НУКЛЕОЗИДЫ- соединения азотистого основания и углеводов (рибозы и дезоксирибозы). Нуклеозиды образуются за счет N-гликозидной связи между ДЕВЯТЫМ атомом азота у ПУРИНОВЫХ (первым атомом азота- у ПИРИМИДИНОВЫХ) оснований и гидроксилом ПЕРВОГО атома углерода рибозы или дезоксирибозы. Во избежание путаницы, нумерация атомов азотистых оснований осуществляется арабскими цифрами, а у атомов углерода рибоз- арабскими цифрами со “штрихом”.
НУКЛЕОТИДЫ отличаются от нуклеозидов наличием остатков фосфорной кислоты (от одного до трех), связанных простой эфирной связью с гидроксилом 5’ атома углерода рибоз. Остатки фосфорных кислот между собой также связаны простой эфирной связью. В зависимости от числа остатков фосфорной кислоты в нуклеотидах различают МОНО-, ДИ- и ТРИФОСФОНУКЛЕОТИДЫ. Их номенклатура приведена в таблице:
Тема. Нуклеиновые кислоты. Структура, функции ДНК РНК.
Обмен нуклеиновых кислот.
ПЛАН
1. Строение нуклеиновых кислот. Строение нуклеозидов, нуклеотидов
2. Функции и структура нуклеиновых кислот.
3. Типы РНК (тРНК, рРНК, мРНК).
4. Обмен нуклеозидфосфатов. Распад пуриновых и пиримидиновых оснований.
Биосинтез нуклеозидов.
Химия нуклеиновых кислот развивается быстрыми темпами. Это связано с важностью
биологической роли этих соединений в организме человека, животных, растений ,
микроорганизмов.
К нуклеиновым кислотам относятся высокомолекулярные вещества имеющие
определенный химический состав. При полном гидролизе нуклеиновых кислот (НК)
выделены соединения:
а) пуриновые и пиримидиновые основания;
б) моносахариды РИБОЗА или ДЕЗОКСИРИБОЗА (С5Н10О5);в) фосфорная
кислота.
HOCH2
H
H
OH
OH
1
H
H
H
HOCH2
OH
1
H
H
H
H
OH OH
β Dдезоксирибоза β Dрибоза
Строение моносахаридов:
O
NH2
HN
N
N
N
NH2
N
N
H
N
N
H
Аденин (А) Гуанин (Г)
Строение пуриновых оснований:NH2
N
N
HN
O
N
HN
O
N
H
O
CH3
Цитозин (Ц) ) Урацил (У) Тимин (Т)
Строение пиримидиновых оснований:
Различают рибонуклеиновые(РНК) и дезоксирибонуклеиновые (ДНК) кислоты,
отличающиеся по составу, строению и функциям.
Отличия в составе нуклеиновых кислот.
Отличающиеся компоненты
ДНК
ДЕЗОКСИРИБОЗА
ТИМИН
РНК
РИБОЗА
УРАЦИЛ
Одинаковые
компоненты
АДЕНИН
ГУАНИН
ЦИТОЗИН
НУКЛЕОЗИДЫ соединения азотистого основания и углеводов (рибозы и
дезоксирибозы). Нуклеозиды образуются за счет Nгликозидной связи между ДЕВЯТЫМ
атомом азота у ПУРИНОВЫХ (первым атомом азота у ПИРИМИДИНОВЫХ) оснований и
гидроксилом ПЕРВОГО атома углерода рибозы или дезоксирибозы. Во избежание
путаницы, нумерация атомов азотистых оснований осуществляется арабскими цифрами, а
у атомов углерода рибоз арабскими цифрами со “штрихом”.
