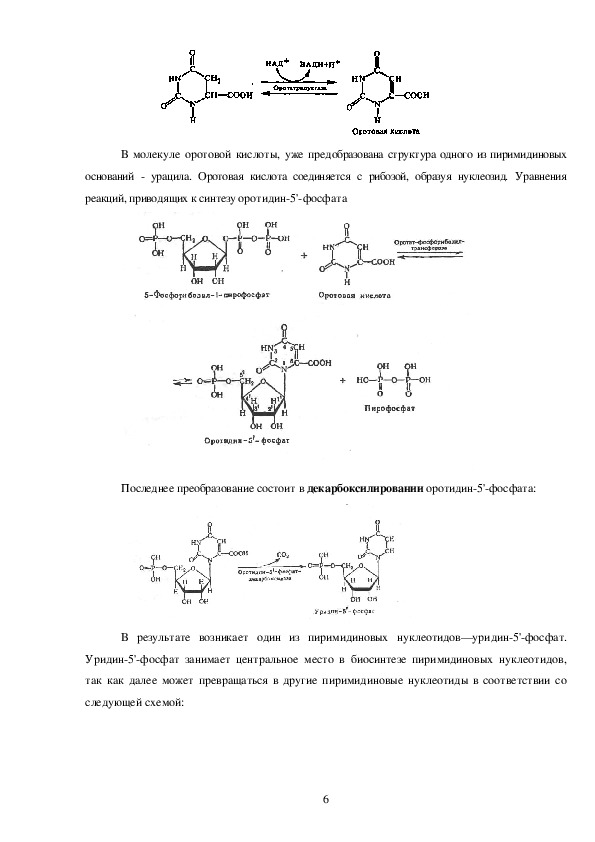
5В молекуле оротовой кислоты, уже предобразована структура одного из пиримидиновых
оснований урацила. Оротовая кислота соединяется с рибозой, образуя нуклеозид. Уравнения
реакций, приводящих к синтезу оротидин5'фосфата
Последнее преобразование состоит в декарбоксилировании оротидин5'фосфата:
В результате возникает один из пиримидиновых нуклеотидов—уридин5'фосфат.
Уридин5'фосфат занимает центральное место в биосинтезе пиримидиновых нуклеотидов,
так как далее может превращаться в другие пиримидиновые нуклеотиды в соответствии со
следующей схемой:
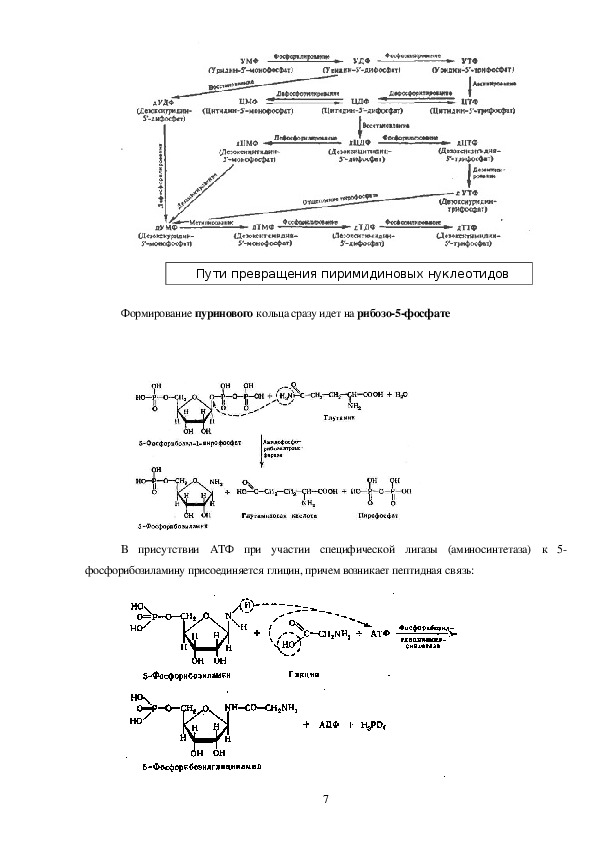
6Пути превращения пиримидиновых нуклеотидов
Формирование пуринового кольца сразу идет на рибозо5фосфате
В присутствии АТФ при участии специфической лигазы (аминосинтетаза) к 5
фосфорибозиламину присоединяется глицин, причем возникает пептидная связь:
7Схема, включающая главные этапы синтеза:
Из приведенной схемы ясно, что 1й атом азота пуринового цикла ведет свое
происхождение от аспарагиновой кислоты, 3й и 9й от глутамина, а 7й от глицина. Что
касается происхождения атомов углерода пуринового кольца, то видно, что источниками их
явились формиат (2й и 8й атомы углерода), глицин (4й и 5й) и СО2 (6й атом). Химические
уравнения, приведенные на схеме 2, показывают детали включения тех или иных атомов N и С из
состава перечисленных соединений в пуриновую часть нуклеотида в процессе его биосинтеза.
Пиримидиновое кольцо пиримидиновых нуклеотидов синтезируется в организме из
аналогичных соединений: NH3, CО2 и аспарагиновой кислоты.
Таким образом, исходные вещества для биосинтеза пуриновых и пиримидиновых оснований всегда
присутствуют в организме:
аммиак, формильная группа и оксид углерода (IV) образуются в процессе деструкции
разнообразных органических соединений или поступают в организм извне;
глутаминовая и аспарагиновая кислоты и их амиды представляют первичные в большом
объеме синтезируемые аминокислоты, что имеет большое значение для обеспечения
беспрепятственного синтеза этих важнейших для организма соединений.
8