НУКЛЕОТИДЫ отличаются от нуклеозидов наличием остатков фосфорной
кислоты (от одного до трех), связанных простой эфирной связью с гидроксилом 5’ атома
углерода рибоз. Остатки фосфорных кислот между собой также связаны простой эфирной
связью. В зависимости от числа остатков фосфорной кислоты в нуклеотидах различают
МОНО, ДИ и ТРИФОСФОНУКЛЕОТИДЫ. Их номенклатура приведена в таблице:
азотистые
нуклеозиды
Нуклеотиды
основания
аденин
гуанин
цитозин
полное название
Сокращенное
аденозин
гуанозин
цитидин
Аденозинмонофосфат
Гуанозинмонофосфат
Цитидинмонофосфат
АМФ
ГМФ
ЦМФурацил
тимин
уридин
тимидин
Уридинмонофосфат
тимидинмонофосфат
УМФ
ТМФ
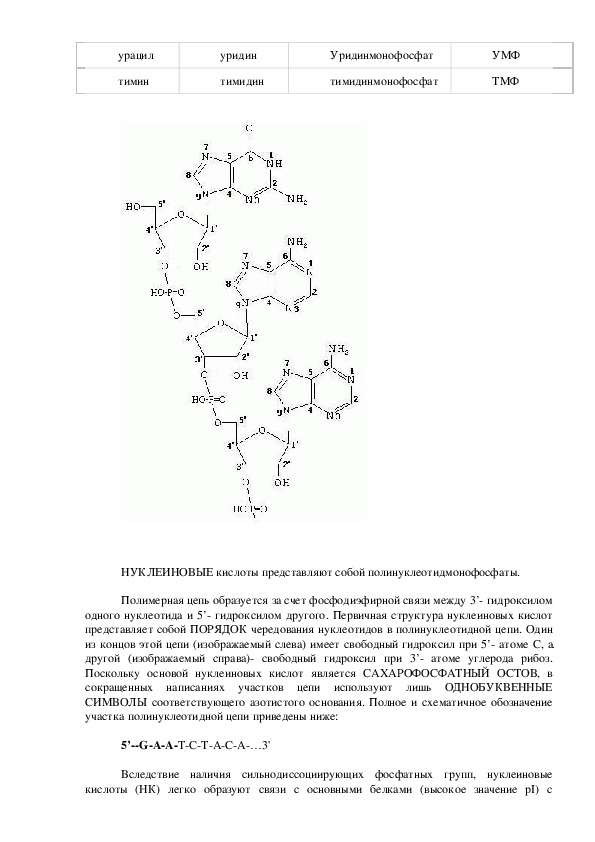
НУКЛЕИНОВЫЕ кислоты представляют собой полинуклеотидмонофосфаты.
Полимерная цепь образуется за счет фосфодиэфирной связи между 3’ гидроксилом
одного нуклеотида и 5’ гидроксилом другого. Первичная структура нуклеиновых кислот
представляет собой ПОРЯДОК чередования нуклеотидов в полинуклеотидной цепи. Один
из концов этой цепи (изображаемый слева) имеет свободный гидроксил при 5’ атоме С, а
другой (изображаемый справа) свободный гидроксил при 3’ атоме углерода рибоз.
Поскольку основой нуклеиновых кислот является САХАРОФОСФАТНЫЙ ОСТОВ, в
сокращенных написаниях участков цепи используют лишь ОДНОБУКВЕННЫЕ
СИМВОЛЫ соответствующего азотистого основания. Полное и схематичное обозначение
участка полинуклеотидной цепи приведены ниже:
5’GAATCTACA…3'
Вследствие наличия сильнодиссоциирующих фосфатных групп, нуклеиновые
кислоты (НК) легко образуют связи с основными белками (высокое значение pI) собразованием НУКЛЕОПРОТЕИНОВ. Протеины отделяются от НК детергентами или
после расщепления белков протеиназами НК осаждаются спиртом.
Подобно белкам, ДНК имеют первичную, вторичную и третичную структуру.
ПЕРВИЧНАЯ СТРУКТУРА есть последовательность чередования нуклеотидов в
цепи ДНК и РНК. Сложность расшифровки структуры связана с наличием всего 4х видов
нуклеотидов при их огромном числе в молекуле.
ПРАВИЛА ЧАРГАФФА
1) сумма ПУРИНОВЫХ оснований в молекуле ДНК всегда равна сумме
ПИРИМИДИНОВЫХ оснований.
2) А=Т, А/Т= 1; 3) Г=Ц, Г/Ц=1; 4) А+Т=Г+Ц; 5) если А+Т>Г+Ц, то АТтип.
ВТОРИЧНАЯ СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ
Расшифрована впервые в 1953 г. Уотсоном и Крик (Watson, Crick).
Вторичная структура ДНК представляет собой свернутые в спираль ДВЕ
комплементарно взаимодействующие и антипараллельные полинуклеотидные цепи.
Образование вторичной структуры НК возможно вследствие проявления эффектов
КОМПЛЕМЕНТАРНОСТИ и СТЭКИНГВЗАИМОДЕЙСТВИЙ.
Очень часто наблюдаются двунитевые спирализованные молекулы ДНК, замкнутые
в кольцо с ковалентно связанными концами. Они не имеют разрывов у каждой в
отдельности полинуклеотидной цепи. Подобные кольцевые ДНК, как правило,
СУПЕРСПИРАЛИЗОВАНЫ, то есть кольцо дополнительно закручено в спираль.
Суперспирализация ПРАВИЛО, а не ИСКЛЮЧЕНИЕ, при условии отсутствия разрывов в
фосфодиэфирных связях полинуклеотидной цепи.
КОМПЛЕМЕНТАРНОСТЬ последовательность нуклеотидов в одной цепи
автоматически определяет строго соответствующую ей последовательность нуклеотидов в
КОМПЛЕМЕНТАРНОЙ ей цепи.
Так, азотистое основание Аденин (А) всегда взаимодействует только с
комплементарным ему азотистым основанием Тимин (Т) в молекулах ДНК. Одновременно
азотистые основания Гуанин (Г) одной цепи взаимодействует только с комплементарними
им азотистыми основаниями Цитозин (Ц) другой цепи (как в ДНК, так и в РНК).
Комплементарность оснований обеспечивается системой водородных связей. В
молекулах РНК, имеющих, в основном, однонитевую структуру, на отдельных участках,
азотистые основания А взаимодействуют с комплементарными им азотистыми
основаниями
(У).
Урацил
Аналогично происходит взаимодействие в процессах транскрипции, когда на матрице
ДНК синтезируется молекула РНК (матричная, транспортная и т.д.), и наоборот,
когда при участии РЕВЕРСТРАНСКРИПТАЗЫ происходит синтез кДНК на матрице РНК.СТЭКИНГВЗАИМОДЕЙСТВИЯ особого рода (ВандерВаальсовы)
взаимодействия между выложенными в стопку (как монеты) друг над другом азотистых
оснований. SBS (sidebyside) форма (бок о бок). Две цепи расположены прямо и
незакручены.
ТРЕТИЧНАЯ СТРУКТУРА ДНК
ДНК имеют формы ЛИНЕЙНУЮ, КОЛЬЦЕВУЮ, 2х и 1ЦЕПОЧЕЧНУЮ.
Двуцепочечные ДНК с "липкими" концами могут образовывать кольцо, которое далее
ковалентно сшивается по сахарофосфатной цепи при помощи ДНКлигазы.
Третичная структура ДНК у эукариотических клеток отличается тем, что
многократная спирализация ДНК сопровождается образованием комплексов с белками.
46 хромосом (хроматид) человека организованы в 23 пары. Средняя длина
хромосомы составляет 130 млн. пар оснований и имеет длину 5 см. Хромосома №1 263
млн. п.о., хромосома № 46 –меньше 50 млн. п.о. Если проложить все ДНК в В
конформации в линию, то их общая длина превысит 2 метра. Человеческая хромосома 16
имеет 2,5 мкм в длину, а длина самой ДНК 3,7 см.
Понятно, что уместить такой длины ДНК в ядре возможно только путем ее определенной
упаковки. При образовании третичной структуры ДНК человека происходит в среднем
уменьшение ее размеров в 100 тысяч раз.
Рибонкулеиновые кислоты. Различают тРНК, рРНК, м РНК
РНК состоит из одной полинуклеотидной цепи, закрученной на себя, образует
короткие двуспиральные шпильки в палиндромных участках (Г с Ц, А с У)
тРНК самые мелкие молекулы (ММ=2330 тыс.) тРНК переносчик аминокислот.
Каждая тРНК переносит только одну аминокислоту, но на одну аминокислоту имеется
более одной тРНК. Всего известно 61 тРНК.
Строение тРНК. Транспортная РНК имеет:
акцептирующий участок (присоединяет АК, при участии АТФ),
общий участок (петля дигидроуридина) обеспечивает связь со специфическим
ферментом,
характерный участок (петля псевдоуридина) всегда содержит последовательность 5
'TyЦГ3', этой петлей взаимодействует с рибосомой, антикодоновая петля содержит 3
нуклеотида, комплементарных кодону данной аминокислоты в мРНК, например, кодону 5'
ГЦЦ3' в мРНК соответствует антикодон 3'ЦГГ5' в тРНК, чем обеспечивается
специфичность взаимодействия с матричной РНК.
рРНК является структурным компонентом рибосом
. В состав рибосом входят
различные по S (единица Сведберга) рРНК и с белками формируют структуру
субъединиц различных рибосом.
мРНК служит матрицей для синтеза белков клетки, информация о которых
заложена в ДНК т.е. переноса и реализации генетической информации с ДНК.Таким образом , нуклеиновые кислоты выполняют важнейшие функции : хранение,
передачу и реализацию генетической информации клетки и организма в целом.
ФИЗИКОХИМИЧЕСКИЕ СВОЙСТВА ДНК
ДНК – вещества белого цвета, плохо растворимы в воде в свободном состоянии,
имеют волокнистое строение, хорошо растворимы в крепких солевых растворах.
Растворы обладают высокой вязкостью и двойным лучепреломлением. Оптически
активны. Имеют отрицательный электрический заряд и подвижны в электрическом поле.
Молекулярная масса ДНК определяется рядом методов, в том числе:
а) Ультрацентрифугированием в градиенте CsCl (ММ от 200000 до 109);
б) по вязкости растворов (ММ> 109, поскольку при центрифугировании такие длинные
молекулы разрываются под действием собственного веса).
Денатурация ДНК заключается в разрыве Нсвязей и стэкингвзаимодействий, что
приводит к расплетанию и разделению цепей. (БЕЗ РАЗРЫВА КОВАЛЕНТНЫХ
СВЯЗЕЙ!) под действием температуры или рН.
О степени денатурации судят по изменению интенсивности поглощения в
ультрафиолете при = 260 нм, поскольку дезэкранирование азотистых оснований в
результате расплетания цепей устраняется, что вызывает увеличение степени поглощения
раствором ДНК ультрафиолета указанной длины волны (гиперхромный эффект).
Денатурацию иначе называют плавлением, а температура плавления соответствует
моменту 50%ной денатурации молекулы. Т плав. разная для каждой ДНК.
Комплементарные цепи ДНК, разделенные при денатурации, при определенных
условиях могут вновь соединиться в двойную спираль. Этот процесс называется
ренатурацией. Если денатурация произошла не полностью и хотя бы несколько оснований
не утратили взаимодействия водородными связями, ренатурация протекает очень быстро.
Ренатурация возможна даже при полностью разделенных цепях. В таком случае
ренатурация требует точного совмещения цепей ДНК, которое может привести к
реассоциации, и этот процесс медленный, к тому же, зависит от концентрации цепей в
растворе. Как правило, выдерживание раствора ДНК при температуре на 1015 С ниже
температуры плавления в условиях средней ионной силы (0,15 М) обеспечивает наиболее
благоприятные условия для ренатурации. При более низкой ионной силе ренатурации
мешает взаимное отталкивание фосфатных групп. Реассоциация начинается со
взаимодействия коротких комплементарных последовательностей нуклеотидов, время
существования которых в ассоциированном состоянии может оказаться
непродолжительны, если соседние участки ДНК окажутся некомплементарными.